Бронхиальная астма и механизмы защиты органов дыхания от неблагоприятных воздействий
Генеральная задача экологической пульмонологии заключается в изыскании средств, способствующих преодолению неблагоприятных воздействий химических и физических факторов природного, техногенного и бытового происхождения [Величковский Б.Т., 1995].
Заболевания органов дыхания являются одной из наиболее важных проблем в педиатрии, поскольку до настоящего времени, несмотря на достигнутые успехи, они занимают одно из первых мест в структуре детской заболеваемости - по данным официальной статистики, 50-73% [Domachowske J.B., Rosenberg H.F., 1999; Rodriguez W.J., 1999; Ottolini M.G., Hemming V.G., 1997]. Высокий уровень распространенности острых респираторных инфекций среди детей - важная социальная проблема, решение которой является одной из основных задач здравоохранения в педиатрии.
Известно, что дети, часто болеющие острыми респираторными инфекционными заболеваниями, составляют группы риска по развитию острых бронхитов, бронхиолитов, формированию рецидивирующих бронхитов, включая обструктивные формы, и хронической бронхолегочной патологии. Самой распространенной формой, особенно среди детей в возрасте 1-3 лет, являются бронхиты [Таточенко В.К., 1990; Артамонов Р.Г., Самсыгина Г.А., Зайцева О.В., Корнюшин М.А., 2000; Каганов С.Ю., 1999; Гавалов С.М., 1999]. Заболеваемость бронхитами, по данным ряда авторов, колеблется от 15 % до 50 %. Этот процент существенно повышается (до 50-90%) у детей, часто болеющих острыми респираторными заболеваниями, особенно в экологически неблагоприятных регионах [Почивалов А.В., 1998]. При этом у превалирующего большинства детей наблюдается обструктивный синдром.
Чувствительность рецепторов бронхов усиливается при вирусной инфекции, сопровождающейся повреждением эпителия слизистой оболочки дыхательных путей. Повышенная чувствительность бронхиального дерева к вирусной инфекции может быть связана с аллергической предрасположенностью и гиперреактивностью бронхов [Таточенко В.К., 2000].
Повторные острые респираторные заболевания могут способствовать сенсибилизации организма и создавать предпосылки для развития генерализованных реакций повышенной чувствительности, с последующим формированием рецидивирующих обструктивных бронхитов, БА и других бронхолегочных заболеваний. Выявлена взаимосвязь респираторных заболеваний с обострением БА и приоритетная роль гриппа и респираторно-синтициальной вирусной инфекции в развитии обострения БА [Таточенко В.К., 2000, Мизерницкий Ю.Л., 1992; Геппе Н.А., Каганов С.Ю, 1998].Суммарная распространенность всех обструктивных заболеваний в отдельных регионах России составляет 12,2 на 1000 детей [Почивалов А.В., 1998]. Синдром бронхиальной обструкции наблюдается у 5-40 % детей, госпитализированных с ОРВИ [Артамонов Р.Г., 1987; Каганов С.Ю., 1998].
Эпидемиологические исследования последних лет с использованием современных критериев диагностики позволяют считать, что рецидивирующие бронхиты, особенно обструктивные формы, являются манифестацией бронхиальной астмы [Артамонов Р.Г., 1992; Геппе Н.А., Каганов С.Ю., 1998].
Независимо от способа проникновения чужеродных веществ в клетки, путем ли фагоцитоза или пиноцитоза и диффузии, одним из наиболее биологически значимых продуктов их метаболизма являются свободные радикалы [Pavia D., Bateman J.R.M., Clarke S.W., 1980]. Защита органов дыхания от неблагоприятных воздействий
осуществляется несколькими взаимосвязанными системами и механизмами. К важнейшим из них относятся:
• система конденционирования вдыхаемого воздуха (нагревание, охлаждение, увлажнение);
• система самоочищения респираторного тракта (механическая очистка воздуха на поверхности верхних дыхательных путей и бронхов и удаление бронхиального секрета, содержащего экзогенные включения);
• система самоочищения дыхательной поверхности альвеол и неспецифической
бактерицидной защиты легких:
система метаболизма и обезвреживания токсических веществ и липофильных эндогенных макромолекул;
• система специфической иммунной защиты от инфекционных возбудителей, чужеродных макромолекул и аутоантигенов [Baosly R., et al., 1993].
Все системы поддержания гомеостаза в органах дыхания характеризуются высокой эффективностью. Так, механизмы самоочищения легких от пылевых частиц удаляют 9798% осевшей в них пыли. Микроорганизмы и пылевые частицы, при дыхании осевшие на слизистой трахеобронхиального дерева, быстро и эффективно выводятся наружу. Пыль, отложившаяся в альвеолах, может оказаться на мукоцилиарном эскалаторе дыхательных путей только при помощи фагоцитоза [Bames P.J., 1994]. Фагоцит, таким образом, объединяет функции выведения и бактерицидной защиты, а явление фагоцитоза, открытое И.И.Мечниковым в 1883 году, представляет собой одно из важнейших звеньев невосприимчивости к инфекционным болезням. Молекулярные основы бактерицидности фагоцитоза уточняются вплоть до наших дней. Важнейшую роль в этом процессе играет кислородзависимая бактерицидная система фагоцитов, генерирующая АФК. Первоначально пыль переносится альвеолярными макрофагами в лимфоидные скопления устьев терминальных бронхиол, то есть не столько выводится, сколько концентрируется в
тех местах, где не мешает осуществлению газообмена. Лишь затем развертывается следующая стадия процесса, приводящая к удалению пыли из легких [Baosly R., et al., 1993]. Тем самым не нарушается аэродинамика воздушных потоков в альвеолярной области и снижается опасность развития “болезней малых дыхательных путей” - бронхиолитов и др. [Чучалин А.Г., 1998].
При контакте возбудителя инфекции, иммунного комплекса или пылевой частицы с мембраной фагоцита так интенсивно повышается уровень потребления кислорода клеткой, что это явление получило название “дыхательного взрыва”.
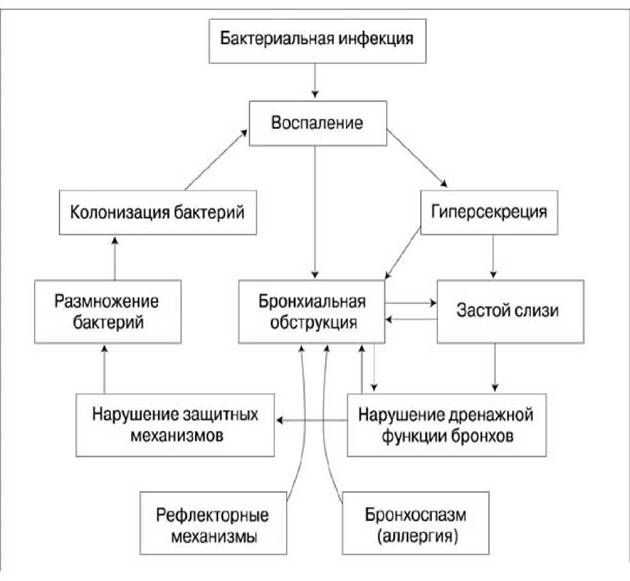
Рис. 18. Взаимосвязь этиопатических факторов и механизмов в формировании и поддрежании хронического воспаления в бронхах (“порочный круг”)
Например, при активации макрофагов пылевыми частицами кварца потребление кислорода увеличивается в 4 раза, частицами диоксида титана — в 1,5 раза.
Еще больше возрастает потребление кислорода при активации макрофагов иммунными комплексами. И практически весь дополнительно поглощенный кислород не используется ни на энергетические, ни на пластические потребности клетки. Основная же часть поглощенного кислорода и энергетических ресурсов клетки расходуется на два параллельно идущих процесса: генерацию бактерицидных радикалов и работу ионных насосов, восстанавливающих ионное и осмотическое равновесие. По указанной причине происходит значительное и быстрое снижение уровня энергетической “валюты” клетки — АТФ.Поэтому хотя общее потребление кислорода гипертрофированной клеткой увеличивается, на единицу массы оно уменьшается. В результате со временем поступление кислорода к митохондриям становится недостаточным, в клетке развитивается энергодефицитное состояние. Внутриклеточная гипоксия нарушает режим работы клеток, в митохондриях начинается усиленное образование супероксидного анион-радикала, вторично нарушающее структуру и работу митохондрий и ведущее к быстрой гибели кониофага. Изменение электронной структуры молекулы кислорода, превращая его в главное оружие бактерицидной защиты клетки — АФК осуществляет особая ферментная система фагоцитов, встроенная во внешнюю клеточную мембрану, НАДФН-оксидаза.
Таким образом, первичный ответ организма на действие как микробного, так и пылевого аэрозоля стереотипен и заключается в мобилизации кислородзависимой бактерицидной системы фагоцитов [Уонер А., 1994].
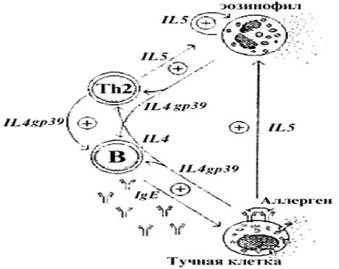
Рис. 19. Т-хелперы 2 типа (Th2), тучные клетки и эозинофилы с помощью схожего профиля цитокинов (IL4, IL5, IL13) и мембраносвязанных молекул (gp39) и секретируемых B лимфоцитами IgE антител создают сеть взаимонаправленных позитивных сигналов, поддерживающих аллергическое воспаление.
На поглощенный корпускулярный объект в фаголизосомах воздействуют высоко химически АФК и молекулы друг их бактерицидных соединений. Затем нежизнеспособный микроорганизм “переваривается” концентрированной смесью лизосомальных ферментов и осуществляется представление (процессинг) выделенного антигена на наружную поверхность мембраны макрофага.
Однако если поглощенный объект — пылевая частица или микроорганизм — к этим влияниям нечувствителен, то длительно и в избыточном количестве образующиеся в процессе фагоцитоза АФК могут стать причиной развития ряда патологических изменений [Pryor W.A., Stone K.,1993].Так, при взаимодействии с белками АФК вызывают окислительную модификацию их антигенных свойств. Этот механизм возникновения аутоантигенов отличается от хорошо известного способа образования путем присоединения гаптена и приводит к развитию аутоиммунного процесса [Spector N.H., 1996]. Последний в своей основе является иммунофизиологическим механизмом и лишь при высокой интенсивности и продолжительности появления аутоантигенов либо наследственной предрасположенности организма становится причиной развития аутоиммунных патологических изменений. Более того, АФК, частично инактивируя и изменяя соотношение протеаз и ингибиторов протеаз в легочной ткани, создают относительную недостаточность а-1-антитрипсина, чреватую опасностью развития эмфиземы и обструктивного синдрома.
При контакте с ДНК радикальные продукты нарушают ее структуру и становятся виновниками появления мутагенных эффектов и развития злокачественных новообразований. При этом гиперобразование АФК является одной из основных причин развития асептического воспаления в органах дыхания.
Обезвреживание чужеродных химических веществ, проникающих в клетки не с помощью фагоцитоза, а путем диффузии и пиноцитоза (жидкие токсические вещества, а также растворимые пары и газы) в органах дыхания осуществляется на обонятельном, бронхиолярном и альвеолярном уровнях. Высокой ферментативной активностью в
процессах обезвреживания обладает апикальная часть обонятельного эпителия и клетки узелков Боумана. В респираторном тракте биотрансформация осуществляется эпителиальными клетками Кларка, количество которых возрастает в терминальных бронхиолах. В альвеолах в процессах обезвреживания задействованы альвеолярные макрофаги, лейкоциты и пневмоциты второго типа.
В самом общем виде систему обезвреживания химических веществ в организме можно представить как состоящую условно из трех звеньев: биотрансформации, конъюгации и антиоксидантной защиты.Задача первой фазы обезвреживания — биотрансформации — заключается в повышении растворимости поглощенных молекул, что облегчает их выведение из организма. Главная роль в реакциях первой фазы принадлежит ферментам монооксигеназам, основным компонентом которых является цитохром Р-450. При этом происходит образование не только нетоксичных, но и токсичных метаболитов, в том числе АФК.
Во второй фазе детоксикации как нетоксичные, так и токсичные метаболиты в результате ферментативной реакции конъюгации присоединяют глюкуроновую кислоту или другие акцепторы. Образующиеся растворимые соединения выводятся из организма. Однако токсичные метаболиты успевают частично взаимодействовать и с другими эндогенными макромолекулами — с ДНК, оказывая на них канцерогенное влияние, с белками, выступая в роли гаптенов и придавая им свойства химических аллергенов техногенной природы.
Третье звено системы обезвреживания ксенобиотиков включает в себя ферментативные и неферментативные средства защиты от токсического действия АФК, возникающих как на первых этапах обезвреживания, так и образующихся под влиянием веществ, обладающих прооксидантными свойствами — озона, оксидов азота и других. Выход высокотоксичных АФК при этом может быть столь значительным, что именно ими определяются пагубные последствия и клинические проявления воздействия исходного ксенобиотика [Тиунов Л.А., 1987].
В основе возникновения гиперреактивности бронхов при бронхиальной астме лежит такой же механизм инактивации фермента АФК, что и при развитии обструктивного синдрома. Не случайно у больных бронхиальной астмой в выдыхаемом воздухе повышено содержание пероксида водорода и оксида азота. Избыток их в легких способен частично инактивировать ферменты, оказывающие влияние на тонус гладкой мускулатуры бронхов.
Весь вопрос в том, что это за фермент? Наиболее подходящим на подобную роль представляется Na, К-АТФаза гладкомышечных клеток бронхов. Частичная инактивация данного фермента приведет к повышению содержания ионов натрия в цитозоле миоцитов и появлению обусловленной этой причиной гиперчувствительности к констрикторным и, напротив, резистентности к дилататорным влияниям [Чучалин А.Г., 1998].
Воспалительный процесс при БА затрагивает все структуры стенки бронха: эпителиальный покров, базальную мембрану, бронх-ассоциированные лимфоидные узелки, сосуды, гладкие мышцы, причем каждая из указанных структур реагирует по- своему. На аутопсийном материале показана большая степень десквамации эпителиальных клеток, которые вместе с эозинофилами и бронхиальным секретом выполняют просвет дыхательных путей [Wilder R.L., 1995]. Считают, что десквамация эпителиальных клеток происходит под действием протеинов, синтезирующихся гранулами эозинофилов и их высвобождение наступает при дегрануляции последних [Величковский Б.Т., 1995]. Базальная мембрана сильно изменена: отечна, утолщена на всем протяжении, основное вещество дезорганизовано. Серозные и бокаловидные клетки гипертрофированы, соотношение эпителиальных и бокаловидных клеток изменено в пользу последних. Сосудистая проницаемость венул и капилляров стенки бронхов заметно изменена. Описываемая морфологическая картина в значительной степени специфична для бронхиальной астмы и свидетельствует о том. что в основе болезни лежит воспаление, персистирующий характер которого подчеркнут и в определении болезни [Мануйлов Б.М., 1994].
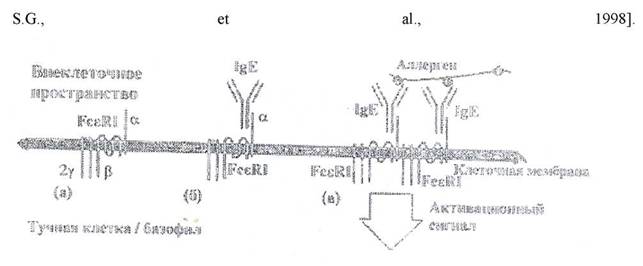
Основным иммунологическим маркером сенсибилизации при БА является повышение общего уровня ІдЕ в сыворотке крови больного и наличие специфических ІдЕ антител к экзоаллергенам. В норме концентрация ІдЕ очень низкая и составляет у здоровых людей примерно 0,00005 г/л. что приблизительно в 100000-200000 раз ниже нормального уровня lgG. ІдЕ фиксируется своим Fc-фрагментом к специфическим рецепторам. На поверхности тучных клеток и базофилов экспрессирован высокоаффинный рецептор к Fc фрагменту молекулы ІдЕ, так называемый Fc-эпсилон рецептор 1 типа или Fc-эпсилон-RI [Abdelilah
Рис. 20. Структура и функция высокоаффинного ІдЕ рецептора.
Существенная часть обшего ІдЕ фиксирована на тучных клетках и базофилах. В отличие от других рецепторов, которые соединяются с fc-эпсилон регионом антител регионом антител других класов только тогда, когда они сцеплены с антигеном, Fc-эпсилон-Rl приклепляет мономерную молекулу свободного ІдЕ. Однако это не приводит к его активации. Для активации рецептора и передачи сигнала несущей его клетке необходима агрегация Fc-эnсилон-Rl. Она достигается при распозновании фиксированными к рецептору молекулами ІдЕ поливалентного антигена (аллергена). Кроме Fc- эпсилон-Rl существует второй тип рецептора, сродство которого к ІдЕ примерно в 100 раз ниже, чем у рецептора 1 типа. Это так называемый низкоаффинный рецептор ІдЕ: Fc-эnсилон-Rll. Существует две формы этого рецептора, алфа и бета ІдЕ: Fc-эnсилон-Rllb экспрессируется главным образом зрелыми В- клетками и моноцитами под воздействием интерлейкина (JL)-4. Fc-эпсилон-Rll существует в растворимой форме, являщейся лигандом CD21, увеличивающим синтез В-лимфоцитами ІдЕ.
(a) F-эпсилон-RI состоит из целей ; (б) альфа цепь связывает мономерный ІдЕ; (в) активация тучной клетки происходит при агрегации с F-эпсилон-RI за счет агрегации ІдЕ при распозновании поливалентного аллергена.
Сигнальный каскад очень быстро, в течение одной минуты приводит к акгивации тучной клетки (базофила) и секреции преформированных и вновь образущихся биологически активных веществ — медиаторов аллергии [Giembycz M.A., Lindsay M.A., 1999]. Этот феномен благодаря морфологической перестройке клетки получил название дегрануляции тучной клетки или базофила [Desreumaux P., Carpon M., 1996]. В результате активации тучной клетки происходит высвобождение серии уже имеющихся липидных соединений (гистамина), а также образующихся вновь медиаторов, к которым относится простагландины Е2и F2-a, тромбоксан А2, а также фактор активации тромбоцитов и лейкотриены (LT) — LTB4, LTD4, LTC4, LTE4 (табл. 3) [Fishman S., Hobbs K., Borish l., 1996; Giembycz M.A., Lindsay M.A., 1999].
Таблица 3. Основные медиаторы воспаления при бронхиальной астме
| Основные медиаторы тучных клеток и базофилов | |
| Медиаторы Преформированные: | Фармакологические эффекты |
| Гистамин | Повышает сосудистую проницаемость, сокращает гладкую мускулатуру бронхов. усиливает продукцию слизи |
| Нейтральные протеазы (триптаза. Химотриптаза, карбоксипротеаза) | Роль при аллергии не ясна: различные популяции клеток продуцируют разные ферменты |
| Синтезируемы de novo: Липидные: | |
| LTC4, LTD4. LTE4 | Повышает сосудистую проницаемость, сокращает гладкую мускулатуру бронхов. усиливает продукцию слизи |
| LTB4 | Хемотаксис нейтрофилов |
| ІПЕ2/ІП F2a | Сокращает гладкую мускулатуру бронхов |
| ТХА2 | Вызывает вазоконстрикцию, сокращает гладкую мускулатуру бронхов, агрегацию тромбоцитов |
| PAF | Повышает сосудистую проницаемость, сокращает гладкую мускулатуру бронхов, вызывает хемотаксис и активацию эозинофилов и нейтрофилов, агрегацию тромбоцитов |
| Цитокины: | |
| Ил-4 | Стимуляция гуморального (Th2) и ингибиция клеточного (Th1) иммунного ответа. переключение В-клеток на синтез ІдЕ, стимуляция экспрессии VCAM-1 эндотелиальными клетками, активация фибробластов |
| Ил-5 | Активация эозинофилов |
| a-TNF | Стимуляция экспрессии ІСАМ-1 эндотелиальными клетками |
| Сокращения: ПГЕ2 /ПГБ2(— пристатдндины Е2 и F2), лейкотриены : LTB4. 1-ТС4, LTD4. LTE4 — лейкотриены В4. LTC4. LTD4 и LTE4: ТХА2 - тромооксан А2, PAF — фактор активации тромбоцитов, Ил — интерлейкин: Th1. Th2 — Т хелперы 1 и 2 типов; ICAM -I молекулы адгезии: TNF-альфа фактор некроза опухоли альфа | |
С действием этих медиаторов связывают немедленный ответ — острый бронхоспазм,
отек слизистой бронхов, гиперсекцию бронхиальной слизи при астме. Эти же медиаторы (главным образом фактор активации тромбоцитов [Kagoshima M, et al., 1997] и LTB 4 [Kajita T., et al, 1985]) ответственны, в свою очередь, за продолжительную активацию эозинофилов, нейтрофилов, тромбоцитов и Т-лимфоцитов, формирование воспаления в тканях и развитие отсроченной реакции [Giembycz M.A., Lindsay M.A., 1999]. У больных БА повышенный уровень лейкотриенов обнаружен в плазме, бронхиальном секрете, бронхоальвеолярной лаважной жидкости. Показана выраженная корреляция уровней LTC4 и LTD4, высвобождаемых из стимулированных лейкоцитов in vitro и тяжестью заболевания при БА у детей и подростков [Kajita T., et al, 1985; Hirata K, Maghni K., Borgeat P., Sirois P. , 1990 ].
Наименее ясным является вопрос, что именно при аллергии и БА создает "сигнальный фон" приводящий к начальному коммутированию Th0 клеток в Th2 направлении. Тучные
клетки могут играть активную регулирующую роль в формировании Th2/lgE опосредованного иммунного ответа при бронхиальной астме [Martin RJ., 1995].
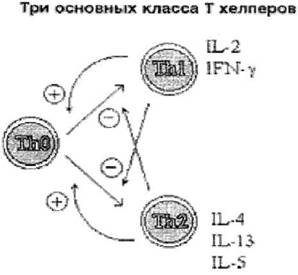
Рис. 21. Три основных класса Т-хелперов (CD4+). Цитоксииы Th1 клеток способствуют дифференцировке Th0 в Th1 и подавляют развитие Th2, цитотоксины Th2, напротив способоствуют дифференцировке Th2 и подавляют Th 1.
Однако, учитывая необходимость первичной сенсибилизации тучных клеток, можно предположить, что их регулирующая активность сводится к усилению Th2 зависимого иммунного ответа и привлечению клеток - посредников воспаления (эозинофилов) [Desreumaux P., Carpon M., 1996].
Регуляция активности эозинофилов в бронхах, по-видимому, зависит в первую очередь от Т-лимфоцитов (Th2) и таких их цитокинов, как Ил-5, GM-CSF, Ил-3 [Hessel E.M., et al, 1998]. Источником этих цитокинов, кроме Th2 клеток, могут быть тучные клетки и сами эозинофилы (аутокринная регуляция) [Dutton P.W., Swain S.L., 1999; DeSanctics G.T., et al., 1997]. Биологически активные продукты эозинофилов кроме повреждающего могут оказывать регулирующее действие. В дополнение к уже разобранным эффектам следует отметить возможность секреции Ил-8; он специфически вызывает хемотаксис нейтрофилов. Кроме того, эозинофилы могут активировать тучные клетки и, возможно, фибробласты. CD4+ лимфоцит может спонтанно приобретать Th2 фенотип. В работах последних двух лет показана возможность секреции Th0 клеткой в нейтральных условиях Ил-4, приводящего к спонтанной дифференцировке в лимфоцит, продуцирующий Th2 профиль цитокинов. Таким образом, сам Т-лимфоцит может выступать в роли клетки, инициирующей формирование Th2/lgE варианта иммунного ответа [Giembycz M.A., Lindsay M.A., 1999]. Экстрацеллюлярные антигены, по-видимому,
привлекают другие типы клеток, которые могут быть источником Ил-4. Ил-4, как уже упоминалось, дифференцирует пролиферирующие CD4+ Т-лимфоциты в направлении Th2 клеток, подавляет их дифференцировку в Th1 клетки и, по-видимому, является ключевым цитокином в организации синтеза ІдЕ (в эксперименте мыши с разрушенным геном, кодирующим Ил-4 оказались неспособными продуцировать ІдЕ). Среди CD8+ лимфоцитов есть клетки, вырабатывающие Th1 и Th2 подобный профиль цитокинов [Daser A., Meissner N., Herz U., 1995]. Последние гипотетически могут играть роль как в регуляции дифференцировки Th0 в Th2. так и в привлечения эозинофилов к месту аллергического воспаления [Giembycz M.A., Lindsay M.A., 1999].
При БА эозинофилы часто обнаруживаются в большом количестве в жидкости бронхоальвеолярного лаважа, а также в биопсийном и аутопсийном материале. Они определяются не только во время обострения, но и в ремиссии заболевания. При обострении заболевания выявляются признаки активации эозинофилов и повышенной секреции ими медиаторов (рис. 20, 21), среди которых следует выделить высокотоксичные основные белки, такие как главный основной протеин (major basic protein - МБР) и эозинофильный катионный протеин (eosinophil cationic protein - ECP) [Giembycz M.A., Lindsay M.A., 1999]. Уровень эозинофильного катионного белка в биологических средах организма является в настоящее время одним из самых информативных маркеров аллергического воспаления.
Острая аллергическая реакция наступает в течение нескольких минут после контакта с причинным аллергеном и длится 30-40 минут. В изолированном виде она встречается у некоторого количества больных БА. В большинстве случаев отмечается более сложная и комплексная реакция на провокацию: после нескольких (3-12) часов благополучного периода наступает постепенное ухудшение бронхиальной проходимости, получившее название реакции поздней фазы или отсроченной реакции.
Реакция поздней фазы отмечается в ответ на ингаляционную провокацию причинным аллергеном у 60-70% больных БА, развивших немедленную реакцию. С одной стороны, наличие реакция поздней фазы жестко ассоциировано с характерным признаком бронхиальной астмы — феноменом бронхиальной гиперреактивности. С другой стороны, реакция поздней фазы характеризуется появлением уже описанных выше признаков воспалительных изменений со стороны слизистой оболочки дыхательных путей, с инфильтрацией воспалительными клетками. Отмечается второй пик повышения концентрации гистамина не сопровождающийся, однако, повышением концентрации
простагландинов и триптазы, что указывает на его базофильное происхождение. Отмечается повышение уровня продуктов эозинофилов — МБР, ЕСР, цитокин Ил-5. Специфическая иммунотерапия и противовоспалительная терапия кортикостероидами эффективны главным образом в отношение выраженности реакции поздней фазы и не оказывают существенного влияния на немедленную реакцию [Kaiser J., Bickel C.A., Bochner B.S., Schleimer R.P., 1993].
Таким образом, выявляется связь между четырьмя характеристиками болезни: клиникой БА, бронхиальной гиперреактивностью, наличием воспалительных изменений в дыхательных путях и реакцией поздней фазы. Комплексное влияние некоторых биологически активных соединений во время реакции поздней фазы хорошо согласуется с находками в воспалительном очаге. Например. Ил-4 может вызывать адгезию эозинофилов, но не нейтрофилов, а Ил-5 в сочетании с Ил-3 и GM-CSF (гранулоцитарномоноцитарный колониестимулирующий фактор) активирует эозинофилы, поступившие в ткань [Kameyoshi Y. et al., 1992]. Кроме того, некоторые представители новой группы соединений, хемокинов, такие как М1 Р-1 альфа и RANTES (макрофагального и эпителиального происхождения) [Kakazu T., Chihara J., Saito A., Nakajima S., 1995], судя по экспериментальным данным также могут рекрутировать и аккумулировать эозинофилы в дыхательных путях [Чучалин, 1998]. Точное представление об участии молекул адгезии в аллергическом воспалении позволяет рассматривать их как потенциальную терапевтическую мишень при лечении аллергии и астмы [Abdelaziz M.M., et al, 1997].
Вторичная недостаточность антиоксидантных ферментов, выявляющаяся при бронхиальной астме, может приводить к увеличению уровня свободных радикалов (рис. 22), образующихся в процессе НАДФН оксидазного пути метаболизма кислорода:
О2'—>О2' (супероксид)—> Н2О2(перекись водорода) —> НО' (гидроксильный радикал) —> Н2О
Кроме прямого повреждающего действия на клетки тканей дыхательных путей, АФК, по-видимому, способны индуцировать секрецию хемокинов, что приводит к усилению инфильтративного компонента воспаления.
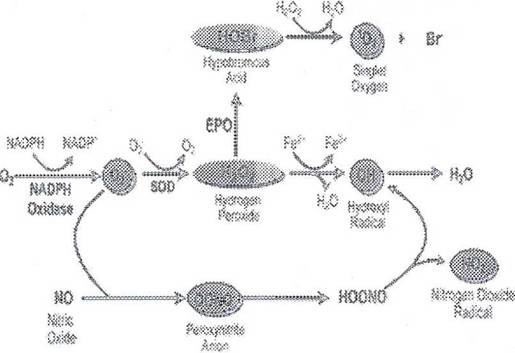
Рис. 22. Механизмы образования свободных радикалов в эозинофилах и других фагоцитах в результате активации НАДФН-оксидазы с генерацией супероксидного аниона [Giembycz M.A., Lindsay M.A., 1999].
1.5.1.
Еще по теме Бронхиальная астма и механизмы защиты органов дыхания от неблагоприятных воздействий:
- Тема 3. Дифференциальная диагностика и лечение бронхиальной обструкции (бронхиальная астма).
- Глава 7. Бронхиальная астма
- Бронхиальная астма
- Бронхиальная астма
- Бронхиальная астма
- Бронхиальная астма
- Бронхиальная астма
- 1.2. Неблагоприятное воздействие лекарственной терапии на плод
- Бронхиальная астма у детей, проживающих в условиях санитарно-гигиенического неблагополучия
- Бронхиальная астма
- Бронхиальная астма
- Бронхиальная астма
- 20. Гиподинамия как фактор неблагоприятного воздействия на здоровье.
- Протоколы диагностики и лечения хронического гастродуоденита у детей, ассоциированного с неблагоприятным воздействием химических факторов среды обитания