Ингибиторы «переходного состояния»
Существует много аналогов, препятствующих действию ферментов, занимая место нормального субстрата или кофермента и лишая их способности взаимодействовать с этим ферментом. Таким образом они блокируют нормальные биохимические реакции этого фермента.
Если, согласно Э. Фишеру, описывать соответствие субстрата и фермента как «ключ к замку», то в данном случае можно сказать, что ингибитор будет несоответствующим ключом, не способным открыть замок и в то же время препятствующим введению нужного ключа. Диссоциация комплекса ингибитора с ферментом обычно протекает быстро, поэтому трудно установить, произошло ли только образование первичного комплекса ингибитор — фермент или этот комплекс перешел в нестабильное промежуточное соединение путем образования ковалентной связи между субстратом и ферментом[2]. Рентгеноструктурный анализ позволяет разрешить эту проблему при условии, что имеются источники чистых ферментов. Дляметотрексата, который прочно связывается с дигидрофолатре- дуктазой, установлено, что сокристаллизация фермента, кофермента и субстрата не приводит к образованию ковалентной связи (разд. 9.33). Высокий индекс ингибирования, характерный для метотрексата, объясняется тем, что он более эффективно, чем субстрат, взаимодействует с активным центром фермента.
С этим же явлением связана способность некоторых ингибиторов вызывать более сильные конформационные изменения фермента, чем те, которые происходят при действии субстрата. Так кинетика образования связи между аллопурннолом (9.51)—антиметаболитом гипоксантина (9.52), и ксантиноксида- зой указывает на то, что этот процесс происходит медленно, но в результате образуется прочная связь; устойчивое промежуточное соединение не зафиксировано [Cha, Agarwal, Parks, 1975]. Авторы полагают, что подобное течение реакции свидетельствует о сильных конформационных изменениях.
Рассмотрим ингибиторы, которые создаются по аналогии не с самим субстратом, а с его переходным состоянием. Цель такой работы — получение ингибиторов, способных образовывать значительно более прочные связи с ферментом. Интерес к ним возник в связи с широко распространенным мнением о том, что активный центр свободного фермента комплементарен переходному состоянию субстрата, а не свободному субстрату [Pauling, 1948].
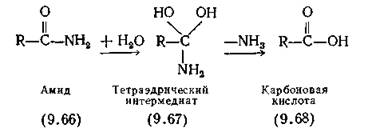
Во многих случаях неизвестно, какое именно переходное состояние образуется при соединении субстрата с ферментом. Поэтому обычно предполагают, что это состояние эквивалентно переходному состоянию, установленному для аналогичной реакции, проводимой в небелковом окружении в присутствии неорганического катализатора, такого как кислота или щелочь. Рассмотрим некоторые общие случаи. При гидролизе амидной (или пептидной) связи на первой стади происходит присоединение молекулы воды по двойной связи С=О с образованием тетраэдрического аддукта. Так, при взаимодействии амида (9.66) с водой происходит образование интермедиата (9.67), который быстро диссоциирует до соответствующей карбоксильной кислоты (9.68). Другим примером может служить конденсация первичного амина с соединением, содержащим альдегидную группу (9.69) (как в пиридоксале) с образованием нестабильного интермедиата (9.70) и последующей его диссоциацией до азоме- тина (основания Шиффа) (9.71).
60
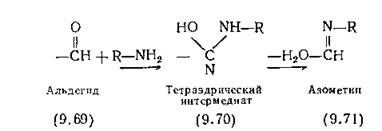
Пентостатин (4.18) (дезоксикоформицин) и его рибозидный аналог коформицин обладают сильным ингибирующим действием по отношению к аденозиндеаминазе, ферменту, превращающему аденозин (9.72) в инозин (9.74) предположительно через интермедиат (9.73). Эту реакцию, подобную гидролизу амидина с образованием амида, необходимо ингибировать при лечении больных препаратами, содержащими адениновый фрагмент, например видарабином (4.16). Молекула пентостатина (4.18) по структуре напоминает молекулу интермедиата (9.73) в области протекания реакции.
Пентостатин и коформицин связаны с ферментом очень прочно, практически необратимо [Cha, Agarwal, Parks 1975].
Точно так же 3,4,5,6-тетрагидроуридин (9.76) — сильный и специфичный ингибитор реакции дезаминирования цитидина (9.75) цитидиндеаминазой [Carmenier, 1968]. Прочная связь, образуемая этим ингибитором с ферментом, объясняется его структурным сходством с тетраэдрическим интермедиатом, подобным (9.75). Аналогично 6-гидроксиметил-1,6-дигидропурин. (9.77)—сильный ингибитор аденозиндезаминазы [Evans, Wol- fenden, 1970].
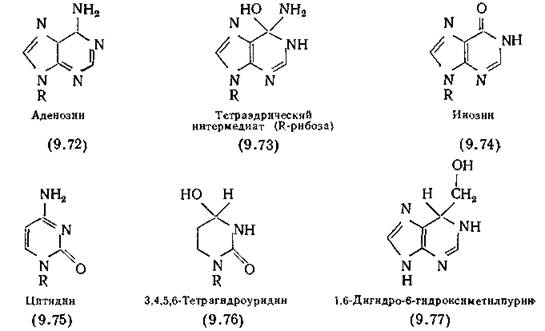
Борные кислоты обладают сильным ингибирующим действием на пептидазы за счет стабильности тетраэдрической структуры, образованной гидроксильными группами, что делает их аналогами нестабильных интермедиатов, образующихся при гидролизе пептидной связи (9.67). Соединение (9.78) специфично действует на химотрипсин, так как необходимым условием для субстрата химотрипсина служит наличие ароматического ядра в боковой цепи [Koehler, Lienhard, 1971].
Альдегиды также представляют собой сильнодействующие ингибиторы пептидазы и находятся в состоянии равновесия с тетраэдрической гидратированной структурой, например (9.79). При этом, чем больше электроноакцепторных заместителей содержит молекула, тем сильнее равновесие смещено в сторону образования метиленгликоля. Некоторые организмы вырабатывают альдегиды в качестве специфичных ингибиторов протеолитических ферментов. Так, например, лейпептины (типичный представитель — ацетил-Ь-лейцил-Ь-лейцилангиналь), вырабатываемые по меньшей мере 17 видами Streptomyces, эффективно ингибируют плазмин, трипсин и папаин, но не химотрипсин. При пероральном введении мышам лейпептины оказывают противовоспалительное действие, кроме того, они обладают радиозащитными свойствами, ингибируют свертывание крови и возникновение опухолей [Umezava, 1972].
Механизм действия ингибиторов «переходного состояния» другого типа связан с карбанионами и карбениевыми катионами.
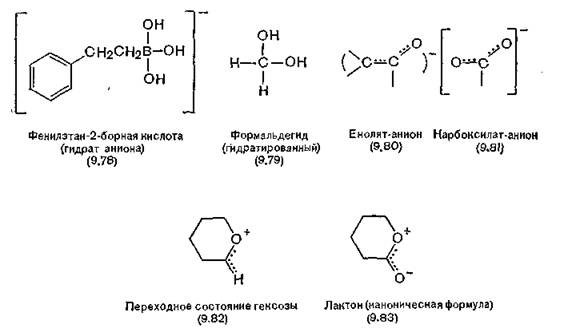
Установлено, что аналоги карбоксилат-иона (9.81) способны эффективно ингибировать некоторые ферментативные реакции, в которых переходное состояние, по-видимому, напоминает енолят-анион (9.80). Таким же образом лактоны, аналогичные по структуре субстрату гексозе, являются сильно действующими ингибиторами большинства гликозилтрансфераз. Переходное состояние в этих реакциях напоминает алкоксикарбениевыйион (9.82), который, так же как и лактон (9.83), имеет конформацию полукреола и положительный заряд на атоме кислорода в цикле.
Было высказано предположение, хотя и недостаточно обоснованное, о том, что при взаимодействии пенициллина с пеп- тидогликантранспептидазой (разд. 13.1) сначала происходит ингибирование по типу «переходного состояния», а затем уже ацилирование антибиотика ферментом [Lee, 1971]. Некоторые 6-дигидро-8-азапурины показали высокие терапевтические индексы при действии на опухоли мышей [Albert, 1980]. Эти экспериментальные препараты разрабатывались как ингибиторы «переходного состояния» биосинтеза пуринов. Вероятно, такие ингибиторы «переходного состояния» имеют большое будущее, однако достижения в этой области пока невелики.
Подробнее об ингибиторах «переходного состояния» см. Lindquist (1975) и Lienhard (1972). В некоторых случаях существует второе место связывания необратимо действующих ингибиторов ферментом (разд. 9.7).
9.6.
Еще по теме Ингибиторы «переходного состояния»:
- Оценка действия ингибиторов АПФ на клиническое состояние и проаритмический статус больных инфарктом миокарда
- Виды переходных пояснично-крестцовых позвонков:
- 3.3.3 Переходные экономики
- Переходно-клеточная папиллома
- 2. Переходно-клеточный рак.
- Формирование признакового пространства по переходным характеристикам биоматериала
- Построение линейной параметрической модели переходной характеристики биоматериала
- Переходно-клеточный рак
- Переходный пояснично-крестцовый позвонок
- Программно-аппаратный комплекс для исследования переходных характеристик в аномальных зонах электропроводимости биоматериала
- Конкуренция в условиях экономики переходного периода
- Ферментативно-активируемые необратимые ингибиторы (ФАНИ)