хйянднбше йкеых (Ixodidae)
б ТЮСМЕ ХЙЯНДНБШУ ЙКЕЫЕИ яНБЕРЯЙНЦН яНЧГЮ МЮЯВХРШБЮЕРЯЪ 66 БХДНБ, ОПХМЮДКЕФЮЫХУ Й 6 ПНДЮЛ. пЮЯОПНЯРПЮМЕМШ ХЙЯНДНБШЕ ЙКЕЫХ МЮ БЯЕУ ЛЮРЕ╜ПХЙЮУ ГЕЛМНЦН ОРЮПЮ, Б ПЮГМННАПЮГМШУ КЮМДЬЮТРМШУ ГНМЮУ.
б яяяп МЮХАНКЭЬЕЕ ВХЯКН БХДНБ ХЙЯНДНБШУ ЙКЕЫЕИ БЯРПЕВЮЕРЯЪ МЮ ЧЦЕ. щРНЛС АКЮЦНОПХЪРЯРБСЕР РЕОКШИ ЙКХЛЮР, ПЮГМННАПЮГХЕ ПЮЯРХРЕКЭМШУ ЦПСО╜ОХПНБНЙ, НАХКХЕ ОПНЙНПЛХРЕКЕИ.
йКЕЫХ ЯЕЛЕИЯРБЮ Ixodidae ЪБКЪЧРЯЪ БПЕЛЕММШЛХ ЙПНБНЯНЯСЫХЛХ ОЮПЮ╜ГХРЮЛХ МЮГЕЛМШУ ОНГБНМНВМШУ. б НЯМНБМНЛ НМХ ОЮПЮГХРХПСЧР МЮ ЛКЕЙНОХ╜РЮЧЫХУ, ПЕФЕ МЮ ОРХЖЮУ Х ПЕДЙН МЮ ПЕОРХКХЪУ. оХРЮМХЕ МЮ УНГЪХМЕ ДКХРЯЪ МЕЯЙНКЭЙН ДМЕИ, ОПХ ЩРНЛ НМХ ЯОНЯНАМШ ОНЦКНЫЮРЭ АНКЭЬНЕ ЙНКХВЕЯРБН ЙПНБХ, ЙНРНПНИ ХЛ УБЮРЮЕР ДКЪ ПЮГБХРХЪ ЪХЖ.
оНКНБНГПЕКШЕ ХЙЯНДНБШЕ ЙКЕЫХ ЙПСОМНЦН ПЮГЛЕПЮ (ЦНКНДМШЕ ЯЮЛЙХ ≈ 3≈9 ЛЛ, ЦНКНДМШЕ КХВХМЙХ 0,77≈1 ЛЛ, ЦНКНДМШЕ МХЛТШ ≈ 0,96 ≈ 1 ЛЛ Б ДКХМС).
рЕКН ЯНЯРНХР ХГ ХДХНЯНЛШ (ЯНАЯРБЕММН РЕКЮ) Х ЦМЮРНЯНЛШ (УНАНРЙЮ). яНАЯРБЕММН РЕКН ОПЕДЯРЮБКЪЕР ЯНАНИ ЯКХРМШИ ЛЕЬНЙ. уНАНРНЙ ОНДБХФМН ЯНВКЕМЕМ Я РЕКНЛ Х ПЮЯ╜ОНКНФЕМ МЮ ОЕПЕДМЕЛ ЙНМЖЕ РЕКЮ, БХДЕМ ЯБЕПУС С НЯНАЕИ БЯЕУ ТЮГ ПЮГБХРХЪ. с ОНКНБН╜ГПЕКШУ ЙКЕЫЕИ Х МХЛТ ВЕРШПЕ ОЮПШ МНЦ, С КХВХМНЙ ≈ РПХ. оНЙПНБ ЙНФХЯРШИ. мЮ ЯОХМ╜МНИ ЯРНПНМЕ БЯЕЦДЮ ХЛЕЕРЯЪ ЫХРНЙ: С ЙКЕЫЕИ-ЯЮЛЖНБ МЕЙНРНПШУ ПНДНБ ЕЯРЭ Х АПЧЬМШЕ ЫХРЙХ. яОХММНИ ЫХРНЙ С ЯЮЛЖНБ ОНЙПШБЮЕР БЯЧ ЯОХММСЧ ОНБЕПУМНЯРЭ. с ЯЮЛНЙ, КХВХ╜МНЙ, МХЛТ НМ ГЮМХЛЮЕР РНКЭЙН ОЕПЕДМЧЧ ВЮЯРЭ РЕКЮ. нДМЮ ОЮПЮ ДШУЮРЕКЭМШУ НРБЕП╜ЯРХИ С ЙКЕЫЕИ ПЮЯОНКНФЕМЮ МЮ ОКЮЯРХМЙЮУ ≈ ОЮПХРПЕЛЮУ ОН АНЙЮЛ РЕКЮ ОНГЮДХ НЯМН╜БЮМХЪ IV ОЮПШ МНЦ. цКЮГЮ ВЮЯРН ХЛЕЧРЯЪ Х КЕФЮР МЮ АНЙНБШУ ЙПЮЪУ ЯОХММНЦН ЫХРЙЮ. с ЯЮЛНЙ МЮ НЯМНБЮМХХ УНАНРЙЮ ПЮЯОНКНФЕМШ ОНПНБШЕ ОНКЪ.
лЕДХЖХМЯЙНЕ ГМЮВЕМХЕ. хЙЯНДНБШЕ ЙКЕЫХ НАПЮРХКХ МЮ ЯЕАЪ БМХЛЮМХЕ Б МЮВЮКЕ XX БЕЙЮ Б ЯБЪГХ Я ХГСВЕМХЕЛ ЦЕЛНЯОНПХДХНГНБ ДНЛЮЬМХУ ФХБНР╜МШУ Х МЕНАУНДХЛНЯРЭЧ ПЮГПЮАНРЙХ ОПНТХКЮЙРХВЕЯЙХУ ЛЕПНОПХЪРХИ ДКЪ АНПЭАШ Я ЩРХЛХ ГЮАНКЕБЮМХЪЛХ.
лЕДХЖХМЯЙНЕ ХУ ГМЮВЕМХЕ ЯРЮКН ХГБЕЯРМН Б 30-Е ЦНДШ БН БПЕЛЪ БШЪЯМЕ╜МХЪ ХУ ПНКХ Б ЩОХГННРНКНЦХХ Х ЩОХДЕЛХНКНЦХХ ВСЛШ, РСКЪПЕЛХХ, Ю ГЮРЕЛ ЛЮПЯЕКЭЯЙНИ КХУНПЮДЙХ Х ЙКЕЫЕБНЦН ЯШОМНЦН РХТЮ юГХХ.
хЯЯКЕДНБЮМХЕ ХЙЯНДНБШУ ЙКЕЫЕИ НЯНАЕММН ЬХПНЙН АШКН ОНЯРЮБКЕМН ОНЯКЕ БШЪБКЕМХЪ ХУ ПНКХ ЙЮЙ ОЕПЕМНЯВХЙНБ Х УПЮМХРЕКЕИ БХПСЯЮ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ МЮ дЮКЭМЕЛ бНЯРНЙЕ, Ю БОНЯКЕДЯРБХХ Б ДПСЦХУ ПЮИНМЮУ ЯРПЮМШ.пЮЯЯЛНРПЕМХЕ ПНКХ ХЙЯНДНБШУ ЙКЕЫЕИ Б ЩЙНМНЛХЙЕ КЧДЕИ ГЮЯКСФХБЮЕР НЯНАНЦН БМХЛЮМХЪ, РЮЙ ЙЮЙ Б СЛЕПЕММШУ Х РПНОХВЕЯЙХУ ЯРПЮМЮУ НМХ ОПЕБНЯ╜УНДЪР БЯЕУ ВКЕМХЯРНМНЦХУ ОН ВХЯКС ГЮАНКЕБЮМХИ, ОЕПЕМНЯХЛШУ ВЕКНБЕЙС Х ДНЛЮЬМХЛ ФХБНРМШЛ. йЮЙ ОЕПЕМНЯВХЙХ ГЮАНКЕБЮМХИ ВЕКНБЕЙС НМХ ЯРНЪР МЮ БРНПНЛ ЛЕЯРЕ Х СЯРСОЮЧР Б ЩРНЛ НРМНЬЕМХХ РНКЭЙН ЙНЛЮПЮЛ.
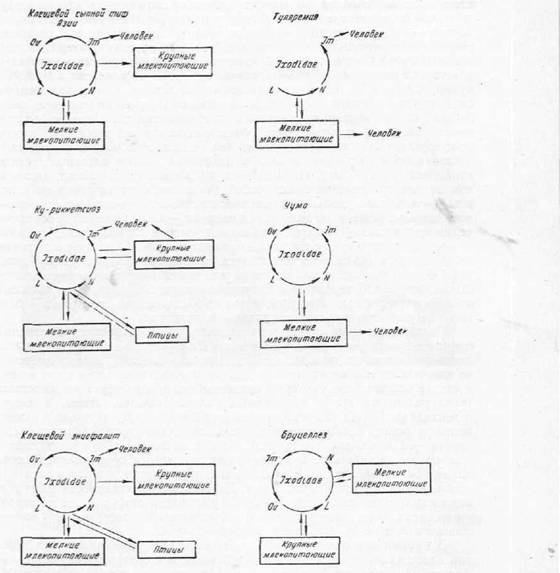
б РЕВЕМХЕ ЯБНЕИ ФХГМХ ЙКЕЫХ ЯНЯСР ЙПНБЭ ОНГБНМНВМШУ ФХБНРМШУ РПХФДШ, Р. Е. НДХМ ПЮГ Б ЙЮФДНИ ТЮГЕ ПЮГБХРХЪ. мЮ ВЕКНБЕЙЮ НАШВМН МЮОЮ╜ДЮЧР ОНКНБНГПЕКШЕ НЯНАХ (Ixodes persulcatus, Dermacentor silvarum, Der- macentor nuttail Ё Х ДП.), ПЕФЕ ОНКНБНГПЕКШЕ ЙКЕЫХ Х МХЛТШ (Haemaphy- salis concinna, Ixodes ricinus), КХВХМЙХ Х МХЛТШ (Ixodes Iaguri Iaguri). оПХЙКЕОКЪЧРЯЪ Й РЕКС ВЕКНБЕЙЮ: МЮ ЯОХМЕ, ФХБНРЕ, ЦПСДХ; Й МНЦЮЛ ≈ (АЕДПС), ЬЕЕ, ЦНКНБЕ. сЙСЯШ ЙКЕЫЕИ ВСБЯРБХРЕКЭМШ ДКЪ ВЕКНБЕЙЮ, МН ХМНЦДЮ ОПХЯНЯЮММШЕ ЙКЕЫХ НЯРЮЧРЯЪ МЕГЮЛЕВЕММШЛХ ДН ОНКМНЦН МЮЯШЫЕМХЪ (ДН 7 ДМЕИ).
мЮ ЛЕЯРЕ СЙСЯЮ ЙКЕЫЮ С ВЕКНБЕЙЮ ЛНФЕР ПЮГБХБЮРЭЯЪ ЪГБНВЙЮ. вЮЯРН НАПЮГСЕРЯЪ ХМТХКЭРПЮР (I. persulcatus).
лНФМН НРЛЕРХРЭ АНКЭЬСЧ ОПХЯОНЯНАКЕММНЯРЭ НПЦЮМХГЛЮ ЙКЕЫЕИ Й БНГ╜АСДХРЕКЪЛ ЛМНЦХУ ГЮАНКЕБЮМХИ ВЕКНБЕЙЮ. рЕЯМЮЪ ЯБЪГЭ ЯСЫЕЯРБСЕР ЛЕФДС ЙКЕЫЮЛХ Х ПХЙЙЕРЯХЪЛХ, Б ЛЕМЭЬЕИ ЯРЕОЕМХ ЛЕФДС МХЛХ Х БХПСЯЮЛХ Х АЮЙ╜РЕПХЪЛХ. еЫЕ МЕДНЯРЮРНВМН ХГСВЕМН ХУ БГЮХЛННРМНЬЕМХЕ Я ОПНЯРЕИЬХЛХ БНГАСДХРЕКЪЛХ ГЮАНКЕБЮМХИ ВЕКНБЕЙЮ. нПЦЮМХГЛ ЙКЕЫЮ, ОН ХЛЕЧЫХЛЯЪ Б МЮЯРНЪЫЕЕ БПЕЛЪ ЯБЕДЕМХЪЛ, МЕ ЪБКЪЕРЯЪ АКЮЦНОПХЪРМНИ ЯПЕДНИ НАХРЮМХЪ ДКЪ БНГАСДХРЕКЕИ РНЙЯНОКЮГЛНГЮ, КЕОРНЯОХПНГЮ.
б МЕОНЯПЕДЯРБЕММНИ ОЕПЕДЮВЕ ХМТЕЙЖХНММНЦН ЮЦЕМРЮ ВЕКНБЕЙС СВЮЯРБСЕР РНКЭЙН ВЮЯРЭ БХДНБ ХЙЯНДНБШУ ЙКЕЫЕИ. рЮЙ, I. persulcatus ОЕПЕДЮЕР ВЕКНБЕЙС БНГАСДХРЕКЪ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ; D. nuttalli ≈ БНГАСДХРЕКЪ ПХЙЙЕРЯХН- ГНБ, D. pictus ≈ РСКЪПЕЛХЧ Х Р. Д.
дПСЦХЕ БХДШ ЙКЕЫЕИ ЮЙРХБМН ПЮЯОПНЯРПЮМЪЧР БНГАСДХРЕКЪ КХЬЭ Б ЙПСЦС ЕЯРЕЯРБЕММШУ ОПНЙНПЛХРЕКЕИ ≈ ДХЙХУ ОНГБНМНВМШУ ФХБНРМШУ.
щРН НРЛЕ╜ВЕМН БН ЛМНЦХУ ОПХПНДМШУ НВЮЦЮУ йС-КХУНПЮДЙХ, РСКЪПЕЛХХ, ВСЛШ.гМЮВЕМХЕ ХЙЯНДНБШУ ЙКЕЫЕИ-ОЕПЕМНЯВХЙНБ МЕ НЦПЮМХВХБЮЕРЯЪ РНКЭЙН ЯОНЯНАМНЯРЭЧ ОЕПЕДЮБЮРЭ БНГАСДХРЕКЪ ВЕКНБЕЙС Х ФХБНРМШЛ Б ОПНЖЕЯЯЕ ЙПНБНЯНЯЮМХЪ. вЮЯРН ХУ ПНКЭ НЯНАЕММН БЕКХЙЮ ЙЮЙ ЯРНИЙХУ УПЮМХРЕКЕИ БНГ╜АСДХРЕКЕИ Б ОПХПНДЕ.
рЮЙ, ПХЙЙЕРЯХХ ЛЮПЯЕКЭЯЙНИ КХУНПЮДЙХ, ЙКЕЫЕБНЦН ЯШОМНЦН РХТЮ юГХХ, ОЪРМХЯРНИ КХУНПЮДЙХ яЙЮКХЯРШУ ЦНП яью ЯНУПЮМЪЧРЯЪ Б ОНКН╜БНГПЕКШУ ЙКЕЫЮУ ОНФХГМЕММН, ОЕПЕДЮЧРЯЪ Б ОПНЖЕЯЯЕ ЛЕРЮЛНПТНГЮ Х ВЕ╜ПЕГ ЪИЖН ДНВЕПМЕЛС ОНЙНКЕМХЧ. сЯРЮМНБКЕМЮ РПЮМЯТЮГНБЮЪ Х РПЮМЯ╜НБЮПХЮКЭМЮЪ ОЕПЕДЮВЮ БНГАСДХРЕКЪ РЮЙФЕ ДКЪ ОЕПЕМНЯВХЙНБ ЙКЕЫЕБНЦН ЩМЖЕ╜ТЮКХРЮ.
Atnonie БХДШ ЙКЕЫЕИ ЪБКЪЧРЯЪ ЕЯРЕЯРБЕММШЛХ МНЯХРЕКЪЛХ ДБСУ Х АНКЕЕ БХДНБ БНГАСДХРЕКЕИ АНКЕГМЕИ ВЕКНБЕЙЮ; МЮОПХЛЕП, I. ricinus Х Dermacentor pictus ХГБЕЯРМШ ЙЮЙ МНЯХРЕКХ БНГАСДХРЕКЕИ 6 ХМТЕЙЖХИ, Dermacentor mar- ginatus ≈ 7 МР. Д.
оПХ ЩРНЛ НРДЕКЭМШЕ БХДШ ЙКЕЫЕИ ПЮГКХВМН НРМНЯЪРЯЪ Й РНЛС ХКХ ХМНЛС БНГАСДХРЕКЧ АНКЕГМХ. нЯНАЕММН ЩРН ОПНЪБКЪЕРЯЪ Б НРМНЬЕМХХ ЛЕФДС ЙКЕЫЮЛХ Х БНГАСДХРЕКЪЛХ МЕЙНРНПШУ АЮЙРЕПХЮКЭМШУ ХМТЕЙЖХИ. оНКСВЕМШ ОНКНФХРЕКЭМШЕ ПЕГСКЭРЮРШ Н ДКХРЕКЭМНЛ ЯНУПЮМЕМХХ Б ЙКЕЫЮУ БНГАСДХРЕ╜КЕИ РСКЪПЕЛХХ, ВСЛШ Х АПСЖЕККЕГЮ. мН КХЬЭ Б НРДЕКЭМШУ ЯКСВЮЪУ МЮАКЧ╜ДЮЕРЯЪ ЯОНЯНАМНЯРЭ РПЮМЯТЮГНБНИ Х РПЮМЯНБЮПХЮКЭМНИ ОЕПЕДЮВХ P. pestis (I. Crenulatus, Rh. schulzei).
щОХДЕЛХНКНЦХВЕЯЙНЕ ГМЮВЕМХЕ НРДЕКЭМШУ БХДНБ ХЙЯНДНБШУ ЙКЕЫЕИ ПЕГЙН БЮПЭХПСЕР Б ЯБЪГХ Я ПЮГКХВМНИ ХУ ЮЦПЕЯЯХБМНЯРЭЧ ОН НРМНЬЕМХЧ Й ВЕКН-
БЕЙС (ВЮЯРНРНИ МЮОЮДЕМХЪ Х ОПХЯЮЯШБЮМХЪ) Х ЯОНЯНАМНЯРЭЧ ЕЦН ХМТХЖХПНБЮРЭ ОПХ ЙПНБНЯНЯЮМХХ.
б ЛЕЯРЮУ ПЮЯОПНЯРПЮМЕМХЪ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ Я МЮКХВХЕЛ ОЕПЕМНЯ╜ВХЙЮ I. persulcatus ГЮПЮФЕМХЕ КЧДЕИ ОПНХЯУНДХР Б ПЕГСКЭРЮРЕ ОПХЯЮЯШБЮМХЪ ЩРНЦН БХДЮ ЙКЕЬЮ. бЯКЕДЯРБХЕ АНКЭЬНИ ЮЦПЕЯЯХБМНЯРХ НМ ЪБКЪЕРЯЪ НВЕМЭ ЮЙРХБМШЛ ОЕПЕМНЯВХЙНЛ ДЮММНЦН ГЮАНКЕБЮМХЪ. б РН ФЕ БПЕЛЪ ЙКЕЫ I. riciπus Б НВЮЦЮУ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ ХЛЕЕР АНКЭЬЕЕ ГМЮВЕМХЕ ЙЮЙ ПЕГЕПБСЮП БХПС╜ЯЮ ОН ЯПЮБМЕМХЧ Я ЕЦН ОЕПЕМНЯВХЙНЛ. гЮПЮФЕМХЕ КЧДЕИ Б ЩРХУ НВЮЦЮУ ОПН╜ХЯУНДХР ВЮЯРН ВЕПЕГ ЛНКНЙН ЙНГ.
оЕПЕМНЯВХЙ ЛЮПЯЕКЭЯЙНИ КХУНПЮДЙХ (Rh. sanguineus) ПЕДЙН МЮОЮДЮЕР МЮ ВЕКНБЕЙЮ, ВЕЛ, ОН-БХДХЛНЛС, Х НАЗЪЯМЪЧРЯЪ ЯОНПЮДХВЕЯЙХЕ ЯКСВЮХ ГЮАН╜КЕБЮМХЪ Б ЛЕЯРЮУ ЛЮЯЯНБНЦН ПЮЯОПНЯРПЮМЕМХЪ ЩРНЦН БХДЮ.
оПХПНДМШЕ НВЮЦХ ГЮАНКЕБЮМХИ ВЕКНБЕЙЮ ПЮЯОПНЯРПЮМЕМШ Б НОПЕДЕКЕМ╜МШУ КЮМДЬЮТРЮУ, ЦДЕ МЮХАНКЕЕ АКЮЦНОПХЪРМШ СЯКНБХЪ ДКЪ ХУ ЯСЫЕЯРБНБЮ╜МХЪ. рЮЙ, ЙКЕЫЕБНИ ЩМЖЕТЮКХР ЬХПНЙН ПЮЯОПНЯРПЮМЕМ Б КЕЯМНИ ГНМЕ Х БЕЯЭЛЮ ПЕДЙН БЯРПЕВЮЕРЯЪ Б КЕЯНЯРЕОХ. йКЕЫЕБНИ ЯШОМНИ РХТ юГХХ ХГБЕЯРЕМ Б ЯРЕО МШУ, КЕЯНЯРЕОМШУ Х ОНКСОСЯРШММШУ КЮМДЬЮТРЮУ. лЮПЯЕКЭЯЙЮЪ КХУНПЮДЙЮ МЮАКЧДЮЕРЯЪ ОПЕХЛСЫЕЯРБЕММН Б ОПХАПЕФМНИ ОНКНЯЕ яПЕДХГЕЛМНЛНПЯЙНЦН АЮЯЯЕИМЮ. оПХПНДМШЕ НВЮЦХ йС-КХУНПЮДЙХ ЬХПНЙН БЯРПЕВЮЧРЯЪ Б ОСЯРШМ╜МНИ, ОНКСОСЯРШММНИ Х ЯРЕОМНИ ГНМЮУ Х ЛЮКН ХГБЕЯРМШ Б КЕЯМНИ ГНМЕ. нОРХ╜ЛЮКЭМШЛХ ДКЪ РСКЪПЕЛХХ ЪБКЪЧРЯЪ КЕЯНЯРЕОЭ, ЯРЕОЭ, ОНИЛШ Х ДЕКЭРШ ПЕЙ, ОНАЕПЕФЭЪ НГЕП, АНКНР Х Б ЛЕМЭЬЕИ ЛЕПЕ ≈ ЬХПНЙНКХЯРБЕММШЕ КЕЯЮ.
б ОПЕДЕКЮУ ЙЮФДНИ ГНМШ ПЮЯОПНЯРПЮМЕМХЕ ОПХПНДМШУ НВЮЦНБ ХМТЕЙЖХИ ОПХСПНВЕМН Й НОПЕДЕКЕММШЛ РХОЮЛ АХНЖЕМНГНБ.
б ГЮБХЯХЛНЯРХ НР УЮПЮЙРЕПЮ КЮМДЬЮТРЮ ЛЕМЪЕРЯЪ Х ЯРПСЙРСПЮ НВЮЦЮ, Р. Е. ЯНЯРЮБ ЙНЛОНМЕМРНБ.
нДХМ Х РНР ФЕ БХД ОЕПЕМНЯВХЙЮ, МН НАХРЮЧЫХИ Б ПЮГКХВМШУ КЮМДЬЮТР╜МШУ СЯКНБХЪУ, ХЛЕЕР ЯОЕЖХТХВЕЯЙХИ ДКЪ ЙЮФДНИ ГНМШ ЙНЛОКЕЙЯ ДХЙХУФХБНР- МШУ-ОПНЙНПЛХРЕКЕИ. оН МЮАКЧДЕМХЪЛ я. о. оХНМРЙНБЯЙНИ (1960), Б ОПХПНД╜МШУ НВЮЦЮУ ЙКЕЫЕБНЦН ЯШОМНЦН РХТЮ юГХХ ОЕПЕМНЯВХЙ D. nuttail Ё МЮ ЧЦЕ ЯРЕОМНИ ВЮЯРХ йПЮЯМНЪПЯЙНЦН ЙПЮЪ ХЛЕЕР РЕЯМШЕ АХНРХВЕЯЙХЕ ЯБЪГХ Я ДКХМ╜МНУБНЯРШЛ ЯСЯКХЙНЛ Х СГЙНВЕПЕОМНИ ОНКЕБЙНИ; Б рСБХМЯЙНИ ЙНРКНБХМЕ, ЙПНЛЕ ЯСЯКХЙЮ, ОПНЙНПЛХРЕКЪЛХ ЪБКЪЧРЯЪ ЯХАХПЯЙХИ РСЬЙЮМВХЙ, ДЮСПЯЙХИ УНЛЪВНЙ; Б НОСЯРШМЕММНИ ЯРЕОХ сАЯСМСПЯЙНИ ЙНРКНБХМШ≈2 БХДЮ ОХЫСУ, РСЬЙЮМВХЙ Х 4БХДЮ УНЛЪВЙНБ.
оПХПНДМШЕ НВЮЦХ РНЦН ФЕ ГЮАНКЕБЮМХЪ, МН Я ОЕПЕМНЯВХЙНЛ D. silva- rum ПЮЯОНКНФЕМШ Б КЕЯНЯРЕОХ ОПЕДЦНПХИ (мНБНЯХАХПЯЙЮЪ НАКЮЯРЭ), ПЕВМШУ ДНКХМ (аСПЪРЯЙЮЪ юяяп), Ю РЮЙФЕ МЮ РЕППХРНПХХ БШПСАНЙ ЬХПНЙНКХЯРБЕМ╜МШУ КЕЯНБ, ОНЙПШРШУ ЙСЯРЮПМХЙНЛ (чФМНЕ оПХЛНПЭЕ).
б РЕУ ЯКСВЮЪУ, ЙНЦДЮ ОЕПЕМНЯВХЙНЛ ЪБКЪЕРЯЪ H. concinna, ОПХПНДМШЕ НВЮЦХ ЙКЕЫЕБНЦН ЯШОМНЦН РХТЮ БЯРПЕВЮЧРЯЪ Б ГНМЕ ЙНВЙНБЮРШУ АНКНР АКХГ КЕЯЮ (чФМНЕ оПХЛНПЭЕ), Б ЙСЯРЮПМХЙНБШУ ОНПНЯКЪУ (бНЯРНВМЮЪ яХАХПЭ).
нЯМНБМШЛХ ОПНЙНПЛХРЕКЪЛХ ЩРХУ БХДНБ ЙКЕЫЕИ Б чФМНЛ оПХЛНПЭЕ ЪБКЪЧРЯЪ ЦПШГСМШ: АСПСМДСЙ, ДЮКЭМЕБНЯРНВМЮЪ Х ЙПЮЯМН-ЯЕПЮЪ ОНКЕБЙХ; ОРХЖШ, ОЪРМХЯРШЕ НКЕМХ Х Р. Д. б гЮОЮДМНИ яХАХПХ ≈ НАШЙМНБЕММШИ УНЛЪЙ, Б йПЮЯМНЪПЯЙНЛ ЙПЮЕ ≈ ЯСЯКХЙ, ОНКЕБЮЪ ЛШЬЭ, СГЙНВЕПЕОМЮЪ ОНКЕБЙЮ.
оПХПНДМШЕ НВЮЦХ КХУНПЮДЙХ Ky Б ЯРЕОМНИ ГНМЕ яЕБЕПМНЦН йЮГЮУЯРЮМЮ ЯБЪГЮМШ Я АЕПЕГНБН-НЯХМНБШЛХ ЙНКЙЮЛХ, ОНИЛЮЛХ ПЕЙ, ОНАЕПЕФЭЪЛХ НГЕП, ЦДЕ ЦКЮБМШЛХ ОПНЙНПЛХРЕКЪЛХ ОЕПЕМНЯВХЙЮ D. marginatus ЯКСФЮР: ОНКЕБЙХ (НАШЙМНБЕММЮЪ, ЙПЮЯМН-ЯЕПЮЪ), НАШЙМНБЕММШИ УНЛЪЙ, ЛЮКШИ ЯСЯКХЙ. бДЮКХ НР БНДНЕЛНБ, ЙПНЛЕ ЩРНЦН БХДЮ, ОЕПЕМНЯВХЙНЛ ЪБКЪЕРЯЪ I. Crenulatus, ЯБЪГЮМ╜МШИ Я ОНЯЕКЕМХЪЛХ ЯСПЙНБ. б ОСЯРШММНИ ГНМЕ ОПХПНДМШЕ НВЮЦХ КХУНПЮДЙХ Ky НАМЮПСФЕМШ Б ЛЕЯРЮУ НАХРЮМХЪ АНКЭЬНИ ОЕЯВЮМЙХ, РНМЙНОЮКНЦН ЯСЯКХЙЮ. 23 гЮЙЮГ Nb 228
оПХПНДМШЕ НВЮЦХ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ Я ОЕПЕМНЯВХЙНЛ I. persulcatus УЮПЮЙРЕПХГСЧРЯЪ АНКЭЬХЛ ПЮГМННАПЮГХЕЛ АХНЖЕМНГНБ. б ОНДГНМЕ ЧФМНИ РЮИЦХ Х ГЮОЮДМШУ КЕЯНБ (йЮКХМХМЯЙЮЪ НАКЮЯРЭ. йЮПЕКЭЯЙЮЪ юяяп) ОЕПЕМНЯ╜ВХЙ ОПХСПНВЕМ Й ЕКНБН-ЬХПНЙНКХЯРБЕММШЛ КЕЯЮЛ (ОПНЯЕЙЮЛ, НОСЬЙЮЛ, БШ╜ПСАЙЮЛ 5≈7-КЕРМЕЦН БНГПЮЯРЮ), МЮ РЕППХРНПХХ ЙНРНПШУ ЦКЮБМШЛХ ОПНЙНП╜ЛХРЕКЪЛХ МЕОНКНБНГПЕКШУ ЙКЕЫЕИ ЪБКЪЧРЯЪ ОНКЕБЙХ (НАШЙМНБЕММЮЪ, ЙПЮЯ╜МН-ЯЕПЮЪ), МЮЯЕЙНЛНЪДМШЕ (ГЕЛКЕПНИЙЮ НАШЙМНБЕММЮЪ, ЙСРНПЮ).
б БНЯРНВМНЕБПНОЕИЯЙНИ РЮИЦЕ ЯРНИЙХЕ ОПХПНДМШЕ НВЮЦХ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ ЯСЫЕЯРБСЧР Б ЦКСУХУ Х МЕНАФХРШУ СВЮЯРЙЮУ РЮИЦХ, Ю РЮЙФЕ АКХГ ОНЯЕКЙНБ МЮ ОПХСЯЮДЕАМШУ СВЮЯРЙЮУ Х НЦНПНДЮУ (оЕПЛЯЙЮЪ НАКЮЯРЭ), Б ЯНЯ╜МНБН-ЕКНБШУ КЕЯЮУ Я ОПХЛЕЯЭЧ КХЯРБЕММШУ ОНПНД (яБЕПДКНБЯЙЮЪ НАКЮЯРЭ), ЯЛЕЬЮММШУ ЕКЭМХЙЮУ Х ЯНЯМЪЙЮУ (сДЛСПРЯЙЮЪ юяяп), КХОНБН-ДСАНБШУ Х АЕПЕГНБН-КХОНБШУ ЮЯЯНЖХЮЖХЪУ (рЮРЮПЯЙЮЪ юяяп, йСИАШЬЕБЯЙЮЪ НАКЮЯРЭ), НЯХМНБН-АЕПЕГНБШУ КЕЯЮУ СПЛЮМН-АНКНРХЯРНИ РЮИЦХ (рНЛЯЙЮЪ, нЛЯЙЮЪ НАКЮЯРХ), Б ЯЛЕЬЮММНЛ КЕЯС ОПЕДЦНПХИ, Я ОПЕНАКЮДЮМХЕЛ УБНИМШУ ОНПНД (йЕЛЕПНБЯЙЮЪ НАКЮЯРЭ), Б ВЕПМЕБНИ РЮИЦЕ, Я ОПЕНАКЮДЮМХЕЛ ОХУРЮПМХЙНБ (юКРЮИЯЙХИ ЙПЮИ), Б ЛЕГНТХКЭМШУ ЙЕДПНБН-ЬХПНЙНКХЯРБЕММШУ КЕЯЮУ (дЮКЭ╜МХИ бНЯРНЙ). б ЙЮФДНИ ХГ ОЕПЕВХЯКЕММШУ КЮМДЬЮТРМШУ ПЮГМНЯРЕИ ОЕПЕ╜МНЯВХЙ ХЛЕЕР ЯННРБЕРЯРБСЧЫХИ ЙНЛОКЕЙЯ ОПНЙНПЛХРЕКЕИ ≈ ОНГБНМНВМШУ ФХБНРМШУ.
гМЮВЕМХЕ ДНЛЮЬМХУ ФХБНРМШУ, ЦКЮБМШЛ НАПЮГНЛ ПНЦЮРНЦН ЯЙНРЮ ЙЮЙ ОПНЙНПЛХРЕКЕИ ОНКНБНГПЕКШУ ЙКЕЫЕИ, БЕКХЙН ОНВРХ БН БЯЕУ ПЮИНМЮУ ПЮЯОПН╜ЯРПЮМЕМХЪ ОЕПЕМНЯВХЙЮ. хЯЙКЧВЕМХЕ ЯНЯРЮБКЪЧР МЕЙНРНПШЕ ОПХПНДМШЕ НВЮЦХ дЮКЭМЕЦН бНЯРНЙЮ. б НАПЮГНБЮМХХ СВЮЯРЙНБ БШЯНЙНИ ВХЯКЕММНЯРХ ОНКНБНГ╜ПЕКШУ ЙКЕЫЕИ Б МЕНАФХРНИ РЮИЦЕ Б ЩРХУ ПЮИНМЮУ ХЦПЮЧР БЕДСЫСЧ ПНКЭ ЙПСО╜МШЕ ДХЙХЕ ФХБНРМШЕ.
б КХЯРБЕММХВМН-ЯНЯМНБНЛ КЕЯС ОНИЛШ ПЕЙХ аЕКНМ (хПЙСРЯЙЮЪ НАКЮЯРЭ) Б НВЮЦЕ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ ОПНЙНПЛХРЕКЪЛХ ОНКНБНГПЕКШУ ЙКЕЫЕИ ХГ ДХЙХУ ЛКЕЙНОХРЮЧЫХУ ЪБКЪЧРЯЪ ЙНГСКЪ Х ГЮЪЖ, ПЕФЕ ЛЕДБЕДЭ Х ЛЮПЮК. б РЕЛМНУБНИМНЛ КЕЯС ОПЕДЦНПХИ БДНКЭ АЕПЕЦНБ БЕПУНБХИ ПЕЙ Х ЙКЧВЕИ≈ ЙЮАЮПЦЮ, ГЮЪЖ, ЛЮПЮК, ЛЕДБЕДЭ, ПЕДЙН ЙНГСКЪ, КНЯЭ (я. й. сЯРХМНБ, 1962).
мЕГМЮВХРЕКЭМШИ ЮПЕЮК ГЮМХЛЮЧР ОПХПНДМШЕ НВЮЦХ ЙКЕЫЕБНЦН ЩМЖЕТЮ╜КХРЮ Я ОЕПЕМНЯВХЙЮЛХ H. concinπa (КЕЯНЯРЕОЭ ХКХ ЙСЯРЮПМХЙНБШЕ ГЮПНЯКХ, ЙНВЙНБЮРШЕ АНКНРЮ БАКХГХ КЕЯЮ Б оПХЛНПЯЙНЛ, уЮАЮПНБЯЙНЛ ЙПЮЪУ), H. ja- poπica (ЬХПНЙНКХЯРБЕММШИ КЕЯ Б оПХЛНПЯЙНЛ ЙПЮЕ), D. Silvarum (КСЦНБШЕ ЮЯЯНЖХЮЖХХ БДНКЭ ПЕЙ Х НЙНКН МЮЯЕКЕММШУ ОСМЙРНБ Б РЮИЦЕ оПХЛНПЯЙНЦН Х уЮАЮПНБЯЙНЦН ЙПЮЕБ), D. marginatus (КЕЯНЯРЕОЭ йЮГЮУЯРЮМЮ). оПХПНДМШЕ НВЮЦХ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ, ЦДЕ ПЕГЕПБСЮПНЛ БНГАСДХРЕКЪ ЪБКЪЧРЯЪ I. riciπus, ПЮЯОПНЯРПЮМЕМШ Б ЕБПНОЕИЯЙХУ ЬХПНЙНКХЯРБЕММШУ КЕЯЮУ, ЦКЮБМШЛ НАПЮГНЛ Б НКЭЬЮРМХЙЮУ Х ДСАНБН-ЦПЮАНБШУ КЕЯЮУ.
б ПЪДЕ ЯКСВЮЕБ ЮПЕЮКШ ОЕПЕМНЯВХЙНБ ЬХПЕ РЮЙНБШУ БНГАСДХРЕКЪ. щРН НРМНЯХРЯЪ Й D. pictus (ОПХ РСКЪПЕЛХХ), D. marginatus, H. concinπa (ОПХ ЙКЕ╜ЫЕБНЛ ЯШОМНЛ РХТЕ юГХХ), Rh. sanguineus (ОПХ ЛЮПЯЕКЭЯЙНИ КХУНПЮДЙЕ) Х Р. Д.
бЛЕЯРЕ Я РЕЛ ХГБЕЯРМШ ГЮАНКЕБЮМХЪ, ЮПЕЮК ЙНРНПШУ ГМЮВХРЕКЭМН ЬХПЕ ЮПЕЮКЮ ОЕПЕМНЯВХЙЮ ДЮММНИ ХМТЕЙЖХХ, МЮОПХЛЕП КХУНПЮДЙХ йС. щРН НАЗЪЯ╜МЪЕРЯЪ ХМШЛ ЛЕУЮМХГЛНЛ ОЕПЕДЮВХ БНГАСДХРЕКЪ ВЕКНБЕЙС, ХЯЙКЧВЮЧЫХЛ РПЮМЯЛХЯЯХЧ.
пЮГМНЯРНПНММЕЕ Х ЙНЛОКЕЙЯМНЕ ХГСВЕМХЕ (1936 ≈ 1940) СВЮЯРХЪ ЙКЕ╜ЫЕИ Б ЩОХДЕЛХНКНЦХХ ПХЙЙЕРЯХНГМШУ Х БХПСЯМШУ ГЮАНКЕБЮМХИ Б ЩЙЯОЕДХ╜ЖХНММШУ Х ЩЙЯОЕПХЛЕМРЮКЭМШУ СЯКНБХЪУ БЯЙПШКН ХЯРНВМХЙХ ЯСЫЕЯРБНБЮМХЪ БНГАСДХРЕКЕИ.
нАНАЫЕМХЕ АНКЭЬНЦН ТЮЙРХВЕЯЙНЦН ЛЮРЕПХЮКЮ ЯРЮКН НЯМНБНИ ДКЪ ЯНГ╜ДЮМХЪ е. м. оЮБКНБЯЙХЛ (1933) СВЕМХЪ Н ОПХПНДМНИ НВЮЦНБНЯРХ АНКЕГМЕИ ВЕКНБЕЙЮ, ЙНРНПНЕ ЪБХКНЯЭ ЯРХЛСКНЛ ДКЪ ЬХПНЙНЦН Х БЯЕЯРНПНММЕЦН ХГСВЕ╜МХЪ ХЙЯНДНБШУ ЙКЕЫЕИ Б яяяп, Ю РЮЙФЕ Х БН ЛМНЦХУ ДЕЛНЙПЮРХВЕЯЙХУ ЯРПЮМЮУ.
пХЙЙЕРЯХНГШ. б яНБЕРЯЙНЛ яНЧГЕ ОЕПБШЕ ДНЙЮГЮРЕКЭЯРБЮ МЮКХВХЪ ПХЙ╜ЙЕРЯХНГМШУ ГЮАНКЕБЮМХИ АШКХ ОНКСВЕМШ Б 1936 Ц. ю. ъ. юКШЛНБШЛ, СЯРЮ╜МНБХБЬХЛ Б яЕБЮЯРНОНКЕ ОЕПЕМНЯВХЙЮ ЛЮПЯЕКЭЯЙНИ КХУНПЮДЙХ ЯНАЮВЭЕЦН ЙКЕЫЮ Rhipicephalus sanguineus Latr. щРН ГЮАНКЕБЮМХЕ, БОЕПБШЕ НРЙПШРНЕ Б 1910 Ц. Б рСМХЯЕ [йНМНП Х аПСВ (Conor Х Bruch); йНМНП Х цЮЪР (Conor Х Hayat)], ЬХПНЙН ПЮЯОПНЯРПЮМЕМН Б ЯРПЮМЮУ яПЕДХГЕЛМНЛНПЯЙНЦН АЮЯЯЕИМЮ, вЕПМНЛНПЯЙНЦН Х йЮЯОХИЯЙНЦН АЮЯЯЕИМНБ.
гМЮВЕМХЕ Rh. sanguineus ЙЮЙ ОЕПЕМНЯВХЙЮ Rickettsia conori БОЕПБШЕ ДНЙЮГЮКХ Б 1930≈1931 ЦЦ. дЧПЮМ, аПСЛОР, йНМЯЕИ (Durand, Brumpt1 Conceil) Х аКЮМ, йЮЛХ- Ц.НОЕРПНЯ (Blanc, Caminopetros, 1932). с ДЮММНЦН БХДЮ ЙКЕЫЕИ СЯРЮМНБКЕМЮ РПЮМЯТЮГН╜БЮЪ Х РПЮМЯНБЮПХЮКЭМЮЪ ОЕПЕДЮВЮ ПХЙЙЕРЯХИ [аПСЛОР, 1932; йНЛАХЕЯЙС Х гНРРЮ (яНР- oiescuu Zotta, 1932); йЮЛХМНОЕРПНЯ, 1931).
б яяяп ЕЯРЕЯРБЕММН ГЮПЮФЕММШЕ Rh. sanguineus НАМЮПСФЕМШ БН ЛМНЦХУ МЮЯЕКЕМ╜МШУ ОСМЙРЮУ йПШЛЯЙНЦН Х йЮБЙЮГЯЙНЦН оНАЕПЕФЭЕБ вЕПМНЦН ЛНПЪ (ю. ъ. юКШЛНБ, 1936; н. я. йНПЬСМНБЮ Х я. о. оХНМРЙНБЯЙЮЪ, 1949; о. т. гДПНДНБЯЙХИ Х е. л. цНКХ- МЕБХВ, 1947) Х МЮ ОНАЕПЕФЭЕ йЮЯОХИЯЙНЦН ЛНПЪ (о. о. оНОНБ, 1946).
сЯРЮМНБКЕМН ДКХРЕКЭМНЕ ДЕЯЪРХКЕРМЕЕ CoxpaneHHeR. conori Б ОНОСКЪЖХХ Rh. sanguineus (н. я. йНПЬСМНБЮ Х я. о. оХНМРЙНБЯЙЮЪ, 1949). пЕГЕПБСЮ╜ПНЛ БНГАСДХРЕКЪ ЯПЕДХ ФХБНРМШУ ЪБКЪЕРЯЪ ЯНАЮЙЮ. цХП Х лЕИККНМ (Geer Х Meillon, 1930, 1941) Х цХП (1954) БШДЕКХКХ ЬРЮЛЛШ ПХЙЙЕРЯХИ НР ЯНАЮЙ Б чФМНИ юТПХЙЕ.
оНЯКЕ НАМЮПСФЕМХЪ ГЮПЮФЕММШУ Rh. sanguineus БНГАСДХРЕКЕЛ ЛЮПЯЕКЭ╜ЯЙНИ КХУНПЮДЙХ Б яЕБЮЯРНОНКЕ МЮВЮКХ ЕФЕЦНДМН ОНОНКМЪРЭЯЪ МЮЬХ ЯБЕДЕ╜МХЪ Н ГМЮВЕМХХ ХЙЯНДНБШУ ЙКЕЫЕИ Б ЩОХДЕЛХНКНЦХХ ПХЙЙЕРЯХНГМШУ ГЮАН╜КЕБЮМХИ. -
б 1938 Ц. ЙНЛОКЕЙЯМНИ ЩЙЯОЕДХЖХЕИ бЯЕЯНЧГМНЦН ХМЯРХРСРЮ ЩЙЯОЕПХЛЕМ╜РЮКЭМНИ ЛЕДХЖХМШ (ОНД ПСЙНБНДЯРБНЛ ОПНТ. л. й. йПНМРНБЯЙНИ) Б НЙПЕЯРМН╜ЯРЪУ йПЮЯМНЪПЯЙЮ АШКЮ БШЪБКЕМЮ МНБЮЪ ТНПЛЮ ПХЙЙЕРЯХНГМНЦН ГЮАНКЕ╜БЮМХЪ ≈ ЙКЕЫЕБНИ ЯШОМНИ РХТ. аШКХ БШДЕКЕМШ ЬРЮЛЛШ ПХЙЙЕРЯХИ ХГ D. nuttalli (о. к. яНКХРЕПЛЮМ Х я. о. оХНМРЙНБЯЙЮЪ, 1939), ХДЕМРХВМШЕ ЬРЮЛ╜ЛЮЛ, БШДЕКЕММШЛ БОЕПБШЕ Б 1938 Ц. н. я. йНПЬСМНБНИ ХГ ЙПНБХ АНКЭМШУ ЙКЕ╜ЫЕБШЛ ЯШОМШЛ РХТНЛ. о. т. гДПНДНБЯЙХИ ДЮК МЮГБЮМХЕ ЩРНЛС БНГАСДХРЕ╜КЧ ≈ Derniacentorxenus sibiricus.
б МЮЯРНЪЫЕЕ БПЕЛЪ БШЪБКЕМН ЬХПНЙНЕ ПЮЯОПНЯРПЮМЕМХЕ ЩРНИ ХМТЕЙЖХХ ОН РЕППХРНПХХ ЮГХЮРЯЙНИ ВЮЯРХ. яяяп (НР рЧЛЕМЯЙНИ НАКЮЯРХ ДН чФМНЦН оПХЛНПЭЪ, БЙКЧВЮЪ рСБХМЯЙСЧ юяяп). б 1959 Ц. ПХЙЙЕРЯХХ D. sibiricus БШДЕКЕМШ ХГ ЙКЕЫЕИ Б юПЛЕМХХ (л. е. йНЖХМЪМ).
б ОЕПЕДЮВЕ БНГАСДХРЕКЪ D. sibiricus ВЕКНБЕЙС СВЮЯРБСЧР ЯКЕДСЧЫХЕ БХДШ ЙКЕЫЕИ: Dermacentor nuttalli (о. к. яНКХРЕПЛЮМ Х я. о. оХНМРЙНБЯЙЮЪ, 1938), D. Silvarum (е. о. яЮБХЖЙЮЪ, 1943), D. pictus (я. л. йСКЮЦХМ, н. я. йНП╜ЬСМНБЮ, м. х. юКТЕЕБ, 1947), D. margiπatus (л. й. йПНМРНБЯЙЮЪ, е. о. яЮ╜БХЖЙЮЪ, 1946), Haemaphysalis concinna (3. л. фЛЮЕБЮ, 1941), H. punctata (л. й. йПНМРНБЯЙЮЪ, е. о. яЮБХЖЙЮЪ, 1946), Rh. sanguineus (л. е. йНЖХМЪМ, 1959), I. persulcatus (л. я. ьЮИЛЮМ, 1961).
яОНМРЮММЮЪ ГЮПЮФЕММНЯРЭ D. nuttalli СЯРЮМНБКЕМЮ Б МЕЯЙНКЭЙХУ ПЮИН╜МЮУ ЯРПЮМШ: Б НЙПЕЯРМНЯРЪУ йПЮЯМНЪПЯЙЮ (о. к. яНКХРЕПЛЮМ Х я. о. оХНМР╜ЙНБЯЙЮЪ, 1938), Б гЮАЮИЙЮКЭЕ (е. д. оЕРПЪЕБ, 1946), Б уЮЙЮЯЯЙНИ НАКЮЯРХ Х Б гЮОЮДМШУ яЮЪМЮУ (сЯХМЯЙЮЪ ЙНРКНБХМЮ) (н. я. йНПЬСМНБЮ, я. о. оХНМР╜ЙНБЯЙЮЪ, м. ю. мХЙХРХМЮ, 1959), Б рСБХМЯЙНИ ACCP (я. о. оХНМРЙНБЯЙЮЪ,
23*
н. я. йНПЬСМНБЮ, м. й. лХЫЕМЙН, 1959), D. Silvarum Ъ уЮАЮПНБЯЙНЛ ЙПЮЕ (е. о. яЮБХЖЙЮЪ, 1943), Б йЕЛЕПНБЯЙНИ НАКЮЯРХ (д. т. оКЕЖХРШИ, 1947). Б мНБНЯХАХПЯЙНИ НАКЮЯРХ (л. я. ьЮИЛЮХ, 1957), Б оПХЛНПЯЙНЛ ЙПЮЕ (я.л. йСКЮЦХМ Я ЯНЮБРНПЮЛХ, 1959; й- б. кЕЦЙНДХЛНБЮ, 1963); D. marginatus Б юКРЮИЯЙНЛ ЙПЮЕ (я. л. йСКЮЦХМ, н. я. йНПЬСМНБЮ, м. х. юКТЕЕБ, 1947), Б юКЛЮ-юРХМЯЙНИ НАКЮЯРХ йЮГЮУЯЙНИ CCP (д. я. юПУЮМЦЕКЭЯЙХИ, 1955), Б бНЯРНВМНЛ йЮГЮУЯРЮМЕ (н. я. йНПЬСМНБЮ Х я. о. оХНМРЙНБЯЙЮЪ, 1957), Б хЯЯШЙ-йСКЭЯЙНИ НАКЮЯРХ йХПЦХГХХ (е. о. яЮБХЖЙЮЪ, 1946), Б юПЛЕМХХ (л. е. йНЖХМЪМ, 1959); D. pictus Б мНБНЯХАХПЯЙНИ НАКЮЯРХ (л. я. ьЮИ- ЛЮМ, 1957), Б юКРЮИЯЙНЛ ЙПЮЕ (я. л. йСКЮЦХМ, н. я. йНПЬСМНБЮ, м. х. юК╜ТЕЕБ, 1947); H. conciπna Б чФМНЛ оПХЛНПЭЕ (3. л. фЛЮЕБЮ, 1941; я. л. йСКЮЦХМ Х ДП.. 1959; б. к. кЕЦЙНДХЛНБЮ, 1963), Б уЮАЮПНБЯЙНЛ ЙПЮЕ (е. о. яЮБХЖЙЮЪ, 1943), МЮ НЯРПНБЮУ чФМНЦН оПХЛНПЭЪ (ц. о. яНЛНБ, л. х. ьЮОХ╜ПН, ю. ю. оЕРПНБ, 1957); H. punctata ≈ Б хЯЯШЙ-йСКЭЯЙНИ НАКЮЯРХ йХПЦХ╜ГХХ (л. й. йПНМРНБЯЙЮЪ Х е. о. яЮБХЖЙЮЪ, 1946), Б юКЛЮ-юРХМЯЙНИ НАКЮЯРХ (д. я. юПУЮМЦЕКЭЯЙХИ, 1955).
гЮПЮФЕМХЕ ВЕКНБЕЙЮ ЛЮПЯЕКЭЯЙНИ КХУНПЮДЙНИ, ЙКЕЫЕБШЛ ЯШОМШЛ РХТНЛ ОПНХЯУНДХР Б ОПНЖЕЯЯЕ ЙПНБНЯНЯЮМХЪ ЙКЕЫЕИ-ОЕПЕМНЯВХЙНБ. мН ОПХ ЛЮПЯЕКЭЯЙНИ КХУНПЮДЙЕ ХГБЕЯРМН РЮЙФЕ ХМТХЖХПНБЮМХЕ ВЕПЕГ ЯКХГХЯРШЕ НАНКНВЙХ (НЯНАЕММН ЦКЮГ).
лЮПЯЕКЭЯЙЮЪ КХУНПЮДЙЮ Х ЙКЕЫЕБНИ ЯШОМНИ РХТ НРМНЯЪРЯЪ Й ЦПСООЕ ОЪРМХЯРНИ КХУНПЮДЙХ яЙЮКХЯРШУ ЦНП. яНАЯРБЕММН ОЪРМХЯРЮЪ КХУНПЮДЙЮ (Rocky Mountain Spotted Fever) АШКЮ НРЙПШРЮ Б 1899 Ц. Б НАКЮЯРХ яЙЮКХЯРШУ ЦНП яью Б ЬРЮРЕ лНМРЮМЮ.
б МЮЯРНЪЫЕЕ БПЕЛЪ ЩРН ГЮАНКЕБЮМХЕ ЬХПНЙН ПЮЯОПНЯРПЮМЕМН Б яЕБЕПОНИ юЛЕПХЙЕ Х ХГБЕЯРМН ОНВРХ БН БЯЕУ ЬРЮРЮУ яью [йСКЕИ (Cowlley), сЩККЕП (WheIler, 1957)], ХЛЕЕРЯЪ Б йЮМЮДЕ, лЕЙЯХЙЕ, оЮМЮЛЕ, хМДХХ Х Б ПЮГКХВМШУ ВЮЯРЪУ юТПХЙХ [лЕЦЮС (Megaw). пНДФЕПЯ (Rodgers, 1952)].
б яНЕДХМЕММШУ ьРЮРЮУ Х Б йЮМЮДЕ ХГБЕЯРМН ВЕРШПЕ БХДЮ ХЙЯНДНБШУ ЙКЕЫЕИ, НЯМНБ╜МШУ ОЕПЕМНЯВХЙНБ Rickettsia riskettsi: Derniacentor andersoni [пХЙЙЕРЯ,йХМЦ (Ricketts, King, 1906); йНККЮ (Me. Colla1 1908)], D. Variabilis [дЮИЕП, аЕДФЕП, лЮБЕП (Dyer, Badger, Maver1 1932)], Haemaphysalis Ieporis palustris [оЮПЙЕП (Parker, 1923)]1Ambly- omma Cajennense [йНКГ (Kohls. 1948]). б аПЮГХКХХ, оЮМЮЛЕ ОЕПЕМНЯВХЙНЛ ЩРНЦН БНГАСДХРЕКЪ ЪБКЪЕРЯЪ A. Cajennense [лЕЦЕКЭУЮКЭЯ (Magalhdls, 1949); йНКГ (Kohls, 1948); пНДЮМХЬ (Rodanische1 1953)]. Б яЮМ-оЮСКН ≈ ЙКЕЫХ A. striatum [рПЮБЮЯНЯ (Travassos1 1938)], Б бНЯРНВМНИ юТПХЙЕ ≈ Haemaphysalis Ieachi [дХЙ, кЭЧХЯ (Diek. Lewis, 1948)1. хМТЕЙЖХНММШИ ЮЦЕМР ОЪРМХЯРНИ КХУНПЮДЙХ БШДЕКЕМ ХГ Rb. sanguineus Ъ лЕЙЯХЙЕ [аСЯРЮЛЕМР Х бЩПЕКЭ (Bustamante Х Varela. 1943)1, Ю РЮЙФЕ ХГ Ixodes dentatus [ХХР. ОН юПРСПС (Arthur, 1962)].
дНЙЮГЮМЮ РПЮМЯТЮГНБЮЪ Х РПЮМЯНБЮПХЮКЭМЮЪ ОЕПЕДЮВЮ ПХЙЙЕРЯХИ С ЙКЕЫЕИ D. andersoni, D. variabilis1 H. Ieporis polustr is, A. CajennenseH A. striatum бНЯОПХХЛ ВХБШЛХ УНГЪЕБЮЛХ Б ПЮИНМЕ яЙЮКХЯРШУ ЦНП ЪБКЪЧРЯЪ ЦКЮБМШЛ НАПЮГНЛ ЛНКНДШЕ ЦПШ╜ГСМШ ≈ ЙПНКХЙХ ХКХ ГЮИЖШ. еЯРЕЯРБЕММЮЪ ГЮПЮФЕММНЯРЭ ПХЙЙЕРЯХЪЛХ СЯРЮМНБКЕМЮ С Microtus pennsilvaπicus (дСЦКЮЯ (Douglas, 1954)). Silvilagus IJoridanus [ьХПХ Х ДП. (Shiri11961)], Lepus americanus citellus lateralis. Entamias amoenus (аЧПЦДНПТЕП Х ДП. (Burgdorfer1 1962)J.
хЙЯНДНБШЕ ЙКЕЫХ ХЛЕЧР БЮФМНЕ ГМЮВЕМХЕ Б ОНДДЕПФЮМХХ ОПХПНДМШУ НВЮЦНБ йС-КХУНПЮДЙХ. сЯРЮМНБКЕМН ЛХПНБНЕ ПЮЯОПНЯРПЮМЕМХЕ ЩРНЦН ГЮАНКЕ БЮМХЪ. бОЕПБШЕ НМН АШКН НАМЮПСФЕМН дЕППХЙНЛ (Derrick) Б 1933 Ц. Б аПХЯ- АНМЕ (юБЯРПЮКХЪ) ЯПЕДХ ЯКСФЮЫХУ ЙПСОМШУ АНЕМ. бНГАСДХРЕКЭ АШК БШДЕ╜КЕМ ХГ ЙПНБХ Х ЛНВХ АНКЭМШУ ЩРХЛ ФЕ ЮБРНПНЛ Б 1937 Ц. б РНЛ ФЕ ЦНДС аЕП МЕР Х тПХЛЮМ (Burnet, Freeman) НОХЯЮК БНГАСДХРЕКЪ ЙЮЙ ПХЙЙЕРЯХЕОНДНАМШИ НПЦЮМХГЛ, Ю дЕППХЙ ДЮК МЮГБЮМХЕ Rickettsia bur∏eti. б юЛЕПХЙЕ ХГСВЕМХЕ ПХЙЙЕРЯХИ аЕПМЕРЮ МЮВЮРН Я 1935 Ц., ЙНЦДЮ ЩРНР БНГАСДХРЕКЭ АШК БШДЕКЕМ ХГ ЙКЕЫЕИ D. andersoni [дЩБХЯ Х йНЙЯ (Davis Х яНУ, 1938)1.
тХКХО (Philip) Б 1948 Ц. ОПЕДКНФХК ДКЪ R. burneti МНБНЕ ПНДНБНЕ МЮГБЮ ,МХЕ ≈Coxiella burneti. ОНЯЙНКЭЙС НМХ ХЛЕЧР МЕЙНРНПШЕ НРКХВХЪ НР ДПСЦХУ
ПХЙЙЕРЯХИ. б КХРЕПЮРСПЕ ДН МЮЯРНЪЫЕЦН БПЕЛЕМХ СОНРПЕАКЪЧРЯЪ НАЮ ЩРХ МЮХЛЕМНБЮМХЪ.
б юБЯРПЮКХХ ЯОНМРЮММЮЪ ГЮПЮФЕММНЯРЭ ЙКЕЫЕИ БНГАСДХРЕКЕЛ КХУНПЮД╜ЙХ Ky БОЕПБШЕ АШКЮ СЯРЮМНБКЕМЮ С ЙКЕЫЕИ Haemaphysalis IiunierosaLeach. ЯНАПЮММШУ Я ЦПШГСМНБ ЛЕЯРМНИ ТЮСМШ ≈ Bandicoot torosus (дЕППХЙ Х ДП., 1939, 1940).
C ЛНЛЕМРЮ НАМЮПСФЕМХЪ ОЕПБШУ ЙКЕЫЕИ D. andersoni,a ГЮРЕЛ H. hume- rosa, ГЮПЮФЕММШУ ПХЙЙЕРЯХЕИ аЕПМЕРЮ, ДН МЮЯРНЪЫХУ ДМЕИ СЯРЮМНБКЕМН 30 БХДНБ ХЙЯНДНБШУ ЙКЕЫЕИ ≈ ЕЯРЕЯРБЕММШУ МНЯХРЕКЕИ R. burneti Б ПЮГ╜КХВМШУ ЯРПЮМЮУ ЛХПЮ [аЕПЦ (Berg1 1953), бЕИЕП (Weyer, 1953)]. б яНБЕР╜ЯЙНЛ яНЧГЕ ЯОНМРЮММНЕ МНЯХРЕКЭЯРБН ЙКЕЫЕИ R. burneti БШЪБКЕМН БОЕПБШЕ Б 1953 Ц. ОНВРХ НДМНБПЕЛЕММН Я ДХЮЦМНЯРХПНБЮМХЕЛ ГЮАНКЕБЮМХЪ КЧДЕИ КХУН╜ПЮДЙНИ йС. йКЕЫХ, ГЮПЮФЕММШЕ ПХЙЙЕРЯХЪЛХ аЕПМЕРЮ, НАМЮПСФЕМШ Б ПЕЯ╜ОСАКХЙЮУ яПЕДМЕИ юГХХ: рСПЙЛЕМЯЙНИ, рЮДФХЙЯЙНИ, сГАЕЙЯЙНИ, йХПЦХГ╜ЯЙНИ яяп, Ю РЮЙФЕ Б йЮГЮУЯЙНИ, сЙПЮХМЯЙНИ, юГЕПАЮИДФЮМЯЙНИ, цПСГХМ╜ЯЙНИ яяп, дЮЦЕЯРЮМЯЙНИ ACCP Х Б пятяп.
й МЮЯРНЪЫЕЛС БПЕЛЕМХ Б яяяп ХГБЕЯРМН 20 БХДНБ ХЙЯНДНБШУ ЙКЕЫЕИ ЕЯРЕЯРБЕММШУ МНЯХРЕКЕИ R. burneti.
яОНЯНАМНЯРЭ БНЯОПХМХЛЮРЭ ПХЙЙЕРЯХХ аЕПМЕРЮ Х ОЕПЕДЮБЮРЭ ХУ Б ОПН╜ЖЕЯЯЕ ЙПНБНЯНЯЮМХЪ ОНДНОШРМШЛ ЛНПЯЙХЛ ЯБХМЙЮЛ СЯРЮМНБКЕМЮ ДКЪ ЯКЕДСЧ╜ЫХУ БХДНБ: Hyaloinma asiaticum, H. anatolicum anatolicum, H. plumbeum turaπicum, Rhipicephalus turanicus (3. л. фЛЮЕБЮ, ю. ю. оВЕКЙХМЮ Х ДП., 1955), H. plumbeum plumbeum (х. б. рЮПЮЯЕБХВ, 1956), (Rhipicephalus bursa (х. б. рЮПЮЯЕБХВ, 1956; ю. ю. лЮПЙНБ, л. о. вСЛЮЙНБ, ю. о. гНРНБ, м. х. яРЕОЮМНБЮ, 1957), Rh. sanguineus (р. к. оПНПЕЬМЮЪ, ю. б. хБЮМНБ. 1957; ю. х. ъЙНБКЕБ, ю. х. оНКНГНБ, 1962), Rh. turanicus (т. м. пХГЮЕБЮ, I960), I. ricinus, H. asiaticum (ю. а. дЮИРЕП, 1963). щРХ БХДШ ЙКЕЫЕИ ЯНУПЮМЪЧР ПХЙЙЕРЯХХ ОН УНДС ЛЕРЮЛНПТНГЮ Х РПЮМЯНБЮПХЮКЭМН. оНД╜ПНАМШЕ ХЯЯКЕДНБЮМХЪ Н КНЙЮКХГЮЖХХ ПХЙЙЕРЯХИ аЕПМЕРЮ Б НПЦЮМХГЛЕ ЙКЕ╜ЫЕИ ОПНБЕДЕМШ IO. я. аЮКЮЬНБШЛ Х ю. а. дЮИРЕПНЛ (1963). оН ДЮММШЛ ЩРХУ ЮБРНПНБ, ЦКЮБМШЛ ЛЕЯРНЛ КНЙЮКХГЮЖХХ ПХЙЙЕРЯХИ аЕПМЕРЮ Б ЙКЕЫЮУ H. asia╜ticum ЯКСФЮР ОХЫЕБЮПХРЕКЭМШЕ ЙКЕРЙХ ЯПЕДМЕИ ЙХЬЙХ. оПХ ПЮГПСЬЕМХХ ЩРХУ ЙКЕРНЙ БНГАСДХРЕКЭ ОНОЮДЮЕР Б ОПНЯБЕР ЙХЬЕВМХЙЮ, Ю ГЮРЕЛ БЛЕЯРЕ Я ТЕ╜ЙЮКХЪЛХ Х ОНКСОЕПЕБЮПЕММНИ ЙПНБЭЧ БШБНДХРЯЪ МЮПСФС.
пХЙЙЕРЯХХ БШБНДЪРЯЪ Б АНКЭЬНЛ ЙНКХВЕЯРБЕ БН БМЕЬМЧЧ ЯПЕДС БН БПЕЛЪ ОХРЮМХЪ Х ОНЯКЕКХМНВМНЦН ДНПЮГБХРХЪ. б ЯКЧММШУ ФЕКЕГЮУ, ОН ХУ ДЮММШЛ, Б ЙКЕЫЮУ H. asiaticum ПХЙЙЕРЯХХ НРЯСРЯРБСЧР. оПХЯСРЯРБХЕ ПХЙЙЕРЯХИ МЕ МЮПСЬЮКН С ЙКЕЫЕИ ОПНЖЕЯЯШ БМСРПХЙКЕРНВМНЦН ОХЫЕБЮПЕМХЪ. гЮПЮФЕМ╜МНЯРЭ ПХЙЙЕРЯХЪЛХ аЕПМЕРЮ МЕ БКХЪКЮ МХ МЮ ПЮГБХРХЕ НРДЕКЭМШУ ТЮГ ЙКЕ╜ЫЕИ, МХ МЮ ЯНГПЕБЮМХЕ ОНКНБШУ ОПНДСЙРНБ.
щРХ ДЮММШЕ ЕЫЕ ПЮГ ОНЙЮГШБЮЧР, ВРН НПЦЮМХГЛ ЙКЕЫЮ ЪБКЪЕРЯЪ ДКЪ ПХЙ╜ЙЕРЯХИ аЕПМЕРЮ АКЮЦНОПХЪРМНИ ЯПЕДНИ НАХРЮМХЪ.
нРЯСРЯРБХЕ РПЮМЯНБЮПХЮКЭМНИ ОЕПЕДЮВХ ПХЙЙЕРЯХИ аЕПМЕРЮ С ВЮЯРХ ХМТХЖХПНБЮММШУ ЯЮЛНЙ ЮБРНПШ НАЗЪЯМЪЧР МЮКХВХЕЛ ГЮЫХРМШУ АЮПЭЕПНБ, ОПЕОЪРЯРБСЧЫХУ ОПНМХЙМНБЕМХЧ ПХЙЙЕРЯХИ Б ПЮГБХБЮЧЫХЕЯЪ ЪИЖЕЙКЕРЙХ. мЮ ПЮММХУ ЯРЮДХЪУ НБНЦЕМЕГЮ, ЙНЦДЮ ННЖХРШ НРДЕКЕМШ НР ЦЕЛНКХЛТШ КХЬЭ РНМЙНИ НАНКНВЙНИ ≈ tunica propria ≈ МЕР ОПЕОЪРЯРБХИ ДКЪ ОПНМХЙМНБЕМХЪ Б МХУ ПХЙЙЕРЯХИ, РНЦДЮ ЙЮЙ Б ННЖХРЮУ IV ЯРЮДХХ ЯТНПЛХПНБЮММЮЪ МЮПСФМЮЪ ЙСРХЙСКЪПМЮЪ НАНКНВЙЮ УНПХНМЮ ОПЕОЪРЯРБСЕР ХУ ХМТХЖХПНБЮМХЧ. нАЪГЮ╜РЕКЭМШЛ СЯКНБХЕЛ ОЕПЕДЮВХ ПХЙЙЕРЯХИ аЕПМЕРЮ ОНРНЛЯРБС ДНКФМЮ АШРЭ ДНЯРЮРНВМН БШЯНЙЮЪ ЙНМЖЕМРПЮЖХЪ БНГАСДХРЕКЪ Б ЦЕЛНКХЛТЕ, ГЮПЮФЕМХЕ ННЖХРНБ ДН НЙНМВЮМХЪ ТНПЛХПНБЮМХЪ УНПХНМЮ С ЙКЕЫЕИ, ЯНЯРНЪМХЕ БНГАС╜ДХРЕКЪ Х ПЪД ДПСЦХУ ТЮЙРНПНБ.
рЮАКХЖЮ 16
йКЕЬХ-МНЯХРЕКХ ПХЙЙЕРЯХИ аЕПМЕРЮ Б НВЮЦЮУ йС-КХУНПЮДЙХ Б ОСЯРШММШУ, ОПЕДЦНПМН-ЯРЕОМШУ, ЯРЕОМШУ Х КЕЯМШУ КЮМДЬЮТРЮУ яяяп
| бХД | лЕЯРН НАМЮПСФЕМХЪ | юБРНПШ |
| Hyalomma anatolicum | йХПЦХГЯЙЮЪ яяп, дФЮКЮК- юАЮДЯЙЮЪ НАК. | вСЛЮЙНБ, 1954 |
| H. anatolicum anato- | йХПЦХГЯЙЮЪ яяп, юПЮБЮМЯЙХИ | фЛЮЕБЮ, лХЫЕМЙН, оВЕКЙХМЮ. |
| Iicum | ПЮИНМ | 1956 |
| рН ФЕ | рЮДФХЙЯЙЮЪ CCP | нЯРПНБЯЙЮЪ, ъЯХМЯЙХИ, уЮЬХ╜ЛНБ, 1955 фЛЮЕБЮ, оВЕКЙХМЮ, йЮПСКХМ. гСАЙНБЮ, лХЫЕМЙН, 1954. 1955 |
| H. anatolicum ЕУЯЮ- | рУ'ПЙЛЕМЯЙЮЪ яяп. ДНКХМЮ | |
| vatum | юЛС-дЮПЭХ | |
| рН ФЕ | сЙПЮХМЯЙЮЪ CCP | оЮЙЕЖХМ Х йНЯРЕЖЙХМ, 1957 |
| ╩ > | йХПЦХГЯЙЮЪ CCP | оПНПЕЬМЮЪ. 1959 фЛЮЕБЮ, оВЕКЙХМЮ, йЮПСКХМ. гСАЙНБЮ, лХЫЕМЙН, 1954, 1955 |
| H. detritum | сГАЕЙЯЙЮЪ CCP | |
| рН ФЕ | сГАЕЙЯЙЮЪ CCP | кШЯСМЙХМЮ, гБЮЦЕКЭЯЙЮЪ, 1957 нЯРПНБЯЙЮЪ. ъЯХМЯЙХИ, уЮЬХ╜ЛНБ, 1955 кШЯСМЙХМЮ, гБЮЦЕКЭЯЙЮЪ, 1957 |
| ╩ > | рЮДФХЙЯЙЮЪ CCP | |
| H. plumbeum turani- | сГАЕЙЯЙЮЪ яяп, рЮЬЙЕМР | |
| cum | ||
| рН ФЕ | йХПЦХГЯЙЮЪ CCP | оПНПЕЬМЮЪ, 1957. оПНПЕЬМЮЪ, еБДНЬЕМЙН, 1959, хЛЮМНБ. 1962 |
| H. scupense | йЮГЮУЯЙЮЪ яяп, юКЛЮ-юРХМ╜ЯЙЮЪ НАК. | яСУНДНЕБЮ, 1961 |
| рН ФЕ | рН ФЕ | юЛЮМФСКНБ, юЛНЯЕМЙНБЮ, оН- ЯРПХВЕБЮ, 1963 |
| H. plumbeum plum- | сЙПЮХМЯЙЮЪ яяп. йПШЛ | рЮПЮЯЕБХВ, 1956 |
| beum | ||
| рН ФЕ | рЮДФХЙЯЙЮЪ CCP | нЯРПНБЯЙЮЪ, ъЯХМЯЙХИ, уЮЬХ╜ЛНБ, 1955 яРЮПЙНБЮ Х лХПГЮЕБЮ, 1956 |
| >> | юГЕПАЮИДФЮМЯЙЮЪ яяп, юДФМ- ЙЮЛАСКЭЯЙХИ ПЮИНМ | |
| Dermacentor daghes- Ianicus | рЮДФХЙЯЮЪ CCP | нЯРПНБЯЙЮЪ, ъЯХМЯЙХИ, яСПЙН╜БЮ, 1959 юПУЮМЦЕКЭЯЙХИ, юУХЛАЮЕБ, 1960 |
| рН ФЕ | йЮГЮУЯЙЮЪ яяп. юКЛЮ-юРХМ╜ЯЙЮЪ НАК. | |
| ╩ ╩ | рН ФЕ | яСУНДНЕБЮ, 1961 |
| ╩ > | ╩ > | юЛЮМФСКНБ, юЛНЯЕМЙНБЮ, оН- ЯРПХВЕБЮ, 1963 |
| D. povlovskyi | йХПЦХГЯЙЮЪ CCP | хЛЮМНБ, 1962 |
| D. Tnarginatus | йЮГЮУЯЙЮЪ яяп, юЙЛНКХМЯЙЮЪ НАК. | фЛЮЕБЮ, йЮПСКХМ, оВЕКЙХМЮ. ьЕУЮМНБ, 1956 |
| рН ФЕ | сЙПЮХМЯЙЮЪ яяп, дМЕОПНОЕР╜ПНБЯЙЮЪ НАК. | йЕЯЮПЕБ, 1957 |
| ╩ > | йХПЦХГЯЙЮЪ CCP | оПНПЕЬМЮЪ, пЮОНОНПР, еБДН ЬЕМЙН, йХВЮРНБ, 1960 йНЖХМЪМ, 1960 |
| ╩ ╩ | юПЛЪМЯЙЮЪ CCP | |
| Rhipicephalus bursa | сЙПЮХМЯЙЮЪ CCP1йПШЛ | рЮПЮЯЕБХВ, 1955 оЮЬЙХМ Х йНЯРЕЖЙХИ, 1957 |
| рН ФЕ | рН ФЕ | |
| ╩ ╩ | юГЕПАЮИДФЮМЯЙЮЪ CCP | яРЕПУНБЮ. лХПГЮЕБЮ, 1956 |
| Rh. turanicus | сГАЕЙЯЙЮЪ CCP | кШЯСМЙХМЮ, 1960 |
| рН ФЕ | йХПЦХГЯЙЮЪ CCP | хБЮМНБ, 1962 |
| Rhipicephalus rossi- | дЮЦЕЯРЮМЯЙЮЪ ACCP | аЮЯНБЮ Х ДП., 1960 |
| CUS Rh. sanguineus | сГАЕЙЯЙЮЪ CCP | кШЯСМЙХМЮ, гБЮЦЕКЭЯЙЮЪ, 1956 |
| рН ФЕ | рЮДФХЙЯЙЮЪ CCP | нЯРПНБЯЙЮЪ, ъЯХМЯЙХИ, уЮЬХ╜ЛНБ, 1955 |
| ╩ > | йХПЦХГЯЙЮЪ яяп, хЯЯШЙ-йСКЭ╜ЯЙЮЪ НАК. | оПНПЕЬМЮЪ, хБЮМНБ, 1957 |
оПНДНКФЕМХЕ РЮАК. 16
| бХД | УвЕЯРН НАМЮПСФЕМХЪ | юБРНПШ |
| Rh. sanguineus | йХПЦХГЯЙЮЪ яяп, хЯЯШЙ-йСКЭ╜ЯЙЮЪ НАК. | оПНПЕЬМЮЪ, лХЫЕМЙН, 1958 |
| рН ФЕ | юГЕПАЮИДФЮМЯЙЮЪ CCP дЮЦЕЯРЮМЯЙЮЪACCP | яРЕПУНБЮ, лХПГЮЕБЮ. 1956 тЕДНПНБЮ, рЮПЮЯЕБХВ, яЕПЦЕЕБЮ, ьКЪУРСПНБЮ, оНОНБЮ, 1957 |
| Boophilus Calcaratus | ||
| Ixodes ricinus | цПСГХМЯЙЮЪ CCP | аЮАЮМНБЮ, 1959 |
| рН ФЕ | пятяп, кЕМХМЦПЮДЯЙЮЪ НАК. | дЮИРЕП, юЛНЯЕМЙНБЮ, йКЕМНБЮ, 1961 оВЕКЙХМЮ, фЛЮЕБЮ. гСАЙНБЮ, 1956 |
| I. Crenulatus | йЮГЮУЯЙЮЪ яяп, йСЯРЮМЮИЯЙЮЪ НАК. | |
| I. Iividus (-plumbeus) | йЮГЮУЯЙЮЪ яяп, йСЯРЮМЮИ╜ЯЙЮЪ НАК., юЙЛНКХМЯЙЮЪ НАК. | лЮУЛЕРНБ, 1961 фЛЮЕБЮ, оВЕКЙХМЮ, 1957 |
| I Irontalis | дЮЦЕЯРЮМЯЙЮЪ ACCP | аЮЯНБЮ. вЕПМХЙНБ, яСВЙНБ, пСДМЕБ, 1960 |
бНГЛНФМН ОЕПЕФХБЮМХЕ ПХЙЙЕРЯХИ аЕПМЕРЮ Б ГХЛСЧЫХУ ХМТХЖХПНБЮМ╜МШУ ЦНКНДМШУ МХЛТЮУ I. riciπus Х Б ЯШРШУ КХВХМЙЮУ. сЯРЮМНБКЕМН РЮЙФЕ ЯНУПЮМЕМХЕ БНГАСДХРЕКЪ Б ОЕПЕКХМЪБЬХУ Б ОПХПНДЕ МХЛТЮУ. оПХ ЩРНЛ ОНЙЮ╜ГЮМН, ВРН ДКХРЕКЭМНЕ ОПЕАШБЮМХЕ ПХЙЙЕРЯХИ аЕПМЕРЮ Б ГХЛСЧЫХУ ЙКЕЫЮУ МЕ НЯКЮАКЪЕР БХПСКЕМРМШУ ЯБНИЯРБ БНГАСДХРЕКЪ (ю. а. дЮИРЕП, 1963).
мЮ РЕЯМСЧ ЯБЪГЭ ПХЙЙЕРЯХХ аЕПМЕРЮ Х ХЙЯНДНБШУ ЙКЕЫЕИ СЙЮГШБЮЧР ХЯЯКЕДНБЮМХЪ Н ДКХРЕКЭМНЛ ОЕПЕФХБЮМХХ БНГАСДХРЕКЪ Б ЛЕПРБШУ ГЮПЮФЕМ╜МШУ ЙКЕЫЮУ. нАМЮПСФЕМШ ПХЙЙЕРЯХХ Б ОНЦХАЬХУ H. asiaticum, Rh. tura- πicus, УПЮМХБЬХУЯЪ Б РЕВЕМХЕ 8 ≈ 10 КЕР Б ЙНЛМЮРМШУ СЯКНБХЪУ Х Б КЕРМЕЛ ЯЮПЮЕ (о. ю. оЕРПХЫЕБЮ, 3. л. фЛЮЕБЮ, ю. ю. оВЕКЙХМЮ, 1963).
аНКЭЬЮЪ ЙНМЖЕМРПЮЖХЪ БНГАСДХРЕКЪ МЮАКЧДЮЕРЯЪ Б ХЯОПЮФМЕМХЪУ ХЙЯН╜ДНБШУ ЙКЕЫЕИ. щЙЯЙПЕЛЕМРШ ОНКНБНГПЕКШУ I. ricinus ЯНДЕПФЮКХ ПХЙЙЕРЯХИ ОПХ ПЮГБЕДЕМХХ 10 5(к. а. дЮИРЕП, 1963). хЯОПЮФЕММЪ D. pictus ЯНДЕПФЮКХ ПХЙЙЕРЯХИ Б РХРПЕ 10 3(пЕМ (Rehn, 1958)1, D. andersoni ≈ Б РХРПЕ 2 ? ? 10 12(тХК ХО, 1948). хЯОПЮФЕМХЪ ХЙЯНДНБШУ ЙКЕЫЕИ ДКХРЕКЭМНЕ БПЕЛЪ, Б РЕВЕМХЕ МЕЯЙНКЭЙХУ ЛЕЯЪЖЕБ, НЯРЮЧРЯЪ ХМТЕЙЖХНГМШЛХ ДКЪ ЛНПЯЙХУ ЯБХ╜МНЙ: С D. andersoni ДН 586 ДМЕИ (тХКХО, 1948), С Rh. Iuranicus НР 180 ДН 225 ДМЕИ, ОПХ РЕЛОЕПЮРСПЕ + 4, 4- 35╟ (т. м. пХГЮЕБЮ).
пНКЭ ХЙЯНДНБШУ ЙКЕЫЕИ Б НВЮЦЮУ йС-КХУНПЮДЙХ ЯБНДХРЯЪ Й ЯНУПЮМЕМХЧ БНГАСДХРЕКЪ Б ОПХПНДЕ Х Й ПЮЯОПНЯРПЮМЕМХЧ ЕЦН ЯПЕДХ ДХЙХУ Х ДНЛЮЬМХУ ФХБНРМШУ. бНГАСДХРЕКЭ ДКХРЕКЭМНЕ БПЕЛЪ ЯНУПЮМЪЕРЯЪ Б ЙКЕЫЮУ, НДМЮЙН ОЕПЕДЮВЮ ЕЦН ВЕКНБЕЙС БН БПЕЛЪ ЙПНБНЯНЯЮМХЪ МЮАКЧДЮЕРЯЪ НВЕМЭ ПЕДЙН. хЯРНВМХЙНЛ ГЮПЮФЕМХЪ ВЕКНБЕЙЮ ОПХ КХУНПЮДЙЕ Ky ЯКСФХР ЛНКНЙН ЙНПНБ, ОШКЭ, ЯНДЕПФЮЫЮЪ ПХЙЙЕРЯХХ, ЩЙЯЙПЕЛЕМРШ ЙКЕЫЕИ.
щЙНКНЦХВЕЯЙХЕ НЯНАЕММНЯРХ ХЙЯНДНБШУ ЙКЕЫЕИ ЯОНЯНАЯРБСЧР ЬХПНЙНЛС ПЮЯОПНЯРПЮМЕМХЧ Х ДКХРЕКЭМНЛС ЯНУПЮМЕМХЧ ПХЙЙЕРЯХИ аЕПМЕРЮ Б ОПХПНДЕ. мЮХАНКЕЕ НРВЕРКХБН ЩРН ОПНЪБКЪЕРЯЪ Б ОПХПНДМШУ НВЮЦЮУ ОСЯРШМЭ. C ОН╜ЛНЫЭЧ Hyalomma, Rhipicephalus ПХЙЙЕРЯХХ ХЛЕЧР БНГЛНФМНЯРЭ ПЮЯЯЕ╜КЪРЭЯЪ МЮ АНКЭЬХЕ ПЮЯЯРНЪМХЪ, Ю ЙКЕЫХ Ornithodorus ЯОНЯНАЯРБСЧР БНГАС╜ДХРЕКЧ СДЕПФХБЮРЭЯЪ МЮ РЕППХРНПХХ ДКХРЕКЭМНЕ БПЕЛЪ ≈ ЛМНЦХЕ ЦНДШ. бНГ╜АСДХРЕКЭ Б ОСЯРШММНИ ГНМЕ ЛНФЕР НЯРЮБЮРЭЯЪ ФХГМЕДЕЪРЕКЭМШЛ ЙПСЦКШИ ЦНД АКЮЦНДЮПЪ ЙПСЦКНЦНДХВМНИ ЮЙРХБМНЯРХ ХЙЯНДНБШУ ЙКЕЫЕИ. б РЕВЕМХЕ БЯЕЦН ЦНДЮ ХЛЕЧРЯЪ ЮЙРХБМШЕ ЙКЕЫХ, Р. Е. ЮЙРХБМНЯРЭ НДМХУ БХДНБ ЯЛЕМЪЕРЯЪ ДПС-
ЦХЛМ. б ЯРЕОМШУ КЮМДЬЮТРЮУ СЯКНБХИ ДКЪ ДКХРЕКЭМНЦН ОЕПЕФХБЮМХЪ ЛЕМЭЬЕ бХДНБНИ ЯНЯРЮБ ЙКЕЬЕИ ГДЕЯЭ ЛЕМЕЕ ПЮГМННАПЮГЕМ Х ЯЕГНМ ЮЙРХБМНЯРХ ЙНПНВЕ
бХПСЯМШЕ ХМТЕЙЖХХ. щОХДЕЛХНКНЦХВЕЯЙНЕ ГМЮВЕМХЕ ХЙЯНДНБШУ ЙКЕЫЕИ ОПХ ХМТЕЙЖХЪУ БХПСЯМНИ ЩРХНКНЦХХ Х, Б ВЮЯРМНЯРХ, ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ АШКН СЯРЮМНБКЕМ БОЕПБШЕ МЮ дЮКЭМЕЛ бНЯРНЙЕ.
хГ ЙКЕЫЕИ Ixodes persulcatus Х Haemophysalis concinna АШКХ БШДЕКЕМШ ЬРЮЛЛШ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ (м. б. пШФНБ Х ю. II. яЙПШММХЙ, 1937. 1938).
хГСВЕМХЕ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ МЮВЮРН Б 1937 Ц. ЙНЛОКЕЙЯМНИ ЩЙЯОЕ╜ДХЖХЕИ мЮПЙНЛГДПЮБЮ (XCP ОНД ПСЙНБНДЯРБНЛ ОПНТ. к. ю. гХКЭАЕПЮ. б 1938 Ц. АШКН ОПНДНКФЕМН ХГСВЕМХЕ РЕУ ФЕ НВЮЦНБ ЙНЛОКЕЙЯМНИ ЩЙЯОЕДХЖХЕИ ОНД ПСЙН╜БНДЯРБНЛ ЮЙЮД. е. м. оЮБКНБЯЙНЦН Х ОПНТ. ю. ю. яЛНПНДХМЖЕБЮ.
нАМЮПСФЕМХЕ ЯОНМРЮММН ГЮПЮФЕММШУ ЙКЕЫЕИ, Ю РЮЙФЕ ДХЙХУ ОНГБНМНВ╜МШУ ФХБНРМШУ-ОПНЙНПЛХРЕКЕИ ЙКЕЫЕИ ДЮКН БНГЛНФМНЯРЭ е. II. оЮБКНБ╜ЯЙНЛС НУЮПЮЙРЕПХГНБЮРЭ ЙКЕЫЕБНИ ЩМЖЕТЮКХР ЙЮЙ ГЮАНКЕБЮМХЕ Я ОПХПНДМНИ НВЮЦНБНЯРЭЧ.
б ОНЯКЕДСЧЫХЕ ЦНДШ ОПХПНДМШЕ НВЮЦХ ДЮММНЦН ГЮАНКЕБЮМХЪ АШКХ НАМЮ╜ПСФЕМШ МЮ АНКЭЬНИ РЕППХРНПХХ КЕЯМНИ ГНМШ яНБЕРЯЙНЦН яНЧГЮ. оЕПЕМНЯВХ╜ЙЮЛХ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ, ОНЛХЛН I. persulcatus, СЯРЮМНБКЕМШ Х ДПСЦХЕ БХДШ ХЙЯНДНБШУ ЙКЕЫЕИ. б аЕКНПСЯЯХХ НВЮЦХ ГЮАНКЕБЮМХЪ Я ОЕПЕМНЯВХЙНЛ I. riciπus ПЮЯОПНЯРПЮМЕМШ БН БЯЕУ ПЮИНМЮУ ПЕЯОСАКХЙХ, МН Б НЯМНБМНЛ НМХ ОПХСПНВЕМШ Й ЖЕМРПЮКЭМНЛС ПЮИНМС ЕКНБН-ЬХПНЙНКХЯРБЕММШУ КЕЯНБ. б сЙ╜ПЮХМЯЙНИ CCP НЯМНБМШЛ ОЕПЕМНЯВХЙНЛ БНГАСДХРЕКЪ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ ЪБКЪЕРЯЪ РЮЙФЕ I. ricinus, ОПХСПНВЕММШИ ГДЕЯЭ Й УБНИМН-КХЯРБЕММШЛ Х КХЯР╜БЕММШЛ (ДСАНБШЛ, АСЙНБШЛ) КЕЯЮЛ, Я ЙСЯРЮПМХЙНБШЛ ОНДКЕЯЙНЛ (е. л. еЛ- ВСЙ, 1962). щРНР БХД ЙКЕЫЮ ЪБКЪЕРЯЪ ОЕПЕМНЯВХЙНЛ Х Б гЮЙЮПОЮРЭЕ, НАХРЮЧ╜ЫХИ ГДЕЯЭ МЮ ОЮЯРАХЫЮУ ЯЕКЭЯЙНУНГЪИЯРБЕММШУ ФХБНРМШУ (к. я. яЕЦЮКЭ.
б. л. лЕЫЕМЙН, 1963). дНЙЮГЮМЮ ЕЯРЕЯРБЕММЮЪ ГЮПЮФЕММНЯРЭ D. pictus (б. к. бЮЯХКЭЕБЮ, 1957), ПЮЯОПНЯРПЮМЕММНЦН Б КСЦНБШУ НВЮЦЮУ оНКЕЯЭЪ.
йКЕЫ H. concinna ЪБКЪЕРЯЪ ОЕПЕМНЯВХЙНЛ ЩРНИ ХМТЕЙЖХХ Б оПХЛНПЯЙНЛ ЙПЮЕ (ю. м. яЙПШММХЙ, м. б. пШФНБ, 1941), Б уЮАЮПНБЯЙНЛ ЙПЮЕ (ю. ю. бЕ╜ПЕРЮ, л. м. яСЬЙХМЮ, б. о. дЕДЧЙХМЮ, 1958), Б йПЮЯМНЪПЯЙНЛ ЙПЮЕ (м. б. бНЫЮЙХМЮ, л. я. дЮБШДНБЮ, 1954). б ОЕПБШУ ДБСУ ПЮИНМЮУ ГЮПЮФЕММШЕ ЙКЕЫХ ПЮЯОПНЯРПЮМЕМШ Б ЙСЯРЮПМХЙНБШУ ГЮПНЯКЪУ Х ЙНВЙНБЮРШУ АНКНРЮУ, Б йПЮЯМНЪПЯЙНЛ ЙПЮЕ НМХ БЯРПЕВЮЧРЯЪ Б АЕПЕГНБН-НЯХМНБШУ ЙНКЙЮУ КЕЯН╜ЯРЕОМНИ ГНМШ. б ОПХЛНПЯЙНЛ ЙПЮЕ БШЪБКЕМЮ ЯОНМРЮММЮЪ ГЮПЮФЕММНЯРЭ ЙКЕ╜ЫЕИ D. Silvarum (ю. м. яЙПШММХЙ, м. б. пШФНБ, 1941) Х Haemaphysalis japonica doughlasi (ю. о. аЕКХЙНБЮ Х к. ц. рЮРЮПХМНБЮ, 1960).
б КЕЯНЯРЕОМНИ ГНМЕ ОЕПЕМНЯВХЙЮЛХ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ СЯРЮМНБКЕМШ D. margiπatus Б йСЯРЮМЮИЯЙНИ НАКЮЯРХ йЮГЮУЯРЮМЮ (х. л. цНПЬЙНБЮ, л. я. оЮБКНБЮ, л. й- рЧЬМЪЙНБЮ, б. л. оНОНБ Я ЯНЮБРНПЮЛХ, 1959) Х Б йСИАШЬЕБЯЙНИ НАКЮЯРХ (к. а. оНОНБЮ, 1963).
б гЮОЮДМНИ яХАХПХ БШЪБКЕМЮ ЯОНМРЮММЮЪ ГЮПЮФЕММНЯРЭ БНГАСДХРЕКЕЛ ЩРНЦН ГЮАНКЕБЮМХЪ ЙКЕЫЕИ I. plumbeus (ч. б. тЕДНПНБ, л. й. рЧЬМЪЙНБЮ 1958).
мЕЙНРНПШЕ БХДШ ЙКЕЫЕИ ЪБКЪЧРЯЪ ЯРНИЙХЛХ УПЮМХРЕКЪЛХ БНГАСДХРЕКЪ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ. яНУПЮМЕМХЕ БХПСЯЮ ОН УНДС ЛЕРЮЛНПТНГЮ ХГСВЕМН С I. persulcatus (ю. й- ьСАКЮДГЕ, ц. б. яЕПДЧЙНБЮ, 1939; л. о. вСЛЮЙНБ, 1939; я. ц. цКЮДЙХУ, 1939; л. о. вСЛЮЙНБ, 1944; ю. к. дСЛХМЮ, 1958), D. sil- varum (ю. м. яЙПШММХЙ Х м. б. пШФНБ, 1941), H. concinna (ю. б. йНГКНБЮ Х б. д. яНКНБЭЕБ, 1941), I. ricinus !аЕМДЮ (Benda, 1958)]. оЕПЕДЮВЮ БХПСЯЮ ЯЮЛЙНИ ЯБНЕЛС ОНРНЛЯРБС СЯРЮМНБКЕМЮ С 1. persulcatus (ю. й. ьСАКЮДГЕ, ц. б. яЕПДЧЙНБЮ. 1939; е. м. оЮБКНБЯЙХИ Х б- д. яНКНБЭЕБ, 1941; л. о.
вСЛЮЙНБ, 1944); 1. ricinus (цПЕВХЙНБЮ Х аЕМДЮ, 1958; пФХЦЮВЕЙ, 1962).
D. Silvarum (ю. м. яЙПШММХЙ, м. б. пШФНБ. 1944) Х H. concinna (ю. б. йНГ╜КНБЮ Х б. д. яНКНБЭЕБ, 1941).
бХПСЯ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ ОЕПЕГХЛНБШБЮЕР Б ЙКЕЬЮУ I. persulcatus (е. м. кЕБЙНБХВ, ю. м. яЙПШММХЙ, 1940), Б 1. ricinus (пФХЦЮВЕЙ, 1960).
пЮЯОПЕДЕКЕМХЕ БХПСЯЮ Б НПЦЮМХГЛЕ 1. persulcatus, х. concinna ХГСВЕМН
E. м. оЮБКНБЯЙХЛ Х б. д. яНКНБЭЕБШЛ (1940). нМХ ОНЙЮГЮКХ, ВРН БХПСЯ МЮУН╜ДХРЯЪ БН БЯЕУ НПЦЮМЮУ ГЮПЮФЕММНЦН ЙКЕЫЮ. юЙРХБМШИ БХПСЯ БШЪБКЕМ ХЛХ Б ЯКЧММШУ ФЕКЕГЮУ, ЪХВМХЙЕ.'
оПХ ЩЙЯОЕПХЛЕМРЮКЭМНЛ ГЮПЮФЕМХХ I. persulcatus ЙНПЛКЕМХЕЛ МЮ АЕКШУ .ЛШЬЮУ МЕ БЯЕ ЙКЕЫХ ЯНДЕПФЮР БХПСЯ. нРЛЕВЕМН, ВРН БХПСЯ ОЕПЕДЮЕРЯЪ МЕ БЯЕЛС ОНРНЛЯРБС Х МЕ БЯЕЛХ НЯНАЪЛХ ГЮПЮФЕММШУ ЙКЕЫЕИ; БХПСЯ ЯНУПЮМЪКЮ РНКЭЙН ЛЕМЭЬЮЪ ВЮЯРЭ ГЮПЮФЕММШУ НЯНАЕИ (ю. к. дСЛХМЮ, 1958).
бХПСЯНТНПМНЯРЭ ЙКЕЫЕИ ХГЛЕМЪЕРЯЪ Б ПЮГКХВМШЕ ЦНДШ (л. й. рЧЬМЪЙНБЮ, 1958). б гЮОЮДМНИ яХАХПХ Б ПЮИНМЮУ Я МЮХАНКЭЬЕИ ГЮАНКЕБЮЕЛНЯРЭЧ (Б СПЛЮМЕ, ОПЕДЦНП╜МНИ РЮИЦЕ Х Б НЯРПНБМШУ ЯЛЕЬЮММШУ КЕЯЮУ) ГЮПЕЦХЯРПХПНБЮМН Б 1961 Ц. СБЕКХВЕМХЕ ЯРЕОЕМХ БХПСЯНТНПМНЯРХ I. persulcatus МЮ 9 Х 6% ОН ЯПЮБМЕМХЧ Я 1960 Ц. (ц. х. Heu- ЙОИ, р. м. тЕДНПНБЮ, к. ю. лЕКЕМРЭЕБЮ, 1962). б рНЛЯЙНЛ НВЮЦЕ Б НРДЕКЭМШЕ ЦНДШ НРЛЕВЕМШ ЙНКЕАЮМХЪ БХПСЯНТНПМНЯРХ ЙКЕЫЕИ I. persulcatus. нМЮ ПЕГЙН БНГПНЯКЮ Б ОНЯКЕДМХЕ ЦНДШ Я 1,6≈3,2 (1950≈1954) ДН 10,0 (1958), 11,6 (1959) Х 19,5 (1960) (я. о. йЮПОНБ, 1961).
б уЮАЮПНБЯЙНЛ ЙПЮЕ ЩРХ ЙНКЕАЮМХЪ АШКХ МЕГМЮВХРЕКЭМШЛХ МЮ ОПНРЪФЕМХХ 1956≈1960 ЦЦ. (к. ю. бЕПЕРЮ, 1961). б йХПНБЯЙНИ НАКЮЯРХ Б ОПХПНДМШУ НВЮЦЮУ ХМТЕЙ╜ЖХХ БХПСЯНТНПМНЯРЭ ОНКНБНГПЕКШУ I. persulcatus ДНЯРХЦЮКЮ Б 1960 Ц. 8.8%, Б 1961 Ц.≈ 6.4% Х МЕ БШДЕКЕМН МХ НДМНЦН ЬРЮЛЛЮ БНГАСДХРЕКЪ ХГ ЙКЕЫЕИ Б 1962 Х 1963 ЦЦ. (ю. ю. оВЕКЙХМЮ, ю. ю. гЕЛЯЙЮЪ, к. ц. яСБНПНБЮ, 1962). б оЕПЛЯЙНИ НАКЮЯРХ Б ПЮГ╜КХВМШЕ ЦНДШ ХЛЕКХ ЛЕЯРН ПЕГЙХЕ ХГЛЕМЕМХЪ БХПСЯНТНПМНЯРХ ЙКЕЫЕИ ДЮММШЛ БХПСЯНЛ НР 3 ДН 20% (я. ю. ьХКНБЮ, 1961).
б ПЮГКХВМШУ ПЮИНМЮУ НДМНЦН Х РНЦН ФЕ НВЮЦЮ МЮАКЧДЮКХЯЭ ПЕГЙХЕ ЙНКЕАЮМХЪ Б ВХЯКЕММНЯРХ БХПСЯНТНПМШУ ЙКЕЫЕИ. рЮЙ, Б ПЮГМШУ ОСМЙРЮУ ЧФМНЦН ПЮИНМЮ рНЛ╜ЯЙНЦН НВЮЦЮ СЯРЮМНБКЕМН БХПСЯНМНЯХРЕКЭЯРБН I. persulcatus НР 1,6 ДН 1,1%, Б ЖЕМРПЮКЭ╜МНЛ ПЮИНМЕ ≈ 0,42%, Б ЯЕБЕПМНЛ ПЮИНМЕ Б 1952 Ц.≈0,33% Х Б 1953 Ц. БХПСЯНМНЯХ╜РЕКЭЯРБН НРЯСРЯРБНБЮКН." мЮХАНКЕЕ БШЯНЙХИ ОПНЖЕМР ЯНДЕПФЮМХЪ БХПСЯЮ БШЪБКЕМ С ЙКЕ╜ЫЕИ ХГ ЧФМШУ ПЮИНМНБ Х МЮХАНКЕЕ МХГЙХИ С ЙКЕЫЕИ ХГ ЯЕБЕПМШУ ПЮИНМНБ (л. й. рЧЬ╜МЪЙНБЮ, 1955).
б НВЮЦЮУ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ гЮОЮДМН-яХАХПЯЙНИ МХГЛЕММНЯРХ МЮХАНКЕЕ БШЯНЙЮЪ ГЮПЮФЕММНЯРЭ I. PersulcatusoTMeneHa Б ОПЕДЦНПМНИ РЮИЦЕ яЮКЮХПЯЙНЦН ЙПЪФЮ, Б НЯРПНБМШУ ЯЛЕЬЮММШУ КЕЯЮУ 36≈47% Х АШКЮ БДБНЕ ЛЕМЭЬЕ ≈ 14≈20% Б СПЛЮМЕ (ц. х. мЕЖЙХИ Я ЯНЮБРНПЮЛХ, 1962).
бЮПЭХПСЕР ГЮПЮФЕММНЯРЭ ЙКЕЫЕИ Б ПЮГМШЕ ЯЕГНМШ ЦНДЮ. б. л. оНОНБ Х ДП. (1957) СЙЮГШБЮЧР, ВРН Б рНЛЯЙНЛ НВЮЦЕ БХПСЯ ВЮЫЕ Х КЕЦВЕ БШДЕКЪЕРЯЪ ХГ ЙКЕЫЕИ ПЮММЕИ БЕЯМНИ М Б МЮВЮКЕ КЕРЮ.
б яСОСРХМЯЙНЛ ГЮОНБЕДМХЙЕ (сЯЯСПХИЯЙЮЪ НАКЮЯРЭ) Б ОЕПБСЧ ОНКНБХМС КЕРЮ 1939 Ц. АШКН БШДЕКЕМН 27 ЬРЮЛЛНБ БХПСЯЮ ХГ ЙКЕЫЕИ I. persulcatus (м. б. пШФНБ, ю. б. йНГКНБЮ, 1939). б ЙНМЖЕ КЕРЮ РНЦН ФЕ ЦНДЮ ХГ ЙКЕЫЕИ ПЮИНМЮ еЦЕПЯЙНЦН ЙКЧВЮ МЕ СДЮКНЯЭ БШДЕКХРЭ МХ НДМНЦН ЬРЮЛЛЮ БХПСЯЮ (б. д. яНКНБЭЕБ), Ю Б 1940 Ц. РЮЛ ФЕ АШКН БШДЕКЕМН Б ОЕПБНИ ОНКНБХМЕ КЕРЮ 6 ЬРЮЛЛНБ БХПСЯЮ (х. ю. лНЯЙБХМ, 1948).
б йХПНБЯЙНИ НАКЮЯРХ МЮ ОПНРЪФЕМХХ БЯЕЦН ЯЕГНМЮ 1960 ≈1961 ЦЦ. ГЮПЮФЕММШЕ ЙКЕЫХ БЯРПЕВЮКХЯЭ ДНБНКЭМН ПЮБМНЛЕПМН, Х КХЬЭ МЕГМЮВХРЕКЭМНЕ ЯМХФЕМХЕ ХУ НРЛЕ ВЕМН Б ЙНМЖЕ ЯЕГНМЮ (ю. ю. оВЕКЙХМЮ Я ЯНЮБРНПЮЛХ, 1962).
яПЕДХ ДХЙХУ ЛКЕЙНОХРЮЧЫХУ Х ОРХЖ ≈ ОПНЙНПЛХРЕКЕИ ОЕПЕМНЯВХЙНБ ≈ ХГБЕЯРМШ ЛМНЦХЕ БХДШ МНЯХРЕКЕИ БХПСЯЮ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ.
б аЕКНПСЯЯХХ б. х. бНРЪЙНБ Х а. о. яЮБХЖЙХИ БШДЕКЪЧР ДБЮ РХОЮ НВЮ╜ЦНБ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ, ОПХВЕЛ НЯМНБМНЕ ЩОХДЕЛХНКНЦХВЕЯЙНЕ ГМЮВЕМХЕ НМХ НРБНДЪР НВЮЦЮЛ КЕЯМШУ ОЮЯРАХЫ, Ю ЛЮКНЕ ≈ ДХЙХЛ КЕЯМШЛ НВЮЦЮЛ, РЮЙ ЙЮЙ НМХ БЯРПЕВЮЧРЯЪ ГДЕЯЭ ПЕДЙН ≈ РНКЭЙН МЮ РЕППХРНПХХ ГЮОНБЕДМХЙНБ.
яБНЕНАПЮГМШЕ НВЮЦХ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ БШЪБКЕМШ Б вЕУНЯКНБЮЙХХ, бЕМЦПХХ, оНКЭЬЕ, юБЯРПХХ, чЦНЯКЮБХХ. тХМКЪМДХХ, ьБЕЖХХ [сНПЙ (Work, 1964)].
б БНЯРНВМНИ яКНБЮЙХХ СЯРЮМНБКЕМ ОЕПЕМНЯВХЙ Ixodes ricinus (цЮККХЪ Я ЯНРПСДМХЙЮЛХ, 1948, 1949).
яОНМРЮММЮЪ ГЮПЮФЕММНЯРЭ D. marginatus Я МЮКХВХЕЛ РПЮМЯНБЮПХЮКЭМНИ ОЕПЕДЮВХ БХПСЯЮ ДНЙЮГЮМЮ кХАХЙНБНИ Х лЮВХВЙЮ.
мЮ РЕППХРНПХХ оНКЭЬХ (Б цСЮМЭЯЙНЛ, ыЕРХМЯЙНЛ, аЕКНЯРНЙЯЙНЛ БНЕ╜БНДЯРБЮУ Х БАКХГХ ЦПЮМХЖШ Я яяяп) НАМЮПСФЕМШ НВЮЦХ ЙКЕЫЕБНЦН ЩМЖЕТЮ╜КХРЮ Я ОЕПЕМНЯВХЙНЛ I. ricinus (о. б. кЪУЛЮИЯПНБЮ, 1959).
б юБЯРПХХ БШДЕКЕМН 14 ЬРЮЛЛНБ БНГАСДХРЕКЪ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ ХГ 1. ricinus Б 1961 Ц. Х 3 ЬРЮЛЛЮ ≈ Б 1962 Ц., ОПЕХЛСЫЕЯРБЕММН ХГ МХЛТ Х ОНКНБНГПЕКШУ НЯНАЕИ !кНЕБ (Loew, 1963)J. юБРНП ОПХУНДХР Й ГЮЙКЧВЕМХЧ, ВРН РПЮМЯНБЮПХЮКЭМЮЪ ОЕПЕДЮВЮ БНГАСДХРЕКЪ С ЩРНЦН БХДЮ ЙКЕЫЮ ЪБКЪЕРЯЪ ПЕДЙХЛ ЪБКЕМХЕЛ, РЮЙ ЙЮЙ ХГ ХЯЯКЕДНБЮММШУ 143 ОЮПРХИ КХВХМНЙ БШДЕКЕМ РНКЭЙН НДХМ ЬРЮЛЛ БХПСЯЮ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ.
б чЦНЯКЮБХХ Б 1953 Ц. БШЪБКЕМШ НВЮЦХ ГЮАНКЕБЮМХЪ МЮ ЯЕБЕПН-ГЮОЮДЕ ЯРПЮМШ: Б юКЭОЮУ Х Б ГЮОЮДМНЛ оЮММНМХЮМЯЙНЛ АЮЯЯЕИМЕ. бХПСЯ БШДЕКЕМ ХГ ЙКЕЫЕИ I. ricinus [бЕГЕМЭЪЙ (I. Vesenjak Х ДП., 1955); йЛЕР (Kmet Х ДП.,
1955) ; бЕГЕМЭЪЙ, цХПЭЪМ, рНБНПМХЙ, Cooc, 1963; сНПЙ, 1961].
б мХДЕПКЮМДЮУ МЮКХВХЕ ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ ОНДРБЕПФДЕМН БХПСЯН╜КНЦХВЕЯЙХЛХ ХЯЯКЕДНБЮМХЪЛХ [бЕПКЮИМД (Verlinde Х ДП., 1955)]. рНМЦЕПЩМ (Tongeren, 1958), ХГСВЮЪ ЙКЕЫЕИ-ОЕПЕМНЯВХЙНБ, МЕ ЯЛНЦ ОНКСВХРЭ РПЮМ╜ЯНБЮПХЮКЭМСЧ ОЕПЕДЮВС БХПСЯЮ С I. ricinus (ЖХР. ОН сНПЙ, 1961).
нАМЮПСФЕМШ БХПСЯШ Б РПНОХВЕЯЙНИ ГНМЕ ОН ЮМРХЦЕММШЛ ЯБНИЯРБЮЛ, НВЕМЭ ЯУНДМШЕ Я БХПСЯНЛ ПСЯЯЙНЦН ЙКЕЫЕБНЦН ЩМЖЕТЮКХРЮ.
б лЮКЮИЕ БНГАСДХРЕКЭ БШДЕКЕМ ХГ Ixodes granulatus [яЛХР (Smith,
1956) 1. нДМЮЙН МЕР ДНЙЮГЮРЕКЭЯРБ РНЦН, ВРН ДЮММШИ БХПСЯ (Longat virus) ЛНФЕР БШГШБЮРЭ ГЮАНКЕБЮМХЕ КЧДЕИ (ЖХР. ОН бНЙС, 1961).
б хМДХХ, Б ЬРЮРЕ лХЯНП (Mysore) СЯРЮМНБКЕМЮ бНЙНЛ (1958) йЭЪЯЮМСП╜ЯЙЮЪ КЕЯМЮЪ КХУНПЮДЙЮ (й. F. D.), ЙНРНПЮЪ, ОН ЛМЕМХЧ ЮБРНПЮ, ╚ЯУНДМЮ ХКХ ХДЕМРХВМЮ БЕЯЕММЕ-КЕРМЕЛС БХПСЯС╩, Р. Е. ЙКЕЫЕБНЛС ЩМЖЕТЮКХРС Х ОПНЪБ╜КЪЕРЯЪ ГДЕЯЭ РПНОХВЕЯЙНИ БЕЯМНИ ХКХ КЕРНЛ (ЛЕФДС ЪМБЮПЕЛ Х ХЧМЕЛ). оПХ ХЯЯКЕДНБЮМХХ Б 1959 Ц. ХЙЯНДНБШУ ЙКЕЫЕИ МЮ БХПСЯНМНЯХРЕКЭЯРБН МЮХАНКЭ╜ЬЕЕ ВХЯКН ЬРЮЛЛНБ БШДЕКЕМН ГДЕЯЭ ХГ ЦНКНДМШУ ОНКНБНГПЕКШУ Х МХЛТ Haemaphysalis Spinigera, Х РНКЭЙН ДБЮ ЬРЮЛЛЮ ОНКСВЕМН ХГ КХВХМНЙ ЩРНЦН БХДЮ. ,
яКЕДНБЮРЕКЭМН, РПЮМЯНБЮПХЮКЭМЮЪ ОЕПЕДЮВЮ БХПСЯЮ С ЙКЕЫЕИ ЩРНЦН БХДЮ ЪБКЕМХЕ ПЕДЙНЕ. еДХМХВМШЕ БШДЕКЕМХЪ БХПСЯЮ ОНЙЮГЮМШ ХГ Haemaphysalis papuana, II. turturis. лЮЯЯНБНЕ БШДЕКЕМХЕ БХПСЯЮ ХГ H. Spinigera СЙЮГШБЮЕР МЮ БЮФМСЧ ПНКЭ ЩРНЦН ЙКЕЫЮ Б ЯНУПЮМЕМХХ БХПСЯЮ Б ОПХПНДЕ.
щОХДЕЛХНКНЦХВЕЯЙНЕ ГМЮВЕМХЕ ХЙЯНДНБШУ ЙКЕЫЕИ ХГБЕЯРМН Х ОПХ ДПС╜ЦХУ БХПСЯМШУ ХМТЕЙЖХЪУ. б НВЮЦЮУ ЙПШЛЯЙНИ ЦЕЛНППЮЦХВЕЯЙНИ КХУНПЮДЙХ (йцк) МЮ ЯЕБЕПН-ГЮОЮДЕ ЯРЕОМНИ ГНМШ йПШЛЮ АШКЮ ДНЙЮГЮМЮ л. о. вСЛЮЙН╜БШЛ (1945) ЯОНМРЮММЮЪ ГЮПЮФЕММНЯРЭ ОНКНБНГПЕКШУ ЙКЕЫЕИ Hyalomma plumbeum plumbeum БХПСЯНЛ ЩРНИ ХМТЕЙЖХХ. йКЕЫХ АШКХ ЯНАПЮМШ
я. о. оХНМРЙНБЯЙНИ Б ОПХПНДЕ (Я ПЮГКХВМШУ СВЮЯРЙНБ), Я ГЮИЖЕБ Х ЯЙНРЮ.
б гЮОЮДМН-яХАХПЯЙНИ НАКЮЯРХ ХЙЯНДНБШЕ ЙКЕЫХ ЪБКЪЧРЯЪ ОЕПЕМНЯВХ╜ЙЮЛХ нЛЯЙНИ ЦЕЛНППЮЦХВЕЯЙНИ КХУНПЮДЙХ (нцк) Dermacentor pictus (ю. б. цЮЦЮПХМЮ, 1949) Х D. marginatus (ю. б. цЮЦЮПХМЮ, 1954).
аЮЙРЕПХЮКЭМШЕ ГЮАНКЕБЮМХЪ. хЙЯНДНБШЕ ЙКЕЫХ ЪБКЪЧРЯЪ ДКХРЕКЭМШЛХ УПЮМХРЕКЪЛХ БНГАСДХРЕКЕИ МЕЙНРНПШУ АЮЙРЕПХЮКЭМШУ ХМТЕЙЖХИ. мН КХЬЭ МЕЛМНЦХЕ БХДШ ЙКЕЫЕИ ЯОНЯНАМШ ОЕПЕДЮБЮРЭ БНГАСДХРЕКЪ ВЕКНБЕЙС.
рСКЪПЕЛХЪ. б 1928≈1929 ЦЦ. АШКЮ ДНЙЮГЮМЮ ПНКЭ ХЙЯНДНБШУ ЙКЕЫЕИ Б ЩРНИ ХМТЕЙЖХХ. б. м. тЕДНПНБ ОНЙЮГЮК, ВРН 1. aproπophorus Х D. margi╜natus ЛНЦСР БНЯОПХМХЛЮРЭ F. tulareπsis Б ЩЙЯОЕПХЛЕМРЮКЭМШУ СЯКНБХЪУ
Х ЯНУПЮМЪРЭ ОН УНДС ЛЕРЮЛНПТНГЮ. д. ю. цНКНБ (1935) ОЕПБШИ ДНЙЮГЮК, ВРН
D. marginatus ЪБКЪЧРЯЪ ОЕПЕМНЯВХЙНЛ F. tularensis. нМ СЯРЮМНБХК ЯНУПЮМЕ╜МХЕ БНГАСДХРЕКЪ ОН УНДС ЛЕРЮЛНПТНГЮ С ЩЙЯОЕПХЛЕМРЮКЭМН ГЮПЮФЕММШУ ЙКЕЫЕИ D. marginatus, НАМЮПСФХК ЕЦН Б ЩЙЯЙПЕЛЕМРЮУ МХЛТ (ГЮПЮФЕММШУ Б ТЮГЕ КХВХМНЙ) Х СЯРЮМНБХК БШФХБЮЕЛНЯРЭ Б ОНКНБНГПЕКШУ ЙКЕЫЮУ ЩРНЦН БХДЮ МЮ ОПНРЪФЕМХХ 530 ДМЕИ.
б ПЕГСКЭРЮРЕ ЛМНЦНКЕРМХУ МЮАКЧДЕМХИ ГЮ ОПХПНДМШЛ НВЮЦНЛ Б ДЮММНИ НАКЮЯРХ м. ц. нКЯСТЭЕБ НАНЯМНБЮК ГМЮВЕМХЕ D. pictus Б ЩОХГННРНКНЦХХ РСКЪ╜ПЕЛХХ .
яН БПЕЛЕМХ ЩРХУ ХЯЯКЕДНБЮМХИ ЯРЮКН ХГБЕЯРМН 14 БХДНБ ХЙЯНДН╜БШУ ЙКЕЫЕИ ≈ МНЯХРЕКЕИ F. tularensis Б ОПХПНДЕ, Ю ХЛЕММН: 1. ricinus (м. ц. нКЯСТЭЕБ, 1943), I. persulcatus (я. о. йЮПОНБ, б. л. оНОНБ, 1944), 1. Iaguri Iaguri (б. о, пНЛЮМНБ Х ДП., 1949), 1. aproπophorus (д. ю. цНКНБ, 1934), Haeniaphysalis concinna (м. ц. нКЯСТЭЕБ Х ДП., 1959), H. punctata
H. sulcata (Я ЙНГ Х БНКЙНБ)
х. т. рЮПЮМ, 1959
х. л. цПНУНБЯЙЮЪ Я ЯНЮБРНПЮЛХ.
D. marginatus (Я ОЮЯР╜АХЫ)
D. marginatus (Я ЙНПНБ)
б. х. йСПВЮРНБ Я ЯНЮБРНПЮЛХ, 1955
рН ФЕ (Я БНКНБ)
б. м. йСПВЮРНБ Я ЯНЮБРНПЮЛХ, 1955
H. Scupense