хллсмнцкнаскхмш (юмрхрекю)
яХМРЕГ ХЛЛСМНЦКНАСКХМНБ: ХЛЛСМНЦКНАСКХМШ ЯХМРЕГХПСЧРЯЪ ОКЮГЛЮРХВЕЯЙХЛХ ЙКЕРЙЮЛХ, ЙНРНПШЕ НАПЮГСЧРЯЪ ХГ РПЮМЯТНПЛХПНБЮММШУ, ЯРХЛСКХПНБЮММШУ ЮМРХЦЕМНЛ б-КХЛТНЖХРНБ (б-ХЛЛСМНАКЮЯРНБ).
Bce ЛНКЕЙСКШ ХЛЛСМНЦКНАСКХМНБ, ЯХМРЕГХПНБЮММШУ НРДЕКЭМНИ ОКЮГЛЮРХВЕЯЙНИ ЙКЕРЙНИ, ХДЕМРХВМШ Х ХЛЕЧРЯОЕЖХТХ- ВЕЯЙСЧ ПЕЮЙРХБМНЯРЭ ОПНРХБ ЕДХМЯРБЕММНИ ЮМРХЦЕММНИ ДЕРЕПЛХМЮМРШ. юМЮКНЦХВМН, БЯЕ ОКЮГЛЮРХВЕЯЙХЕ ЙКЕРЙХ, ОНКСВЕММШЕ ОСРЕЛ РПЮМЯТНПЛЮЖХХ Х ОПНКХТЕПЮЖХХ НДМНЦН б-КХЛТНЖХРЮ-ОПЕДЫЕЯРБЕММХЙЮ, ХДЕМРХВМШ; РН ЕЯРЭ, НМХ ЯНЯРЮБКЪЧР ЙКНМ. лНКЕЙСКШ ХЛЛСМНЦКНАСКХ- МНБ,ЯХМРЕГХПНБЮММШЕ ЙКЕРЙЮЛХ ПЮГКХВМШУ ЙКНМНБ ОКЮГ- ЛЮРХВЕЯЙХУЙКЕРНЙ,ХЛЕЧРПЮГКХВМШЕ ОНЯКЕДНБЮРЕКЭМНЯРХ ЮЛХМНЙХЯКНР, ВРН НАСЯКНБКХБЮЕР ПЮГКХВМСЧ РПЕРХВМСЧ ЯРПСЙРСПС ЛНКЕЙСК Х ОПХДЮЕР ХМСЧ ЯОЕЖХТХВМНЯРЭ ЮМРХРЕКС (РН ЕЯРЭ, НМХ ПЕЮЦХПСЧР Я ПЮГМШЛХ ЮМРХЦЕМЮ-ЛХ). щРХ ПЮГКХВХЪ Б ОНЯКЕДНБЮРЕКЭМНЯРХ ЮЛХМНЙХЯКНР ОПНХЯУНДЪР Б РЮЙ МЮГШБЮЕЛНЛ V (БЮПХЮАЕКЭМНЛ, ОЕПЕЛЕММНЛ) СВЮЯРЙЕ ЛНКЕЙСКШ ХЛЛСМНЦКНАСКХМЮ (ПХЯ. 10.3).
яРПСЙРСПЮ ХЛЛСМНЦКНАСКХМНБ (ПХЯ. 10.3): АНКЭЬХМЯРБН ЛНКЕЙСК ХЛЛСМНЦКНАСКХМНБ ЯНЯРЮБКЕМШ ХГ ДБСУ РЪФЕКШУ (H) ЖЕОЕИ Х ДБСУ КЕЦЙХУ (L) ЖЕОЕИ, ЯНЕДХМЕМ- МШУДХЯСКЭТХДМШЛХ ЯБЪГЪЛХ. кЕЦЙХЕ ЖЕОХ ЯНЯРНЪР ХКХ ХГ ДБСУ k ЖЕОЕИ, ХКХ ХГ ДБСУ 1 ЖЕОЕИ. рЪФЕКШЕ ЖЕОХ ЛНЦСР АШРЭ НДМНЦН ХГ ОЪРХ ЙКЮЯЯНБ (IgA, IgG, IgM, IgD, Х IgE) (РЮАК.10.4). яСЫЕЯРБСЕРМЕЯЙНКЭЙН ОНДЙКЮЯЯНБРЪ- ФЕКШУ ЖЕОЕИ (ХГНРХОШ). щРХ ПЮГКХВМШЕ ЖЕОХ ХЛЛСМНЦКНАСКХМНБ ЪБКЪЧРЯЪ ЮМРХЦЕМЮЛХ ДКЪ ФХБНРМШУ Х ХЛЕЧР НРКХВЮЧЫХЕЯЪ ЮМРХЦЕММШЕ ДЕРЕПЛХМЮМРШ, ОНЩРНЛС, ОПХ ББЕДЕМХХ ХУ ФХБНРМШЛ, ЮМРХРЕКЮ, ОПНХГБНДХЛШЕ ОПНРХБ МХУ, ЛНЦСР ХЯОНКЭГНБЮРЭЯЪ ДКЪ ПЮЯОНГМЮБЮМХЪ Х НОПЕДЕКЕМХЪ ПЮГКХВМШУРХОНБКЕЦЙХУ ЖЕОЕИХ ЙКЮЯЯНБ РЪФЕКШУ ЖЕОЕИ С ВЕКНБЕЙЮ.
йЮФДЮЪ ЖЕКЭ ХЛЕЕР ОНЯРНЪММШИ Х БЮПХЮАЕКЭМШИ СВЮЯРНЙ. оНЯРНЪММШИ СВЮЯРНЙ НЯРЮЕРЯЪ ОНЯРНЪММШЛ Б ОНЯКЕДНБЮРЕКЭМНЯРХ ЮЛХМНЙХЯКНР Х ЮМРХЦЕММНЯРХ Б ОПЕ- ДЕКЮУДЮММНЦН ЙКЮЯЯЮ ХЛЛСМНЦКНАСКХМНБ; БЮПХЮАЕКЭМШИ СВЮЯРНЙ, МЮОПНРХБ, УЮПЮЙРЕПХГСЕРЯЪ АНКЭЬНИ МЕОНЯРНЪММНЯРЭЧ ОНЯКЕДНБЮРЕКЭМНЯРХ ЮЛХМНЙХЯКНР.
хЛЕММН Б БЮПХЮАЕКЭМНИ ВЮЯРХ ЖЕОХ ОПНХЯУНДХР ПЕЮЙЖХЪ ЯНЕДХМЕМХЪ Я ЮМРХЦЕМНЛ. йЮФДЮЪ ЛНКЕЙСКЮ IgG ЯНЯРНХР ХГ ДБСУ ЯНЕДХМЕММШУ ЖЕОЕИ, ЙНРНПШЕ ТНПЛХПСЧР ДБЮ ЮМ- РХЦЕМ-ЯБЪГШБЮЧЫХУСВЮЯРЙЮ (ПХЯ. 10.3). Ha БЮПХЮАЕКЭМНЛ СВЮЯРЙЕ ЙЮФДНИ ЖЕОХ ХЛЕЧРЯЪ ЦХОЕПБЮПХЮАЕКЭМШЕ СВЮЯРЙХ ≈ РПХ Б КЕЦЙХУ ЖЕОЪУ Х ВЕРШПЕ Б РЪФЕКШУ ЖЕОЪУ. пЮГМНБХДМНЯРХ ОНЯКЕДНБЮРЕКЭМНЯРХ ЮЛХМНЙХЯКНР БЩРХУ ЦХОЕПБЮПХЮАЕКЭМШУСВЮЯРЙЮУНОПЕДЕКЪЧРЯОЕЖХ- ТХВМНЯРЭ ЮМРХРЕКЮ. оПХ НОПЕДЕКЕММШУ СЯКНБХЪУ ЩРХ ЦХОЕПБЮПХЮАЕКЭМШЕ НАКЮЯРХ ЛНЦСР РЮЙФЕ БШЯРСОЮРЭ Б ПНКХ ЮМРХЦЕМНБ (ХДХНРХОШ). юМРХРЕКН ОПНРХБ ХДХНРХ- noB,T.e. ОПНХГБНДХЛНЕ ОПНРХБ ЦХОЕПБЮПХЮАЕКЭМНИ НА- КЮЯРХЮМРХРЕК,ХЛЕЕРНЦПЮМХВЕММШИДХЮОЮГНМ ПЕЮЙРХБМНЯРХ Х ЯНЕДХМЪЕРЯЪ РНКЭЙН Я ЛНКЕЙСКЮЛХ ХЛЛСМНЦКНАСКХМЮ, ХЛЕЧЫХЛХ ДЮММСЧ ЦХОЕПБЮПХЮАЕКЭМСЧ НАКЮЯРЭ. B ЯСЫМНЯРХ, ПЕЮЙРХБМНЯРЭ ЮМРХРЕК ОПНРХБ ХДХН- РХОНБ НЦПЮМХВЕМЮ ХЯЙКЧВХРЕКЭМН ЯОЕЖХТХВЕЯЙХЛХ ЮМРХРЕКЮЛХ, ОНКСВЕММШЛХ ХГ ЕДХМЯРБЕММНЦН ЙКНМЮ. уНРЪ БШЬЕНОХЯЮММНЕ НРМНЯХРЯЪ ЯРПНЦН Й IgG, ДПСЦХЕ ЙКЮЯЯШ ХЛЛСМНЦКНАСКХМНБ ХЛЕЧРРЮЙСЧ ФЕ НЯМНБМСЧ ЯРПСЙРСПС, ГЮ ХЯЙКЧВЕМХЕЛ РНЦН, ВРН IgM ЪБКЪЕРЯЪ ОЕМРЮЛЕПНЛ (РН ЕЯРЭ, ЯНЯРНХР ХГ 5 НЯМНБМШУ ЕДХМХЖ (ЛНКЕЙСК), ЯБЪГЮММШУ Б НАКЮЯРХ Fc-ЙНМЖНБ), a IgA НАШВМН ЯСЫЕЯРБСЕР ЙЮЙДХЛЕП.оНЯРНЪММШИСВЮЯРНЙ ЙЮФДНИ ЛНКЕЙСКШ ХЛЛСМНЦКНАСКХМЮ ХЛЕЕР ПЕЖЕОРНПШ ДКЪ ЙНЛОКЕЛЕМРЮ, Ю РЮЙФЕ
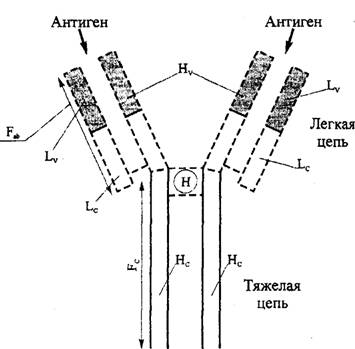
пХЯ. 10.3 яРПНЕМХЕ ХЛЛСМНЦКНАСКХМНБ (IgG)
IgD Х IgE ХЛЕЧР ЯУНДМСЧ ЯРПСЙРСПС. IgA ≈ ДХЛЕП, Ю IgM ≈ ОЕМРЮЛЕП. юМРХЦЕМ-ЯБЪГШБЮЧЫХЕ ЛЕЯРЮ ТНП-ЛХПСЧР- ЯЪ ЙНМЖЮЛХ РЪФЕКНИ Х КЕЦЙНИ ЖЕОЕИ; ЙЮФДЮЪ ЛНКЕЙСКЮ IgG ХЛЕЕРДБЮ ЛЕЯРЮ ЯБЪГШБЮМХЪ. оПХ ЩМ-ГХЛЮРХВЕЯЙНЛ ПЮЯЫЕОКЕМХХ НАПЮГСЧРЯЪ ЯКЕДСЧЫХЕ ТПЮЦЛЕМРШ: Fc-ТПЮЦЛЕМР co- ДЕПФХРСВЮЯРЙХ НАНХУ ОНЯРНЪММШУ ВЮЯРЕИ; Fab-ТПЮЦЛЕМР ЯНДЕПФХР КЕЦЙСЧ Х ВЮЯРЭ РЪФЕКНИ ЖЕОХ Я НДМХЛ ЮМРХЦЕМ-ЯБЪ- ГШБЮЧЫХЛ СВЮЯРЙНЛ. п(Юэ)'2-ТПЮЦЛЕМР ЯНЯРНХР ХГ ДБСУ ЯБЪГЮММШУ ЛЕФДС ЯНАНИ Fab-ТПЮЦЛЕМРНБ. H ≈ НАКЮЯРЭ ╚БХКЙХ╩; HV, LV ≈ БЮПХЮАЕКЭМШЕ ЙНМЖШ РЪФЕКНИ Х КЕЦЙНИ ЖЕОЕИ; мя, LC ≈ ОНЯРНЪММШЕ ЙНМЖШ РЪФЕКНИ Х КЕЦЙНИ ЖЕОЕИ;------------------------------------- ДХ
ЯСКЭТХДМШЕ ЯБЪГХ ЛЕФДС ЖЕОЪЛХ.
рЮАКХЖЮ 10.4.
йКЮЯЯШ ХЛЛСМНЦКНАСКХМНБ
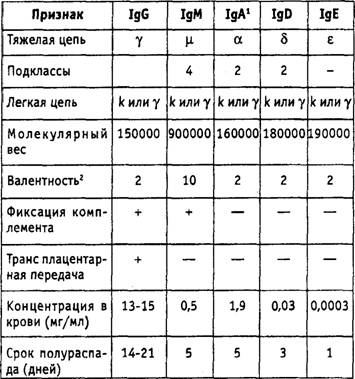
1 нАПЮРХРЕ БМХЛЮМХЕ, ВРН IgA ЯХМРЕГХПСЕРЯЪ Б ОКЮГЛН- ЖХРЮУ ЙЮЙ ЛНМНЛЕП (MB=160 000) Х ЯЕЙПЕРХПСЕРЯЪ ВЕПЕГ ЩОХРЕКХИ СФЕ Б БХДЕ ДХЛЕПЮ, ОПХ ЩРНЛ ДБЕ ЛНКЕЙСКШ НАЗЕДХМЪ- ЧРЯЪ1-ЖЕОЭЧ,ЮРЮЙФЕДНАЮБКЪЕРЯЪ ЯЕЙПЕРНПМШИ ЙНЛОНМЕМР. йНМЕВМШИ ЛНКЕЙСКЪПМШИ БЕЯ ПЮБЕМ 380 000. яЕЙПЕРНПМШИ ЙНЛОНМЕМР НАПЮГСЕРЯЪ Б ЩОХРЕКХЮКЭМШУ ЙКЕРЙЮУ Х ЯКСФХР ДКЪ НАКЕЦВЕМХЪ ЯЕЙПЕЖХХ ЮМРХРЕКЮ ВЕПЕГ ЙКЕРНВМСЧ ЛЕЛАПЮМС Х ГЮЫХЫЮЕР ЕЦН НР ОЕПЕБЮПХБЮМХЪ ТЕПЛЕМРЮЛХ.
2 йНКХВЕЯРБН ЮМРХЦЕМ-ЯБЪГШБЮЧЫХУСВЮЯРЙНБ МЮ ЛНКЕЙСКЕ.
ХЛЕЕРЯЪ МЮ пЯ-ТПЮЦЛЕМРЕСВЮЯРНЙ,ЙНРНПШИ ЯБЪГШБЮЕРЯЪ Я ЙКЕРЙЮЛХ, ХЛЕЧЫХЛХ Fc-ПЕЖЕОРНПШ (ВРН МЕНАУНДХЛН ДКЪ НЯСЫЕЯРБКЕМХЪ ЙКЕРНВМНЦН ХЛЛСМХРЕРЮ). сМЮЯ- КЕДНБЮММШЕЮМРХЦЕММШЕ ПЮГКХВХЪ ЛЕФДСРЪФЕКШЛХ ЖЕОЪЛХ ЯНЯРЮБКЪЧР ЮККНРХОШ. лНКЕЙСКШ ХЛЛСМНЦКНАСКХМНБ ЛНФМН ПЮГАХРЭ МЮ ВЮЯРХ ПЮГКХВМШЛХ ОПНРЕНКХРХВЕЯЙХЛХ ТЕПЛЕМРЮЛХ. оПХ БНГДЕИЯРБХХ ОЮОЮХМЮ ЛНКЕЙСКЮ ПЮГДЕКЪЕРЯЪ Б НАКЮЯРХ ПЮЯУНФДЕМХЪ РЪФЕКШУ ЖЕОЕИ (╚БХКЙХ╩) (ПХЯ. 10.3) МЮ ДБЮ Fab-ТПЮЦЛЕМРЮ Х НДХМ пЯ-ТПЮЦЛЕМР(ЙПХЯРЮККХГСЧЫХИЯЪ). оЕОЯХМ ПЮГПШБЮЕР ЛНКЕЙСКС МЮ п(Юэ)'2-ТПЮЦЛЕМР Х Fc-ТПЮЦЛЕМР. Fc-ТПЮЦЛЕМРОПЕДЯРЮБКЪЕРЯНАНИ ОНЯРНЪММШИСВЮЯРНЙ; НРЯСРЯРБХЕ ХГЛЕМЪЕЛНЯРХ ОНЯКЕДНБЮРЕКЭМНЯРХ ЮЛХМНЙХЯКНР ≈ ЦКЮБМЮЪ ОПХВХМЮ БНГЛНФМНЯРХ ЙПХЯРЮККХГЮЖХХ ДЮММНЦН ТПЮЦЛЕМРЮ. Fab Х п(Юэ)'Ц-ТПЮЦЛЕМРШ МЕЯСР НДХМ Х ДБЮ ЮМРХЦЕМ-ЯБЪГШБЮЧЫХУ СВЮЯРЙЮ ЯННРБЕРЯРБЕММН. Fc-ТПЮЦЛЕМР МЕЯЕР ЯОЕЖХТХВЕЯЙХЕ ЮМРХЦЕМШ, БЙКЧВЮЪ РЕ, йНРНПШЕ НОПЕДЕКЪЧР ХЛЛСМНКНЦХВЕЯЙНЕ ПЮГКХВХЕ ОЪРХ ЦКЮБМШУ ЙКЮЯЯНБ ЮМРХРЕК. сВЮЯРНЙ ТХЙЯЮЖХХ ЙНЛОКЕЛЕМРЮРЮЙФЕ ПЮЯОНКНФЕМ МЮ Fc-ТПЮЦЛЕМРЕ. лЕРНД ТЕПЛЕМРЮРХБМНЦН ПЮЯЫЕОКЕМХЪ ХЛЕЕР ХЯРНПХВЕЯЙНЕ ГМЮВЕМХЕ Б ОПНЖЕЯЯЕ БШЪЯМЕМХЪ ЯРПСЙРСПШ ХЛЛСМНЦКНАСКХМНБ.
пЕЦСКХПНБЮМХЕ ОПНХГБНДЯРБЮ ЮМРХРЕК: ОПНХГБНДЯРБН ЮМРХРЕК МЮВХМЮЕРЯЪ ОНЯКЕ ЮЙРХБЮЖХХ б-ЙКЕ- РНЙ ЮМРХЦЕМНЛ. лЮЙЯХЛЮКЭМЮЪ ЙНМЖЕМРПЮЖХЪ ЮМРХРЕК Б ЯШБНПНРЙЕ МЮАКЧДЮЕРЯЪ Я 1 ОН 2 МЕДЕКЧ Х ГЮРЕЛ МЮВХМЮЕР ЯМХФЮРЭЯЪ. мЕОПЕПШБМНЕ ОПХЯСРЯРБХЕ ЯБНАНДМНЦН ЮМРХЦЕМЮ ОНДДЕПФХБЮЕР НРБЕР ДН РЕУ ОНП, ОНЙЮ СБЕКХВЕМХЕСПНБМЪ ЮМРХРЕК МЕ ОПХБЕДЕРЙСЯХКЕММНЛС СДЮКЕМХЧ ЮМРХЦЕМЮ Х, РЮЙХЛ НАПЮГНЛ, ОПЕЙПЮЫЕМХЧ ЯРХЛСКЪЖХХ б-ЙКЕРНЙ.
яСЫЕЯРБСЧРРЮЙФЕ АНКЕЕРНМЙХЕ ЛЕУЮМХГЛШ ПЕЦСКЪЖХХ ЯХМРЕГЮ ХЛЛСМНЦКНАСКХМНБ. T- УЕКОЕПШ (я04-ОНГХРХБМШЕ) ХЦПЮЧР БЮФМСЧ ПНКЭ Б ПЕЦСКЪЖХХ НРБЕРЮ б-ЙКЕРНЙ МЮ АНКЭЬНЕ ЙНКХВЕЯРБН ЮМРХЦЕМНБ Х ХУ ОНЯРНЪММНЕ ОПХЯСРЯРБХЕ СБЕКХВХБЮЕР ОПНХГБНДЯРБН ЮМРХРЕК. щРНР ЩТТЕЙР БНГМХЙЮЕР АКЮЦН- ДЮПЪ,ОН ЙПЮИМЕИ ЛЕПЕВЮЯРХВМН,БШЯБНАНФДЕМХЧКХЛ- ТНЙХМНБ (РЮАК. 10.3). р-ЯСОПЕЯЯНПШ (я08-ОНГХРХБМШЕ) НЙЮГШБЮЧР ОПНРХБНОНКНФМНЕ БКХЪМХЕ, БШГШБЮЪ ЯМХФЕМХЕ ХЛЛСММНЦН НРБЕРЮ; ЯХКЭМНЕ ОНДЮБКЕМХЕ НРБЕРЮ ЛНФЕР АШРЭ НДМХЛ ХГ ЛЕУЮМХГЛНБ, КЕФЮЫХУ Б НЯМНБЕ РНКЕПЮМРМНЯРХ. нДМХЛ ХГ ДНОНКМХРЕКЭМШУ ПЕЦСКХПСЧЫХУ ЛЕУЮМХГЛНБ ЪБКЪЕРЯЪ БШПЮАНРЙЮ ЮМРХ-ХДХНРХ- ОНБ (Р.Е. ЮМРХРЕК ОПНРХБ ЯНАЯРБЕММШУ ЮМРХРЕК (ЮСРНЮМРХРЕК)). оПЕДОНКЮЦЮЕРЯЪ, ВРН ОПХ ХЛЛСММНЛ НРБЕРЕ ОПНХГБНДЯРБН ЯОЕЖХТХВЕЯЙНЦН ЮМРХРЕКЮ НАЪГЮРЕКЭМН ЯНОПНБНФДЮЕРЯЪ ОПНХГБНДЯРБНЛ БРНПНЦН ЮМРХРЕКЮ (ЮМРХ-ХДХНРХОМНЦН) ЯН ЯОЕЖХТХВМНЯРЭЧ ОПНРХБ БЮПХЮАЕКЭМШУ (V) ОНЯКЕДНБЮРЕКЭМНЯРЕИ (ХДХНРХОНБ ХКХ ЮМРХЦЕМ-ЯБЪГШБЮЧЫХУ СВЮЯРЙНБ) ОЕПБНЦН ЮМРХРЕКЮ. юМРХХДХНРХОМНЕ ЮМРХРЕКН ЯОНЯНАМН Й ПЮЯОНГМЮБЮМХЧ ХДХНРХОНБ МЮ ЮМРХЦЕММНЛ ПЕЖЕОРНПЕ б-ЙКЕРНЙ (ЙНРНПШИ ОНЯРПНЕМ ХГ ХЛЛСМНЦКНАСКХМЮ, ХДЕМРХВМНЦН ОН ЯРПНЕМХЧ ХДХНРХОС ОЕПБНЦН ЮМРХРЕКЮ),РЮЙХЛ НАПЮГНЛ НМН ЙНМЙСПХПСЕРЯЮМРХЦЕМНЛ Х ЯКСФХРДКЪ ХМЦХАХПНБЮМХЪ ЮЙРХБЮЖХХ б-ЙКЕРЙХ.
еЫЕ ОН РЕЛЕ хллсмнцкнаскхмш (юмрхрекю):
- яРПНЕМХЕ ХЛЛСМНЦКНАСКХМНБ.
- 3. тСМЙЖХХ НРДЕКЭМШУ СВЮЯРЙНБ ЛНКЕЙСКШ ХЛЛСМНЦКНАСКХМЮ
- рЕПЮОХЪ ХЛЛСМНЦКНАСКХМЮЛХ
- яНДЕПФЮМХЕ ХЛЛСМНЦКНАСКХМНБ
- нОПЕДЕКЕМХЕ СПНБМЪ ЯШБНПНРНВМШУ ХЛЛСМНЦКНАСКХМНБ.
- яШБНПНРНВМШЕ ХЛЛСМНЦКНАСКХМШ
- яЕЛЕИЯРБН ХЛЛСМНЦКНАСКХМНБ
- нОПЕДЕКЕМХЕ ХЛЛСМНЦКНАСКХМНБ
- 2.2.2 юМРХРЕКЮ
- яНДЕПФЮМХЕ НАЫХУ ХЛЛСМНЦКНАСКХМНБ ю, G, л ЙКЮЯЯНБ
- пЕЮЙЖХХ Я ХЯОНКЭГНБЮМХЕЛ ЛЕВЕМШУ ЮМРХРЕК
- юМРХОПНРПНЛАХМНБШЕ ЮМРХРЕКЮ
- нЯКНФМЕМХЪ ОПХ ОПХЛЕМЕМХХ ЯШБНПНРНЙ, ХЛЛСМНЦКНАСКХМНБ Х ОКЮГЛШ
- оПЕОЮПЮРШ ЛНМНЙКНМЮКЭМШУ ЮМРХРЕК
- лерндш нопедекемхъ юмрхрек.