Общие данные о никотиновых ацетилхолиновых рецепторах: структура, подтипы, локализация
Структурно-функциональные исследования нАХР начались несколько десятилетий назад благодаря наличию в природе богатого источника этого рецептора - электрического органа скатов Torpedo (T.
marmorata или T. californica), а также высокоактивному и селективному блокатору этого рецептора - а-бунгаротоксину из яда крайта Bungarus multicinctus. Именно такое сочетание позволило охарактеризовать нАХР из Torpedo, представляющий собой олигомерный белок с молекулярной массой в 220 кДа, состоящий из 5 субъединиц (двух а1- и по одной pi-, у- и 5-). Благодаря доступности природного нАХР Torpedo именно этот рецептор в течение многих лет широко исследовался во многих лабораториях мира. Все 5 субъединиц располагаются в мембране вокруг ионного канала, проходящего по центральной оси. Два лиганд-связывающих участка холинергических агонистов и конкурентных антагонистов при этом располагаются в областях контакта больших N-концевых внеклеточных доменов двух а1- и соседних с ними у- и 5-субъединиц рецептора примерно на половине их высоты. Подробную информацию о никотиновых ацетилхолиновых рецепторах можно найти в недавних обзорах (Tsetlin et al., 2009; Taly et al., 2009).Для нАХР из электрического органа ската Torpedo marmorata имеется крио- электронно-микроскопическая структура с разрешением 4А (Unwin, 2005). На схеме (рис. 1) хорошо виден наружный лиганд-связывающий домен, трансмембран-
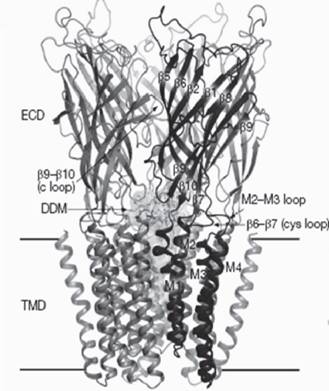
Рис. 1. Структура нАХР Torpedo marmorata по данным криоэлекгронной микроскопии LBD - наружный лиганд-связывающий домен, TMD - трансмембранный домен, CYD- цитоплазматический домен. Под каждой структурой указан ее код в PDB-банке
ный домен, а также часть цитоплазматического домена. Следует напомнить, что в никотиновых рецепторах каждая субъединица между трансмембранными фрагментами М3 и М4 имеет большую цитоплазматическую петлю размером около 150 аминокислотных остатков, которая в наименьшей степени консервативна, и в ней преобладает неупорядоченная структура (Kukhtina et al., 2006).
Последнее обстоятельство, по-видимому, и не позволяет пока добиться кристаллизации нАХР Torpedo или какого-либо иного Cys-петельного рецептора млекопитающих. Другим препятствием служит Cys-петля, которая чрезвычайно гидрофобна в рецепторах млекопитающих - из-за этого не удалось закристаллизовать и их изолированные лиганд-связывающие домены, тогда как существенно меньшая гидрофобность Cys-петли в АХСБ была одним из решающих факторов успешной кристаллизации.По фармакологическим и структурным характеристикам нАХР Torpedo относят к холинорецепторам мышечного типа. Рецепторы из мышечных тканей млекопитающих содержат в своем составе у-субьединицу только на первых этапах развития организма, а у взрослых форм она замещается на е-субьединицу. Прямых данных о пространственной структуре мышечного нАХР пока нет, но многочисленные косвенные данные свидетельствуют о сходной с рецептором Torpedo «пента-субъеди- ничной» организации ионного канала.
С использованием а-бунгаротоксина позднее в мозге млекопитающих был обнаружен новый «нейрональный» нАХР (Salvaterra, Mahler, 1976) - гомоолигомерный а7 подтип рецептора, состоящий из 5 одинаковых а7-субьединиц. В нервных тканях были обнаружены и другие подтипы нАХР. К настоящему времени известны 9 нейрональных а-субъединиц (а2— а10) и 3р-субъединицы (Р2-Р4), которые образуют как гомоолигомерные формы нАХР (из а7-, также из a8- или я9-субъеди- ниц), так и гетеромерные (состоящие из различных комбинаций а- и Р-субъединиц). Наиболее распространенными в нервных тканях являются а4р2, а7, а3абр2, нАХР и некоторые другие подтипы. Пространственные структуры нейрональных нАХР пока не установлены.
Нейрональные нАХР были обнаружены и в пресинаптических областях (в отличие от нАХР мышечного типа, выявленных только в постсинаптической мембране нервно-мышечного сочленения), где они регулируют выброс везикул с различными нейротрансмиттерами. Субъединицы нейрональных нАХР были обнаружены и в самых разных «не-нейрональных» тканях и органах (клетках иммунной системы, легких, коже) (Conti-Fine et al., 2000; Sciamanna et al., 1997; Kumar, Meizel, 2005), где они могут быть сопряжены с рядом патологических состояний. Нарушение работы некоторых подтипов нАХР может вызывать или быть следствием ряда заболеваний, таких как мышечные дистрофии (миастении) (Vincent et al., 2000І и некоторые виды эпилепсии (Steinlein, 2004) и шизофрении (Ripoll et al., 2004). а7нАХР, экспрессированные на пресинаптических мембранах нейронов и на астроцитах, участвуют в высвобождении таких нейромедиаторов, как глутамат и у-аминомасляная кислота (Alkondon et al., 1996; MacDermott et al., 1999), они подавляют экспрессию и встраивание в мембраны NMDA-рецепторов. В этом, вероятно, большую роль играет высокая проницаемость этих рецепторов для кальция - важного внутриклеточного сигнального агента.
3.
Еще по теме Общие данные о никотиновых ацетилхолиновых рецепторах: структура, подтипы, локализация:
- Никотиновые ацетилхолиновые рецепторы: физиологические функции и роль в нейродегенеративных процессах
- Ацетилхолиновый рецептор
- ОБЩИЕ ДАННЫЕ
- ОБЩИЕ ДАННЫЕ
- Общая структура ионотропных глутаматных рецепторов
- ОБЩИЕ ДАННЫЕ
- Общие данные
- ОБЩИЕ ДАННЫЕ О ПЕРЕЛОМАХ
- НЕКОТОРЫЕ ОБЩИЕ ДАННЫЕ ПО ПАЦИЕНТАМ С ГЕМОГЛОБИНОПАТИЕЙ SS
- ОБЩИЕ ДАННЫЕ О ПАТОЛОГИИ НЕРВНОЙ СИСТЕМЫ
- Общие принципы хирургических вмешательств при опухолях различной локализации
- 1.1. Фармакологическая характеристика и данные о влиянии барбитуратов на структуры ЦНС
- Никотиновые холинорецепторы