ВНЕШНЕЕ СТРОЕНИЕ
Иксодоидные клещи — наиболее крупные формы среди Асагіпа, и размеры тела у напитавшихся особей отдельных видов (роды АтЫуотта, Hyalomma, Ornithodoros savignyi) достигают 20—30 мм.
Форма тела голодных клещей — от овальной до почти округлой и сильно уплощенная, а у напитавшихся — линзовидная, яйцевидная или каплевидная (рис. 1—7).На основании исследований сегментального строения тела у разных групп клещей А. А. Захваткин (1952) считает, что в основе организации иксодоидных клещей, как и других представителей отряда Parasitiformes, лежит арахноидный тип расчленения на просому (головогрудь) и опи- стосому (брюшко). Просома включает акрон (первичная головная лопасть) и 6 сегментов (хелидеральный, пальпальный и 4 ножных). Опи- стосома лишена конечностей и включает 9 сегментов и анальную лопасть. Из них прегенитальный сегмент в значительной степени редуцирован и отходит к просоме, а 3 последних сегмента остаются в зачаточном состоянии и объединены в один анальный макросомит. В результате брюшко клещей фактически состоит только из 6 сегментов.
У иксодоидных клещей произошло слияние просомы с идиосомой, и границы между ними исчезли. Следы первичной метамерии обнаруживаются у них только в эмбриональном развитии (Вагнер, 1894; Aeschli- mann, 1958) и в расположении хет у личинок. Взамен этого первичного тагмозиса у них развилось вторичное расчленение тела на лишенное сегментации туловище, или идиосому, и головку, или гнатосому (рис. 4, 10, И).
В состав гнатосомы входят часть акрона, хе лидера льный и паль- пальный сегменты, но границы между ними неразличимы в силу высокой степени интеграции головки и общей для паукообразных тенденции изгибания главной оси тела на спинную сторону. Граница между гнато-
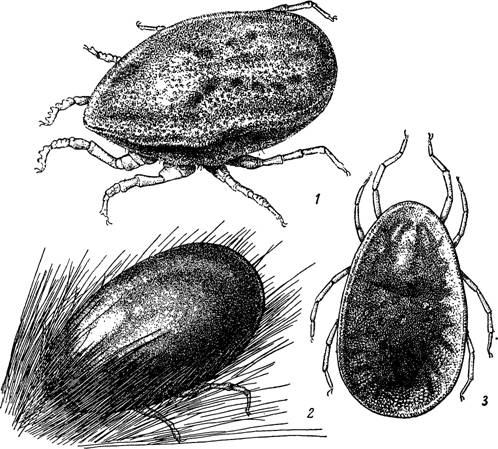
Рнс. 1—3.
1 — голодная самка Alveonasus lahorensis; 2 — заканчивающая питание самка Ornithodoros papillipes; 3 — голодная самка Argas persicus.
сомой и идиосомой проходит внутрисегментально, так что в состав последней входит часть акрона, несущая глаза.
Это послужило поводом для отрицания самостоятельного существования гнатосомы как части тела (Ланге, 1962), что не имеет достаточных морфологических оснований.Внешнее строение клещей надсем. Ixodoidea и особенно структуры, имеющие таксономическое значение, достаточно подробно описаны в сводке по систематике этой группы (Nutall a. oth., 1908; Nutall a. War- burton, 1911, 1915; Robinson, 1926; Cooley a. Kohls, 1944; Галузо, 1946—1953; Померанцев, 1950; Поспелова-Штром, 1953; Arthur, 1960, 1962; Филиппова, 1966, и др.). В настоящем разделе рассматриваются лишь наиболее существенные особенности внешнего строения клещей, необходимые для характеристики группы в целом и затрагиваемые в последующих разделах монографии.
Идиосома. Среди аргасовых клещей различают два главных тина строения идиосомы. У представителей подсем. Argasinae она сильно уплощена, а дорсальная и вентральная поверхности резко отграничены друг от друга боковым швом (рис. 3). У Ornithodorinae боковой шов, за немногими исключениями, отсутствует, тело более выпуклое, и хорошо развиты его боковые поверхности (рис. 9). Поверхность идиосомы голодных особей покрыта системой борозд, наиболее развитых у видов рода Ornithodoros (рис. 8—10). У них различают следующие борозды: дорсальную, маргинальную, парные дорсо-вентральные и продольные супра- коксальные, 2 пары продольных субкоксальных, околоанальные (преанальную, поперечную постанальную и срединную аномаргинальную). У видов рода Avgas число борозд значительно меньше.
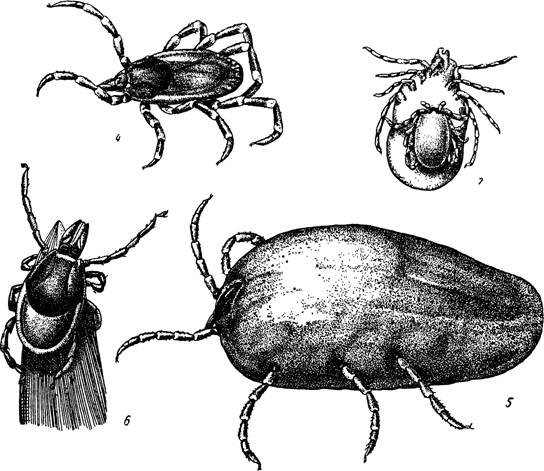
Рис. 4—7.
4 — голодная и 5 — напитавшаяся самка Hyalomma asiaticum; 6 — самка Ixodes persulcatus в позе ожидания (по Померанцеву и Сердюковой, 1948); 7 — спаривание клещей Ixodes ricinus.
Большая часть идиосомы аргазид покрыта растяжимой кутикулой. Твердая кутикула представлена мелкими участками — дисками, служащими для прикрепления мышц (рис. 8) и особенно развитыми у видов рода Argas. Остатки спинного щитка (дорсальная пластинка) сохраняются только у личинок родов Argas, Alveonasus и некоторых видов рода Ornithodoros, тогда как у нимфальной и имагинальной фаз он полностью редуцирован.
На вентральной стороне лежат поперечная щель полового отверстия и далее назад — анальное отверстие. Последнее состоит из двух полукруглых створок и окружающего их кольца из склеротизован- ной кутикулы. В заднелатеральном направлении от III—IV пар кокс лежат небольшие дыхательные пластинки — перитремы, на которых открываются дыхательные отверстия — стигмы. Между I и II парами кокс открываются щелевидные отверстия коксальных желез. Передняя часть идиосомы прикрывает смещенную вентрально гнатосому и образует вокруг нее футляр — камеростом (рис. 10).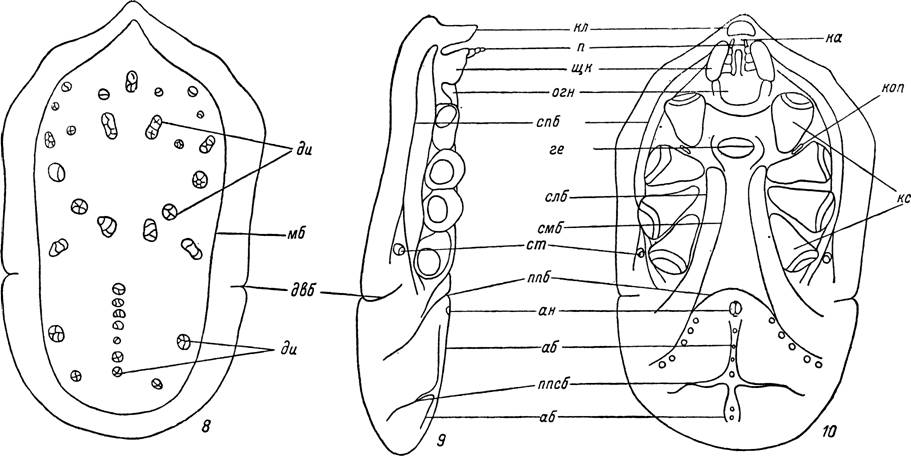
Рис. 8—10. Схема внешнего строения самки Ornithodoros papillipes,
8 — вид сверху, 9 — сбоку, 10 — снизу (по Филипповой, 1966).
он — анус; аб — аномаргинальная борозда; ге — генитальная щель; ди — диски; двб — дорсо-вентральная борозда; ка — камеростом; пл — клюв; коп — коксальная пора; кс — коксы; мб — маргинальная борозда; огп — основание гнатосомы; п — пальпы; ппб — поперечная преанальная борозда; ппсб — поперечная постанальная борозда; слб — субкоксальная латеральная борозда; сліб — субкоксальная медиальная борозда; спб —
супракоксальная борозда; cm — стигма; щк — щеки.У иксодовых клещей наблюдаются заметные различия в строении идиосомы у самок и самцов. У голодных особей идиосома сильно уплощенная, но после питания она сильно растягивается и становится на поперечном разрезе почти округлой.
У самок около трети дорсальной поверхности идиосомы голодных особей занимает спинной щиток (скутум) из твердой кутикулы. Спереди он имеет глубокую вырезку для причленения гнатосомы, а его передне- боковые выступы называют скапулами. На скутуме различают парные боковые и цервикальные борозды. Растяжимая часть спинной поверхности по бокам и сзади от скутума называется аллоскутумом (рис. 11), и она лишена каких-либо склеритов. Задний край идиосомы, исключая виды родов Ixodes и Haemaphy salts, несет 9—11 краевых фестонов. На вентральной стороне лежат половое и анальное отверстия, а по бокам позади IV кокс — крупные перитремы.
На аллоскутуме различают наиболее глубокую дорсальную краевую борозду, парные задне-срединные, задне-боковые, внешние и внутренние добавочные и непарную заднесрединную борозды. С вентральной стороны развиты генитальная, анальная и средняя постанальная борозды (рис. 12). У разных видов наблюдаются значительные вариации в расположении и числе описываемых борозд (рис. 11—20).Внешнее строение идиосомы нимф и личинок в основных чертах сходно с самками. У них лишь относительно большие размеры спинного щитка, меньшее количество борозд, а на вентральной стороне отсутствует половое отверстие. У личинок, кроме того, неразвиты перитремы.
У самцов иксодид спинной щиток покрывает всю дорсальную поверхность идиосомы, так что растяжимая кутикула занимает только узкую латеральную зону (рис. 21, 22). Вентральная сторона покрыта группой щитков. У видов рода Ixodes щитки занимают всю вентральную часть идиосомы, и среди них различают парные аданальные, эпимеральные и непарные прегенительный, срединный и анальный (рис. 13). У АтЫуот- тіпае различают парные аданальные и добавочные щитки, одну или две пары субанальных (рис. 14). Между щитками находятся значительные участки из растяжимой кутикулы.
Гнатосома. Гнатосома сильно склеротизована и состоит из проксимального кольца — основания гнатосомы (basis capituli) и сильно вытянутой дистальной половины — хоботка, или ротового конуса. Хоботок образован гипостомом и наружными хелицеральными футлярами, проксимально переходящими в сплошное склеротизованное кольцо основания гнатосомы (рис. 23, 24 и 197), которое сужается в проксимальном направлении и входит в широкое камеростомальное отверстие идиосомы. С последней гнатосома сочленяется изогнутой камеростомальной складкой. В ней отсутствуют склеротизованные слои, и она состоит только из эпикутикулы и хорошо развитой слоистой эндокутикулы. Дорсо-медиально в камеростомальной складке открывается щелевидное отверстие органа Жене (рис. 27). Основание гнатосомы широким затылочным отверстием открывается в полость тела.
К заднему дорсальному и вентральному краям основания у иксодид прикрепляются две группы мышц, идущие назад и вверх и прикрепляющиеся к спинному щитку в области цервикальной борозды (рис. 25). У аргазид выдвижение и втягивание гнатосомы в камеростомальную складку происходит с помощью группы мышц, идущих от гнатосомы к дискам дорсальной поверхности идиосомы.Нижняя половина хоботка образована вытянутым вперед выростом основания гнатосомы — гипостомом. Снаружи он покрыт направленными назад зубцами, образующими правильные продольные ряды. Зубцы на вершине гипостома наиболее мелкие и у некоторых видов образуют характерную структуру, называемую короной. У самцов гнездово-норо-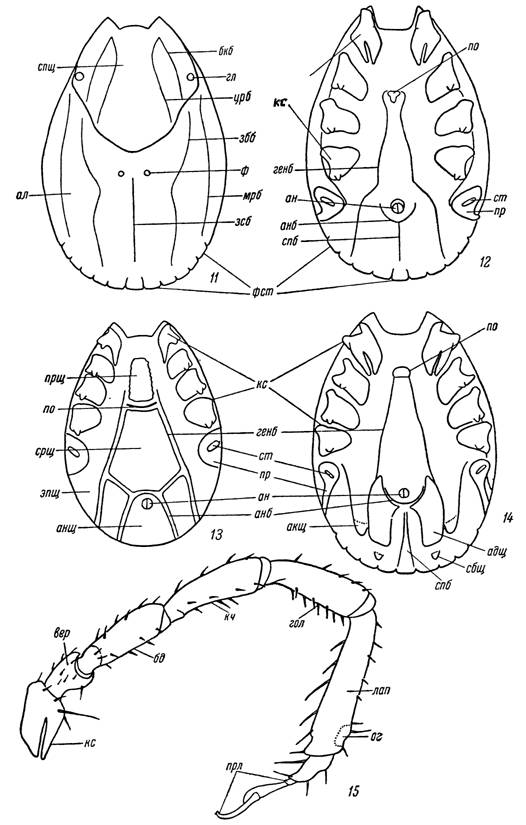
Рис. 11 — 15. Схема внешнего строения иксодовых клещей.
11 — самка Hyalomma asiaticum со спинной и 12 — с брюшной стороны; 13 — самец Ixodes ricinus; 14 — самец Hyalomma asiaticum с брюшной стороны; 15 — передняя нога самки Н. asiaticum.
спщ — спинной щиток; гл — глаза; ал — аллоскутум; црб — цервикальная и бкб — боковая борозды спинного щитка; зсб — заднесрединная, збб ~ заднебоковая и мрб — маргинальная борозды аллоскутума; ф — фовеа; по — половое отверстие; ап — анус; кс — коксы; пр — перитрема; cm — стигма; генб — генитальная борозда; апб — анальная и спб — срединная постанальная борозды; фст — фестон; щитки: прщ — прегенитальный, эпщ — эпимеральный, срщ — срединный, анщ — анальный, адщ — аданальный, акщ — акцессорный и сбщ — субанальный; вер — вертлуг; бд — бедро; кч — коленный членик; гол — голень; лап — лапка; прл — предлапка; ог — орган
Галлера.
вых видов Ixodinae гипостом в ряде случаев полностью лишен зубцов (Ixodes vespertilionis, Ceratixodes putus). Верхняя половина хоботка образована хелицеральными футлярами — парными трубкообразными выростами основания головки, которые окружают снаружи хе лидеры.
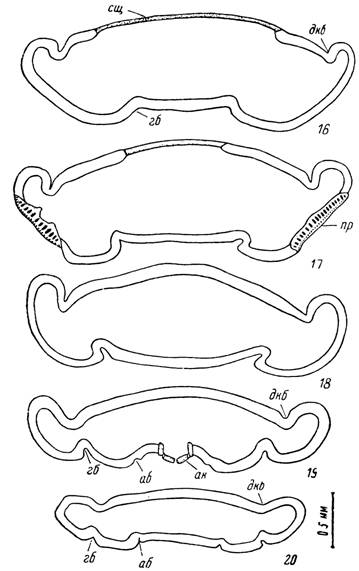
Рис. 16—20. Поперечные срезы через голодную самку Ixodes ricinus.
16 — на уровне спинного щитка, 17 — на уровне перитрем, 18 — позади перитрем, 19 — на уровне ануса, 20 — позади ануса.
Поверхность хелицеральных футляров также несет многочисленные мелкие зубцы.
Хелицеры иксодоидных клещей двучлениковые и полностью утратили свойственную паукообразным клешневидную форму (рис.
26). Проксимальный членик стержневидной формы с бульбо- образно расширенной базальной частью. В ней заключены мощные пучки мышц-антого- нистов, посылающих два сухожилия к подвижному пальцу хелицеры. У видов рода Ixodes сухожилия проходят в трубчатых каналах в стенках стволов хелицер, а у остальных уродов ^свободно лежат в их полости.сщ — спинной щиток; пр — перитрема; ап — анальное кольцо и створки анального клапана; дкб — дорсальная краевая борозда; г б — генитальная борозда; аб — анальная борозда.
Подвижный палец представлен треугольной, сильно склеротизованной пластинкой (внутренний членик), на внешней поверхности которого причленяется наружный членик. По своему наружному краю оба членика несут мощные режущие зубцы. К внутреннему членику с его внутренней поверхности неподвижно прикреплен зазубренный дорсальный отросток, а от ею сочленения со стволом отходит утолщенная в дорсальной и латеральной частях мембрановидная
складка-плащ. Она целиком окружает подвижный палец, исключая режущие концы зубцов (рис. 26).
На дистальных концах наружного и внутреннего члеников, а также на дорсальной поверхности последнего под плащом у Hyalomma asiaticum находятся три сравнительно крупные поры, сообщающиеся с внутренней полостью стволов. Подобные отверстия были найдены и у других видов иксодовых и аргасовых клещей (Robinson a. Davidson, 1913—1914; Gregson, 1960), но их происхождение и функции остаются неясными. Д. Грегсон (Gregson, 1960) предполагает, что они служат для выхода полостной жидкости, обеспечивающей расправление или спадение складок плаща. Возможно, что они представляют рудиментарные отверстия протоков хелицеральных желез, свойственных многим паукообразным.В задней трети стволов на уровне глоточного отверстия значительные участки дорсо-латеральной поверхности стенок представлены сильно складчатой двойной мембраной (рис. 192). Физиологическое значение подобных продольных окон в сильно склеротизованных трубках стволов также остается неясным.
От передне-боковых сторон основания гнатосомы отходят 4-членико- вые пальпы, гомологичные педипальпам остальных паукообразных. У аргазид членики пальп сохраняют цилиндрическую форму и подвижные сочленения (рис. 197).
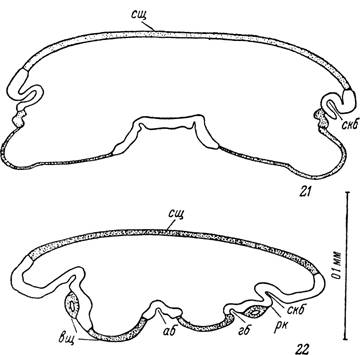
Рис. 21, 22. Поперечные срезы через голодного самца Нуаіотта рІитЬеит.
У иксодовых клещей наиболее крупные II и III членики неподвижно срастаются между собой. IV членик значительно меньше остальных и лежит в углублении на вентральной стороне III членика (рис.
24). Пальпы не принимают непосредственного участия в прикреплении к покровам хозяина и несут сенсорные функции.
На уровне: 21 — второй пары ног, 22 — позади анального отверстия.
сщ — спинной щиток; вщ — вентральные щитки; скб — суп- ракоксальная борозда; гб — генитальная борозда; аб — анальная борозда; рк — растяжимая кутикула вентро- латеральных участков идиосомы.
У голодных иксодовых клещей гнатосома занимает терминальное положение, а у напитавшихся особей, исключая самцов, в связи с неравномерным ростом кутикулы на вентральной и дорсальной сторонах идиосомы она смещается на брюшную сторону и занимает передневентральное положение.
Главная ось гнатосомы при этом перестает совпадать с таковой идиосомы. У имаго и нимф большинства видов аргасовых клещей гнатосома смещена вентрально и занимает передневентральное положение. Первичное терминальное положение сохраняется у личинок большинства видов, а в ряде случаев и у нимф (Alveonasus lahorensis и Otobius megnini).
По эмбриологическим данным Ю. Вагнера (1894) и А. Эшлимана (Aeschlimann, 1958) основная часть гнатосомы построена за счет разрастания кокс пальп. Дорсально они соединяются над хелицерами с остатком первичной головной лопасти и образуют верхнюю стенку головки — тектум. Сливаясь вентрально между собой, они завершают формирование кутикулярного кольца. Медиально-дорсальные выросты кокс, кроме того, как и у других паукообразных, соединяются с эпистомом и формируют так называемую субхелицеральную пластинку. От дистального конца эпистома отходит верхняя губа у аргазид, видоизмененная в сильно вытянутый отросток, называемый стилетом (Sen, 1935), или язычковидным отростком (Bertram, 1939). Вентрально вытянутые передние отростки кокс сливаются по средней линии и образуют гипостом. Наконец, в построении вентральной поверхности основания гнатосомы несомненно принимают участие стерниты хелицерального и пальпального сегментов.
— И —Функциональная анатомия гнатосомы рассматривается в связи с механизмом питания клещей (см. стр. 127—135).
Конечности. Нога клеща состоит из следующих 6 члеников: кокса, вертлуг, бедро, коленный членик, голень и лапка (рис. 15). Коксы срастаются со стенками идиосомы, но благодаря сохранению мембрановидных пограничных участков они сохраняют ограниченную подвижность (подтягивание и отхождение от вентральной поверхности тела). Все остальные членики обладают высокоподвижными сочленениями. Лапка клещей заканчивается парой коготков с перепончатой присоской, и эту структуру называют предлапкой.
Еще по теме ВНЕШНЕЕ СТРОЕНИЕ:
- Тема 11. Кожа. Строение кожи
- 1. Анатомическое строение почек.
- Внешнее строение
- ВНЕШНЕЕ СТРОЕНИЕ
- Факторы, влияющие на построение личного профессионального плана оптанта; типы индивидуальных ситуаций выбора профессии
- 2.4 Построение диагностической гипотезы
- Характер, его строение и формирование. Характер и личность
- 5. Обучение внутри организации или во внешних организациях.
- Характеристика и происхождение долгосрочной памяти на однократное восприятие внешнего мира.
- Строение и функции системы мозгового кровообращения в норме
- ВНУТРЕННЕЕ СТРОЕНИЕ СИМПАТИЧЕСКОЙ СИСТЕМЫ.
- Гистологическое строение.
- ГЛАВА 1 СТРОЕНИЕ ОРГАНА ЗРЕНИЯ И МЕЖДУНАРОДНАЯ АНАТОМИЧЕСКАЯ НОМЕНКЛАТУРА
- Строение данной группы рубцов
- ПОСТРОЕНИЕ ФУНКЦИОНАЛЬНО - СТРУКТУРНОЙ ОРГАНИЗАЦИИ (ФСО) ИАСППР И АЛГОРИТМОВ ЕЕ ФУНКЦИОНИРОВАНИЯ. ЭКСПЕРИМЕНТ ОБРАБОТКИ ВИДЕОКАДРОВ УЗИ.
- Построение базовой модели мультиагентной интеллектуальной системы для прогноза риска инсультов