ВИДОВОЙ СОСТАВ ФАУНЫ ИКСОДОВЫХ КЛЕЩЕЙ (IXODIDAE) РОССИИ И СОПРЕДЕЛЬНЫХ ГОСУДАРСТВ
(по: Филиппова, 1984, с изменениями)
Amblyomma
1. A. gemma Donitz, 1909 Anomalohimalaya
2. A. lama Hoogstraal, Kaiser et Mitchell, 1970
3. A. lotozkyi Filippova et Panova, 1978
Boophilus
4.
* B. annulatus Say, 1821 (В. calcaratus Birula, 1895)Dermacentor
5. D. antrorum Resnik, 1950
6. * D. marginatus Sulzer, 1776
7. D. montanus Filippova et Panova, 1974
8. * D. niveus Neumann, 1897 (D. daghestanicus Olenev, 1929)
9. * D. nuttalli Olenev, 1927
10. D. pavlovskyi Olenev, 1927
11. * D. pomerantzevi Serdjucova, 1951
12. D. raskemensis Pomerantzev, 1946
13. * D. reticulatus Fabricius, 1794 (D. pictus Hermann, 1804)
14. * D.silvarum Olenev, 1927
15. D. ushakovae Filippova et Panova, 1989
Haemaphysalis
16. Я. caucasica Olenev, 1928
17. * Я. concinna Koch, 1844
18. * Я. erinacei Pavesi, 1884 (Я. numidiana Neumann, 1897)
19. * Я filippovae Bolotin, 1979
20. * H. flava Neumann, 1897
21. * H. inermis Birula, 1895
22. * H. japonica Warburton, 1908
23. H. kopetdagihca Kerbabaev, 1962
24. H. kutchensis Hoogstraal et Trapido, 1963
25. * H. longicornis Neumann, 1901 (#. neumanni Donitz, 1905)
26. * H. ornithophila Hoogstraal et Kohls, 1979
27. * H. parva Neumann, 1897 (Я. otophila Schulze, 1918)
28. H. pavlovskyi Pospelova-Shtrom, 1935
29. * H. phasiana Saito, Hoogstraal et Wassef, 1954
30. H. pospelovashtromae Hoogstraal, 1966 (H. warburtoni Nuttall, 1912)
31. * Я. punctata Canestrini et Fanzago, 1877
32. * Я. sulcata Canestrini et Fanzago, 1877
33. * Я. wellingtoni Nuttall et Warburton, 1908
Hyalomma
34. * Я. aegyptium (Linnaeus, 1758)
35. * Я. anatolicum Koch, 1844
36. Я asiaticum Schulze et Schlottke, 1929
37. Я. detritum Schulze, 1919
38. Я. dromedarii Koch, 1844
39. Я. impressum Koch, 1844
40. Я. kozlovi Olenev, 1934
41.
Я. kumari Sharif, 192842. * Я. marginatum Koch, 1844
43. Я schulzei Olenev, 1931
44. * Я. scupense Schulze, 1981
Rhipicephalus -
45. * R. bursa Canestrini et Fanzago, 1877
46. R. leporis Pomerantzev, 1946
47. * R. pumilio Schulze, 1935
48. * /?. rossicus Jakimov et Kohl-Jakimova, 1911
49. * R. sanguineus (Latreille, 1806)
50. * R. schulzei Olenev, 1929
51. * R. turanicus Pomerantzev, 1946
Ixodes
52. * /. angustus Neumann, 1899
53. * /. apronophorus Schulze, 1924
54. * /. arboricola Schulze et Schlottke, 1929
55. * /. berlesei Birula, 1895
56. /. bruneus Koch, 1844
57. * /. caledonicus Nuttall, 1910
58. /. cornutus Lotozkyi, 1956
59. * /. crenulatus Koch, 1844
60. I. eldaricus Djaparidze, 1950
61. * /. frontalis (Panzer, 1798)
62. /. hexagonus Leach, 1815
63. /. kaiseri Arthur, 1957
64. /. kashmiricus Pomerantzev, 1948
65. /. kazakstani Olenev et Sorokoumov, 1934
66. * /. laguri Olenev, 1929
67. * /. lividus Koch, 1844
68. * /. maslovi Emeljanova et Kozlovskaya, 1967
69. * /. nipponensis Kitaoka et Saito, 1967
70. * /. occultus Pomerantzev, 1946
71. * /. pavlovskyi Pomerantzev, 1946
72. * /. persulcatus Schulze, 1930
73. * I. pomerantzevi Serdjukova, 1911
74. * /. redikorzevi Olenev, 1927
75. * /. ricinus (Linnaeus, 1758)
76. * /. sachalinensis Filippova, 1957
77. /. semenovi Olenev, 1929
78. * /. signatus Birula, 1895
79. * I. simplex Neumann, 1906
80. * /. strorqi Filippova, 1957
81. I. subterraneus Filippova, 1961
82. * /. trianguliceps Birula, 1895
83. * /. turdus Nakatsuji, 1943
84. /. unicavatus Neumann, 1908
85. * /. uriae White, 1852
86. * /. vesertilionis Koch, 1884
Примечание. * — виды, встречающиеся на территории России.
странению клещей отсутствуют. Однако существуют значительные различия в видовом составе фаун иксодид европейской части России и Дальнего Востока, разделенных пространствами Сибири с бедной фауной клещей.
Видовые ареалы иксодид в наибольшей степени зависят от гигротермических условий окружающей среды и определяемых ими типов растительного покрова. Соответственно в широтном направлении с севера на юг разнообразие видового состава повсеместно увеличивается. Прослеживается четкая евязь многих видов иксодид с определенными типами ландшафтов.Дальше всех на север проникает I. игіае, заселяющий побережья Баренцева и Белого морей. Восточнее Новой Земли этот вид не проникает из-за суровости климата и снова появляется на побережье и островах морей Тихого океана. Клещи обитают в колониях морских птиц, часто образующих крупные скопления («птичьи
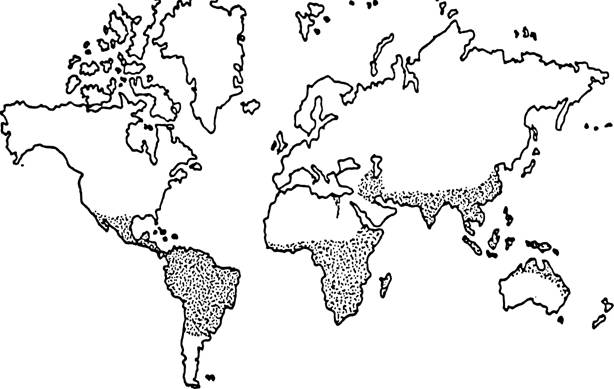
Рис. 31. Ареал клеща Boophilus microplus (точечная штриховка) (по: Nunez et al.t 1985).
базары»), В этих местообитаниях клещи заселяют многочисленные трещины и щели в скалах или же находятся в субстрате оснований гнезд. На островах Белого моря клещи встречаются в вырытых в земле гнездах чистиков. Местами численность клещей бывает очень высокой. На побережье Тихого океана встречается также клещ I. signatus, обладающий сходной экологией и часто встречающийся вместе с /. игіае. В зоне тундры иксодовые клещи отсутствуют.
В лесной зоне России достаточно часто встречается около 18 видов иксодид. Из них пастбищный тип паразитизма свойствен I. persulcatus, /. ricinus, /. pav- lovskyi, /. nipponensis, D. reticulatus, D. silvarum, H. concinna, H. inermis, H. ja- ponica, H. longicornis (Арзамасов, 1961; Филиппова, 1977; Бойко и др., 1982; Успенская, 1987). Эти виды подстерегают своих хозяев, поднимаясь на растительность или же оставаясь на поверхности почвы. Наиболее широким распространением обладает таежный клещ I. persulcatus. Он встречается в среднетаежных, южнотаежных и широколиственных лесах от западных границ России до побережья и островов Тихого океана, включая юг Камчатки, а также в Северном Китае, Корее и Японии (рис. 32). В этих границах,клещи отсутствуют на заболоченных территориях, в сухих сосняках на песчаных почвах, а также на молодых вырубках и открытых лугах.
На Сахалине в условиях высокой влажности воздуха клещи встречаются на закустаренных лугах и вырубках с высоким травостоем. Распространение клещей на север совпадает с изолиниями сумм температур воздуха 1400 °С за период с устойчивой среднесуточной температурой выше 10 °С или суммой температур 1600 °С за период со среднесуточной температурой выше 5 °С. Оптимальные же условия существования этого вида складываются при сумме температур более 2300 °С и коэффициентом увлажнения 0.45—0.60. В северной части ареала таежный клещ встречается только в ограниченных, наиболее теплообеспеченных местообитаниях. В европейской части ареала северная граница распространения вида проходит вблизи 62—63° с. ш., на Урале и в Сибири — около 60° и в Хабаровском крае между 52—54° с. ш. Южная граница ареала в европейской части России проходит от 56° с. ш. на западе до 53° на восток от Волги. В Сибири кроме зоны южной тайги клещи встречаются в изолированных лесных массивах лесостепи и по долинам рек и многочисленны в горных лесах Южной Сибири, Казахстана и Киргизии. На Дальнем Востоке южная граница ареала уходит на территорию Китая (Коренберг, 1979; Таежный клещ..., 1985). В горных лесах Южной Сибири и Дальнего Востока (рис. 32) вместе с таежным клещом встречается и другой лесной вид — I. pavlovskyi, отличающийся от первого вида преимущественным паразитированием взрослых особей на птицах.Ixodes ricinus распространен от Англии и Ирландии на западе до Волги на востоке (рис. 32). Во многих областях России к западу от Волги I. ricinus встречается вместе с /. persulcatus. Распространение этого вида на север ограничивается дефицитом тепла и совпадает с северной границей среднетаежных лесов. На юге клещи проникают за границы широколиственных лесов и многочисленны в зоне лесостепи и степи в разных типах искусственных лесонасаждений. Клещи также многочисленны в горных лесах Кавказа, Турции, Южной Европы и Северной Африки, где заселяют более влагообеспеченные леса на северных склонах и в речных долинах.
Наиболее высокой численности этот вид достигает в зоне широколиственных лесов, но он достаточно многочислен и в расположенных севернее смешанных и южнотаежных лесах Скандинавского полуострова и России. I. ricinus легко приспосабливается к антропогенной трансформации ландшафта и заселяет разного типа вторичные леса среди сельскохозяйственных угодий (Померанцев, 1935). В условиях влажного климата Англии и Ирландии этот вид встречается не только в лесах, но также на открытых пастбищах, вересковых пустошах и в зарослях кустарников (Varma, 1965).В широколиственных лесах Европы, включая юг России и Кавказ, обитают Н. inermis и Н. сопсіппа. Второй из этих видов встречается также местами в лиственных лесах Тянь-Шаня, Алтая, Саян и многочислен в широколиственных лесах юга Дальнего Востока (рис. 33). В лесах последнего региона встречается еще несколько видов Haemaphysalis, из которых наиболее обычны Н. japonica и Н. longicornis. На увлажненных луговых стациях прибрежной полосы в Южном Приморье встречается I. nipponensis, более многочисленный в Японии. В осветленных лесах, на лесных полянах, лугах и в кустарниковых зарослях по всему Дальнему Востоку и югу Сибири до Оби на западе распространен D. silvarum
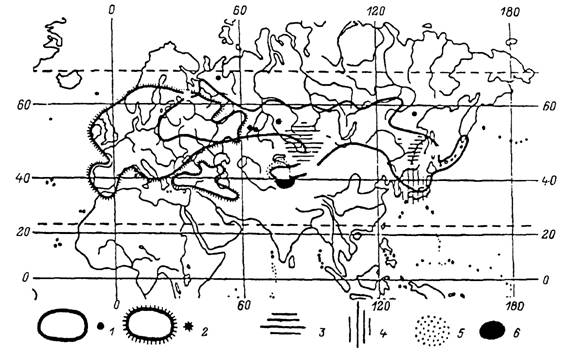
Рис. 32. Распространение видов комплекса Ixodes ricinus в Палеарктике (по: Filippova, 1991). / — /. persulcatus; 2 — I. ricinus', 3 — /. pavlovskyi; 4 — /. nipponensis; 5 — /. kazakstani; 6 — I. kashmiricus.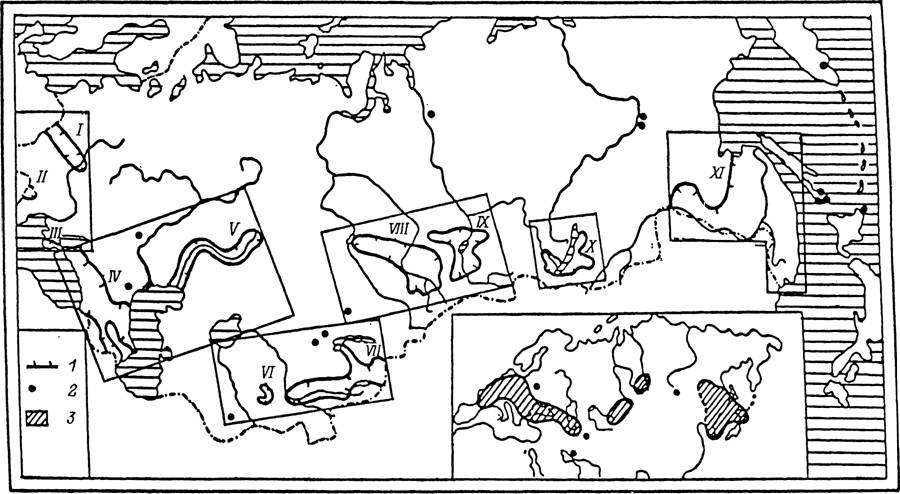
Рис. 33. Распространение клеща Haemaphysalis сопсіппа в России и сопредельных странах (по: Lebedeva, Korenberg, 1981).
/ — гипотетические границы ареала; 2 — места сбора клешей вне границ ареала: 3 — ареал вида в Палеарктике. І—ХІ — географические названия разрозненных частей ареала: /— Припять—Днепр; И — Прут—Днестр; III — Крым; IV — Кавказ; V — Южный Урал; VI — Заревшан; VII — Памир—Тянь-Шань; VIII — Алтай; IX — Саяны; X — Байкал; XI — Дальний Восток.
• 1
-------------------------------------------------------------------------------- 2
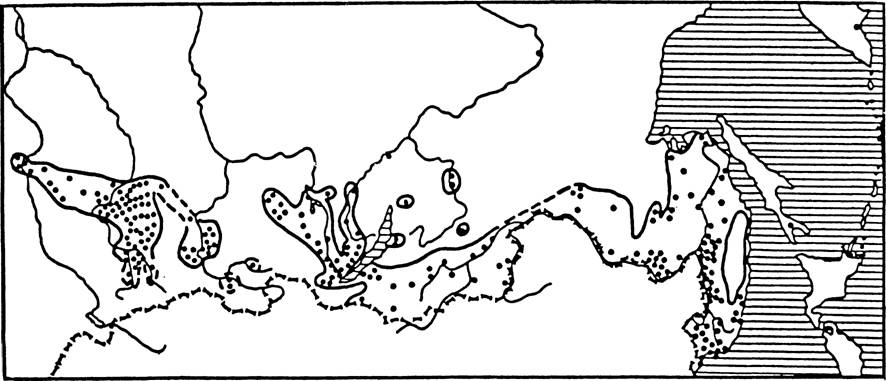
Рис.
34. Ареал клеша Dermaccntor silvorum на территории бывшего СССР (по: Кулик, Винокурова, 1983а). / — места находок клещей; 2 — установленная граница ареала; 3 — предполагаемая граница ареала.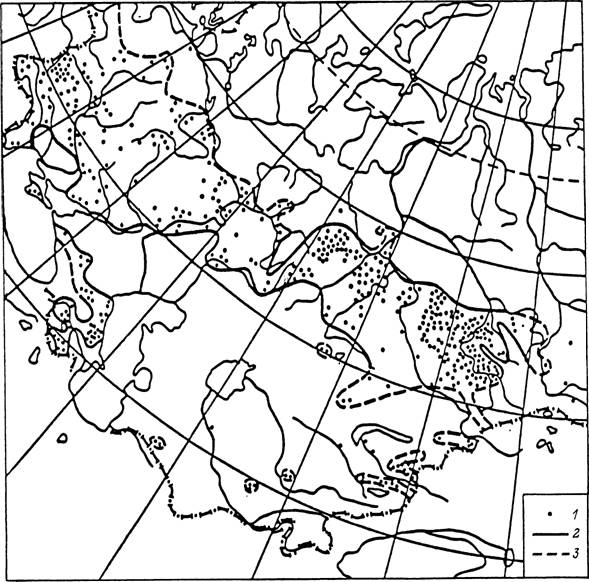
Рис. 35. Ареал клеща Dermacentor reticulatus на территории бывшего СССР (по: Кулик, Винокурова,
19836).
Обозначения как на рис. 34.
(рис. 34). В зоне смешанных и широколиственных лесов Европы и Западной Сибири в луговых стациях, включая пойменные луга, в редколесьях и на вырубках многочислен D. reticulatus (рис. 35).
Гнездово-норовым типом паразитизма обладают клещи /. trianguliceps, /. арго- nophorus, /. crenulatus, /. lividus, /. arboricola, /. kaizeri, /. angustus, /. pomeran- zevi. Клещ /. trianguliceps распространен в смешанных и широколиственных лесах от Англии до Енисея. Этот вид соединяет особенности пастбищного и гнездовонорового паразитизма. Клещи всю свою жизнь проводят внутри лесной подстилки в пронизывающих ее многочисленных ходах мелких млекопитающих. Голодные нимфы и самки этого вида могут подстерегать хозяев и на поверхности почвы, но не поднимаются на растительность. В лесах Дальнего Востока обитателями нор мелких млекопитающих являются /. angustus и /. pomeranzevi. В древесных гнездах птиц клещи встречаются только в зоне широколиственных лесов европейской части России и юга Дальнего Востока, гйе обитает относительно немногочисленный /. arboricola.
По поймам рек и заболоченным берегам озер от побережья Белого моря и до устьев рек юга России и Казахстана вплоть до Средней Сибири встречается /. ар- ronophorus. Для этого вида характерно интразональное распространение, и его ареал охватывает лесную и степную зоны. Он заселяет норы связанных с водоемами главных прокормителей — водяной полевки и ондатры. Вторым интразональным видом является /. lividus, заселяющий гнезда ласточек-береговушек. Он распространен по всей Палеарктике от Атлантичекого до Тихого океана и по долинам рек встречается в полосе от 62° до 43° с. ш.
Видовой состав фауны клещей степной зоны значительно разнообразнее такового лесной зоны (Емчук, 1960; Миронов и др., 1965; Успенская, 1987). Часть из встречающихся в ней видов (/. apronophorus, I. lividus) отличается интразональным распространением, а в небольшие островки леса и искусственные лесонасаждения проникают с севера некоторые из типично лесных видов (/. ricinus, /. persulcatus, D. reticulatus, D. silvarum). Отличительной особенностью степной фауны является широкое распространение гнездово-норовых паразитов. От юга лесной зоны по всей степной зоне и горным степям Кавказа, Средней Азии и Южной Сибири в полосе от 54° до 35° с. ш. встречается /. crenulatus, заселяющий постоянные норы более 30 видов млекопитающих из числа хищников и сурков. В норах сусликов, хомяков и других грызунов на Украине, в южных областях России и в Западном Казахстане многочислен /. laguri. В норах сусликов и мелких хищников на юге европейской России, в Закавказье, Казахстане и Средней Азии часто встречается R. schulzei.
Характерными обитателями степей являются несколько видов с пастбищным типом паразитизма. В степных ландшафтах Европы, включая территорию России на восток до рек Томь и Катунь, обычен и многочислен D. marginatus (рис. 36). Этот же вид распространен в степных ландшафтах гор Кавказа и Средней Азии. В степях долины Енисея и особенно в Забайкалье обитает D. nuttalli, многочисленный также в Монголии и Северном Китае. В камышовых и кустарниковых зарослях пойм рек в аридных частях Ставропольского края, Дагестана, Закавказья, Средней Азии и Казахстана многочислен и часто встречается D. niveus. Из видов рода Haemaphysalis в степной зоне распространены Н. punctata и Н. sulcata, особенно часто встречающиеся в горных ландшафтах. Из рода Rhipicephalus дальше всех проникает на север R. rossicus, встречающийся на Северном Кавказе, в Нижнем Поволжье, Северном Казахстане и Закавказье. В Саратовской, Ростовской и Астраханской областях, Ставропольском и Краснодарском краях, а также в Дагестане в степных и полупустынных ландшафтах встречаются Н. marginatum и Н. scupense. Особенно широко распространен первый вид, напитавшиеся нимфы которого заносятся далеко за границы ареала, от Поволжья и до островов на Балтийском море. Н. scupense паразитирует только на крупном рогатом скоте и имеет более ограниченное распространение.
Наиболее богата фауна южных степей, полупустынь и аридных предгорий Кавказского хребта (Ганиев, 1970). В Астраханской области и Дагестане появляется R. ритіііо, более многочисленный в расположенных южнее пустынях и полупустынях Средней Азии и Казахстана. На овцах в Дагестане паразитирует двуххозя- инный клещ — R. bursa, встречающийся также в Крыму, Краснодарском, Ставропольском краях. На Черноморском побережье Кавказа и Крыма на собаках паразитирует R. sanguineus. Только из Дагестана известны обычные для Закавказья и Средней Азии R. turanicus и Н. anatolicum, а в местообитаниях степной черепахи встречается паразитирующий на них Н. aegyptium. В поймах и заболоченностях в Краснодарском крае и Дагестане местами встречается однохозяинный паразит крупного рогатого скота — В. annulatus. Для трех последних видов приводимые выше местонахождения в России являются крайними северными частями их ареалов. В целом для фауны степей характерно увеличение видового состава с севера на юг и обеднение с запада на восток.
В России полупустынные и пустынные ландшафты ограничены аридными районами Астраханской области, Калмыкии, Ставропольского края и Дагестана. Видовой состав фауны иксодид этих районов обеднен по сравнению с таковым южных степейл В расположенном южнее обширном поясе полупустынь и пустынь Казахстана и Средней Азии видовое разнообразие увеличивается в южном направ-
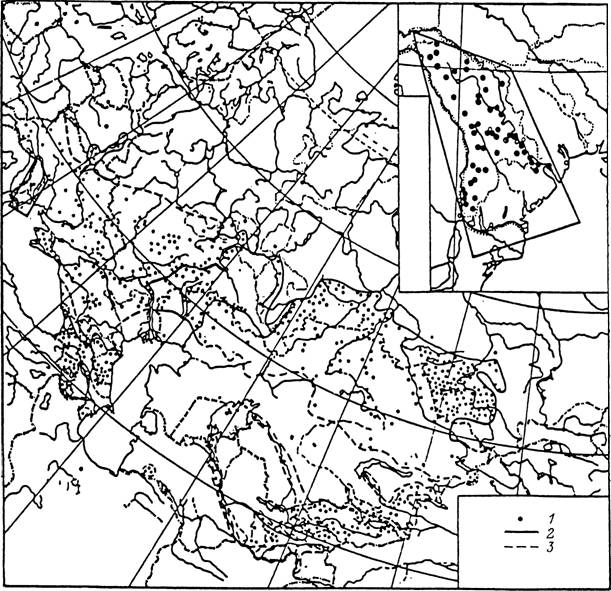
Рис. 36. Ареал клеща Dermacentor marginatus на территории бывшего СССР (по: Кулик, Винокурова,
1982).
Обозначения как на рис. 34.
пении и достигает максимума в пустынях Туркмении (Галузо, 1947, 1948, 1950; Бердыев, 1980). В закрепленных песках, заросших саксаулом, отмечено 13 видов. Из пастбищных паразитов доминирует Н. asiaticum, широко распространенный и в других типах пустынных ландшафтов. Он встречается от Восточного Закавказья и Ирана на западе до пустыни Гоби на востоке и проникает на север до Урало- Эмбинского междуречья в Казахстане. Реже в песках встречаются Н. dromedarii и другие виды этого рода. Личинки и нимфы этого вида обитают в норах своих прокормителей — песчанок и других мелких млекопитающих, а взрослые — на верблюдах, крупном рогатом скоте, овцах и диких копытных. В норах грызунов и хищников встречаются /. occultus, /. redikorzevi, I. crenulatus, H. erinacei, R. schul- zei. В глинистых пустынях Туркмении обнаружено 19 видов, многие из которых встречаются и в песках. Кроме Н. asiaticum на поверхности почвы здесь встречаются Н. anatolicum и Н. detritum, а на растительности — R. turanicus, Н. punctata, Н. sulcata, а из норовых видов многочислен /. subterraneus.
Богата фауна долин рек, протекающих по пустыне. Благодаря стыку пустынных ландшафтов с тугайными лесами, зарослями кустарников и камышей значительно возрастает разнообразие местообитаний, и в них встречается около 23 видов ик- содид. Из них многочисленны Н. anatolicum, Н. dromedarii, Н. detritum, Н. marginatum, В. annulatus, D. niveus, R. pumilio, R. turanicus.
Наиболее богата фауна клещей горных ландшафтов Закавказья и Средней Азии. Особенно разнообразна фауна гор Копетдага, насчитывающая 31 вид (Бердыев, 1980). Наряду с пустынными в ней много степных и горных видов. Особенно многочисленны в остепненных биотопах и стациях с кустарниково-древесной растительностью клещи R. turanicus, D. marginatus, D. niveus, H. sulcata, H. punctata, H. otophila, H. plumbeum, H. aegyptium, I. eldaricus, I. crenulatus. В местах концентрации крупного и мелкого рогатого скота обычны Н. anatolicum, Н. detritum, R. bursa. В расположенных восточнее горных хребтах Памира и Тянь-Шаня вследствие большой высоты хребтов наблюдается большая вертикальная дифференциация фаунистических комплексов клещей (Гребенюк, 1966). В нижних поясах гор с пустынными и степными ландшафтами доминируют Н. marginatum, Н. scupense,
H. anatolicum, Н. asiaticum, В. annulatus, R. turanicus, Н. sulcata. В зоне орехоплодовых лесов и кустарников многочисленны Н. punctata, D. marginatus, I. subterra- neus, H. scupense, H. marginatum, R. turanicus. В еловых лесах, произрастающих на северных склонах и в ущельях Тянь-Шаня, доминирующим видом является
I. persulcatus. В субальпийском и альпийском поясах на высоте 2500 м и более доминируют I. pavlovskyi, I. crenulatus, Н. warburtoni.
Еще по теме ВИДОВОЙ СОСТАВ ФАУНЫ ИКСОДОВЫХ КЛЕЩЕЙ (IXODIDAE) РОССИИ И СОПРЕДЕЛЬНЫХ ГОСУДАРСТВ:
- ВИДОВОЙ СОСТАВ ФАУНЫ ИКСОДОВЫХ КЛЕЩЕЙ (IXODIDAE) РОССИИ И СОПРЕДЕЛЬНЫХ ГОСУДАРСТВ
- Природная очаговость клещевых инфекций