Внешнее строение
Внешнее строение иксодовых клещей достаточно подробно описано в обзорах Балашов, 1967; Атлас..., 1979; Таежный клещ..., 1985; Sonenshine, 1991, 1993). Ик- ;оловые клещи являются наиболее фупными формами среди Асагіпа.
Напитавшие- :я самки наиболее крупных видов из родов Нуаіотта и АтЫуотта достигают в ллину 20—30 мм. У более мелких видов из родов Ixodes, Haemaphysalis, Boophilus, Rhipicephalus длина тела напитавшихся самок не превышает 5—15 мм. Размеры те- ~а голодных клещей значительно меньше, особенно у неполовозрелых фаз. Например. у голодных самок /. persulcatus длина тела составляет 3—4 мм, у самцов — 2—3, > нимф — 1.2—1.7 и у личинок — 0.7—0.9 мм. У Н. asiaticum (рис. 1) длина тела голодных самцов и самок составляет 6.5—8 мм, нимф — 1.3—1.6, личинок — I “4—0.79 мм.У голодных клещей тело сильно уплощено дорсовентрально и латеральные стороны развиты слабо. За время питания наряду с общим значительным увеличением размеров (у самок Я. asiaticum длина тела достигает 16.0—18.0 мм, нимф — - 9—6.1, личинок — 1.2—1.4 мм) происходит расширение туловища в дорсальном направлении, так что на поперечном разрезе оно становится овальным или круглым -абл. I[1]).
Помимо размеров тела личиночная, нимфальная и взрослая фазы клещей существенно отличаются некоторыми особенностями внешнего строения. У взрослой газы резко выражен половой диморфизм, проявляющийся в сохранении у самцов тольшей склеротизации туловища (размеры и число щитков вентральной и дор- сільной сторон), в строении гнатосомы и генитальных отверстий. Личинки и -тимфы по своему внешнему облику больше сходны с самками, но отличаются от ізтослых по строению гнатосомы, сенсорного аппарата, хетотаксии и отсутствию половых отверстий.
Тело клеща образовано из двух отделов: головки, или гнатосомы, и несегмен- ~*рованного туловища, или идиосомы. Задний край идиосомы, исключая виды годов Ixodes и Haemaphysalis, частично прорезан 9—11 краевыми фестонами.
Гнатосома своим основанием телескопически вдвинута в идиосому и подвижно :очленена с ней с помощью камеростомальной складки из эластичной кутикулы. >іа способна отгибаться вниз. У голодных особей гнатосома направлена вперед. Лс мере насыщения самок, нимф и личинок гнатосома смещается вентрально в :іізи с неравномерным ростом и растяжением покровов вентральной и дорсальной гторон идиосомы. У питающихся самцов подобного вентрального изгибания гна- -:сомы не происходит.У самок в дорсальной половине камеростомальной складки спрятано щелевид- ~:е отверстие протока органа Женэ. Последний представляет собой железу, секрет второй обволакивает откладываемые яйца и образует на их поверхности водоне- -рсницаемый слой. При откладке яиц напитавшимися самками мешковидная дис-
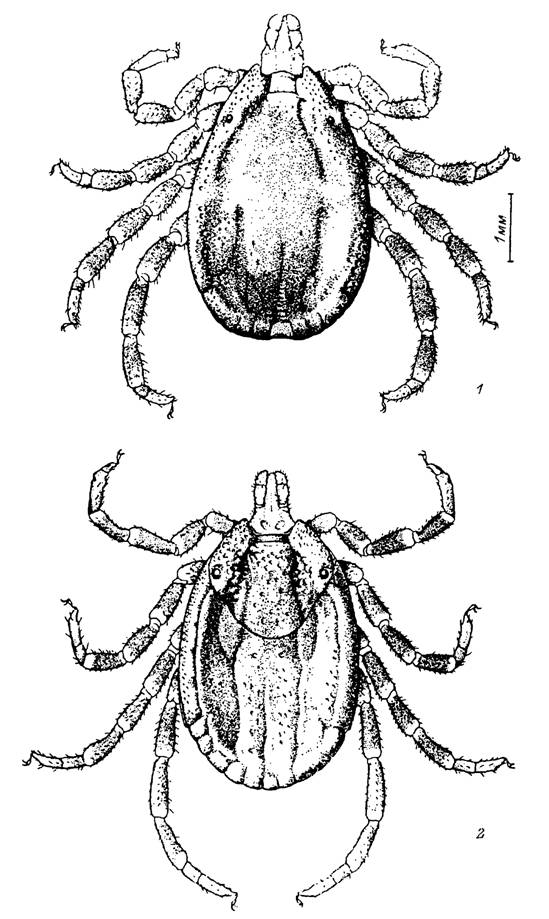
Рис. 1. Внешний вид сверху клеща Hyalomma asiaticum. 1 — самец; 2 — самка.
тальная часть органа Женэ выворачивается наружу и на нее поочередно попадают откладываемые яйца (Балашов, 1967).
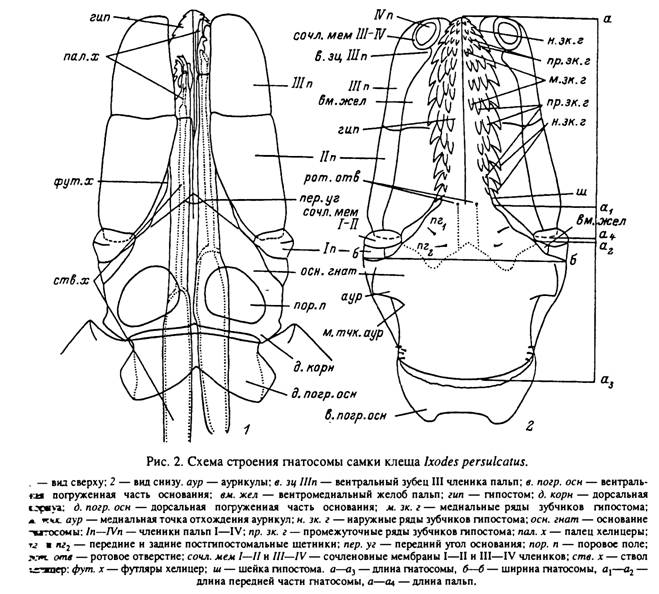
Головка состоит из базального склеротизованного кольца — основания головки. к которому спереди прикреплены вытянутые дистально ротовые части, образующие хоботок, или ротовой конус (рис. 2). По бокам от основания хоботка ттходят 4-члениковые пальпы.
Хоботок. Нижняя половина хоботка образована вытянутым вперед выростом основания гнатосомы — гипостомом. Последний слегка уплощается по направлению к дистальному концу и может быть заостренным (самка и нимфа /. persulcatus\ табл. II, 1) зли ложковидно расширенным (Я. asiaticum\ табл. VII, 7). Вентральная поверхность выпуклая и вооружена параллельными рядами направленных назад зубцов. На дистальном конце зубцы более мелкие и многочисленные. Они теряют правильное расположение и образуют так называемую корону (табл. II, 5; VII, 2). При больших ♦^сличениях видно, что зубчики короны изогнуты назад и заострены на концах. У :снования гипостома по средней линии расположена пара пор (вероятно, отверстия т*?токов каких-то желез), а по сторонам от них — пары симметричных конусовидных постгипостомальных хет, по-видимому хеморецепторной природы.
Дорсальная половина хоботка образована трубчатыми наружными хелицераль- -ісми футлярами, представляющими собой передний вырост крыши основания гна- -геомы (рис.
3). Базально внутренние стенки правого и левого футляров сливается. а дистально они разделены и лишь тесно прилегают друг к другу. Наружная дорсальная поверхность футляров покрыта многочисленными, направленными назадогн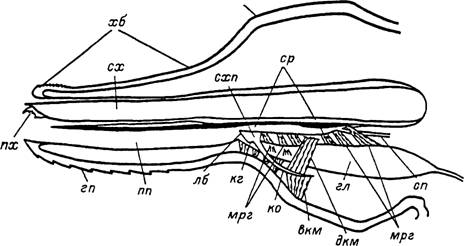
Рис. 3. Продольный разрез через гнатосому иксодового клеща (схема).
вкм — вентральные мышцы крыловидных отростков; гл — глотка; гп — гнпостом; дкм — дорсальные мышцы крыловидных отростков; кг — клиновидный гребень; ко — крыловидный отросток; лб — лабрум; мрг — мышцы-расширители глотки; огн — основание гнатосомы; пп — предротовая полость; пх — палец хелицеры; сп — слюнной проток; ср — слюнной резервуар; сх — ствол хелицеры; схп — субхелнцеральная пластинка; хб — хоботок.и налегающими друг на друга, наподобие черепицы, мелкими кутикулярными зубчиками (табл. V, 7).
Внутри футляров заключены длинные стержневидные хелицеры, состоящие из длинного ствола и пальца хелицеры. Ствол хелицеры образован вытянутым проксимальным члеником с расширенным базальным концом. Внутри последнего заключена пара мышц-антагонистов, связанных сухожилиями с основанием подвижного пальца. К основанию подвижного пальца латерально причленен неподвижный палец. Подвижный палец оканчивается 1 или 2 зубцами, а неподвижный палец также несет ряд острых зубцов (табл. II, 4\ XXIX, 5). Число и форма зубцов хелицер различаются у разных родов иксодид. С дорсальной стороны пальцы хелицеры окружает мембрановидный вырост (плащ), который представляет собой складку кутикулы основания подвижного пальца. На дорсальной поверхности дистального конца подвижного пальца открывается пора, из которой у живых особей выделяются небольшие капельки жидкости.
Стволы хелицер способны к перемещению в передне-заднем направлении внутри своих футляров. В заднем положении подвижный палец почти полностью убран внутрь футляра. В переднем положении подвижные пальцы выдвигаются из футляров и располагаются по сторонам от гипостома (табл. V, 5; VII, 3, 4).
Подвижные пальцы хелицер могут совершать режущие движения в медиолатеральной плоскости, благодаря чему происходит прорезание кожи животных-прокормителей.Между дорсальной поверхностью гипостома и„ вентральной хелицеральных футляров образуется трубчатая предротовая полость*. В заднем нижнем конце полости находится истинное анатомическое ротовое отверстие, тогда как функциональный рот занимает более переднее положение и лежит несколько позади дистального конца гипостома (рис. 3). На поперечном сечении предротовая полость имеет форму сильно сплющенного треугольника, причем в передней части хоботка футляры хелицер и гипостом лежат свободно и целостность ротовой полости обеспечивается только благодаря тесному прилеганию стенок этих органов друг к другу (табл. XV, 7). Далее назад происходит слияние боковых стенок гипостома с наружными хелицеральными футлярами, а также внутренних стенок последних между собой, так что предротовая полость приобретает морфологическую целостность. Дорсальная поверхность гипостома, образующая пол предротовой полости, сильно вогнута и формирует гипостомальный желоб, поверхность которого образована тонкой кутикулярной мембраной толщиной до 1 мкм (табл. XV, 2). Эта мембрана лишь латерально прикреплена к толстым склеротизованным стенкам гипостома, тогда как на большей части дорсальной поверхности гипостома между твердой кѵгикулой и поверхностной мембраной существует свободное пространство. В него поступает полостная жидкость, и, вероятно, в зависимости от степени заполнения подмембранного пространства мембрана может подниматься или опускаться и тем самым регулировать объем предротовой полости. Во время питания хоботок погружается в кожу хозяина так, что клещ получает возможность отсасывать жидкую пищу (кровь, лимфу и продукты лизиса тканей) из ее внутренних слоев, богатых кровеносными сосудами и расположенных много глубже анатомического рта.
В проксимальной части хоботка гипостомальный желоб углубляется, а гипо- стомальная мембрана смыкается над ним с дорсальной стороны, формируя таким образом рот, который ведет непосредственно в глотку и имеет треугольное сечение.
Над ротовым отверстием начинается субхелицеральная пластинка — главная опорная скелетная структура для хелицер и глотки.Часть предротовой полости, продолжающуюся назад над дорсальной поверхностью субхелицеральной пластинки, называют слюнным резервуаром (табл. XV, 3). Он сильно уплощен дорсовентрально, и в его заднебоковые углы открываются главные выводные протоки парных слюнных желез. Последние сначала проходят в толще субхелицеральной пластинки, а затем отделяются от нее в вентральном направлении. Потолок слюнного резервуара образован вентральной стенкой наружных хелицеральных футляров, которая в месте впадения слюнных протоков сливается с субхелицеральной пластинкой. В своей передней части субхелицеральная пластинка соединяется вентролатеральными выростами со стенками основания гнатосомы, образуя так называемый межкоксальный мост. В задней части субхелицеральная пластинка сильно утолщается, теряет связь со стенками гнатосомы и обеспечивает прикрепление на своей вентральной поверхности мышц-расширителей глотки.
Пальпы. Вторую пару придатков гнатосомы иксодовых клещей в связи с их преимущественно сенсорной функцией называют пальпами, хотя они и гомологичны ногочелюстям, или педипальпам, других групп хелицеровых. 4-члениковые пальпы отходят от переднебоковых углов гнатосомы (рис. 2). У большинства ик- содид они вытянуты дистально, уплощены дорсовентрально и благодаря желобообразной вогнутости вентромедиальных поверхностей II и III члеников в сложенном состоянии частично (Ixodes; табл. II, 7; III, 7) или полностью (Dermacentor, Rhipicephalus; табл. VI, 7, 3) прикрывают хоботок. Первый базальный членик небольших размеров подвижно сочленен с основанием гнатосомы и со II члеником. У 7. игіае пальпы почти цилиндрические и свободно лежат по сторонам от гипостома (табл. V). При сложенных пальпах хоботок имеет клиновидно заостренную (Ixodes; табл. II, 7) или тупо обрубленную (Dermacentor; табл. VI, 7) форму. Во время питания пальпы отгибаются почти под прямым (Ixodes, Hyalomma) или косым (Boophilus, Dermacentor) углом к продольной оси хоботка.
II и III членики пальп наиболее крупные, вытянуты дистально и слегка сплющены дорсовентрально.
Они неподвижно соединены между собой и с первым члеником. IV пальпальный членик (табл. VI, 2, 4\ VII, 7) цилиндрической формы, значительно меньше остальных. Он смещен на вентральную сторону III членика, находится в небольшом углублении и сохранил подвижность сочленения. На его вершине находится группа из 10 конусовидных волосков с закругленными вершинами (табл. XXIX, 2), служащих хеморецепторами. На своей наружной поверхности пальпы несут механорецепторные хеты, число, местоположение и форма которых постоянны для каждой из фаз развития; хеты варьируют в зависимости от систематической принадлежности клеща и могут быть важными диагностическими признаками.Строгая классификация пальпальных хет по их форме затруднительна из-за их исключительного морфологического разнообразия. По форме различают две группы пальпальных хет. К первой группе относятся волосковидные, или игольчатые,хеты с гладкими стенками (табл. III, 2). Они могут быть прямыми или слабо изогнутыми, почти одной толщины по всей своей длине или же сильно суживающимися от основания к вершине. Вторая группа включает сложные хеты с различными шипами или зубчиками на поверхности (табл. IV; VII, 1, 3, 4). Зубчики могут быть различной величины — от небольших поверхностных насечек до крупных шипов. Они располагаются по всей длине хеты или же ограничены ее дистальной зоной. Сложные хеты могут быть прямыми или изогнутыми, короткими или очень длинными.
Основание гнатосомы. На дорсальной поверхности основания гнатосомы у самок располагаются правильные парные скопления отверстий в кутикуле, называемые поровыми полями (табл. VIII, 7). Между этими отверстиями располагаются 3—4 небольшие палочковидные хеты. Отверстия на поровых полях являются окончаниями кутикулярных выводных протоков расположенных под ними железистых образований. Предполагают, что секрет этих желез служит смазкой при выворачивании органа Женэ из камеростомальной складки во время яйцекладки.
На заднелатеральных краях основания гнатосомы с вентральной и дорсальной сторон лежат группы невысоких конусовидных хет и обычных прямых или слегка изогнутых остроконечных трихоидных хет (табл. III, 2—4\ VII, 2, 3). Конусовидные хеты, по-видимому, являются хеморецепторными сенсиллами, а трихоидные — механорецепторными.
Спинной щиток. Переднюю половину дорсальной поверхности идиосомы (табл. II, 2), а у самцов всю поверхность (табл. III, 7) занимает спинной щиток, или скутум, образованный из твердой склеротизованной кутикулы. На переднем конце скутума имеется прямоугольная вырезка, по которой проходит сочленение гнатосомы с идиосомой. Передние углы скутума, расположенные уже на уровне гнатосомы, называют скапулами. Поверхность скутума самок, нимф и личинок покрыта характерным ромбическим рисунком и несет несколько борозд, из которых наиболее развиты цервикальные. На срединно-латеральных углах скутума, кроме видов рода Ixodes, расположена пара крупных орбитальных глаз (табл. XXVI, 4)у а на его поверхности обнаружены несколько типов хет и отверстия протоков дермальных желез или погруженных в толщу кутикулы сенсилл. Среди ску- тальных хет мы различаем несколько парных механорецепторных сенсилл с заостренными концами, встречающихся главным образом в средней трети скутума. Очень мелкие конусовидные, палочковидные и тонкие остроконечные хеты, расположенные на переднебоковых зонах скутума, вероятно, несут хеморецепторные функции. Во внутренних углах скапул расположены крупные отверстия погруженных сенсилл.
Поверхность растяжимой кутикулы. Большая часть идиосомы личинок, нимф и самок покрыта эластичной растяжимой кутикулой, образующей у голодных особей правильную систему параллельных микроскладок (табл. IX, 7; X, 7, 2). У напитавшихся клещей эти складки расправляются, а на их границах остаются лишь неглубокие бороздки (табл. IX, 2). Кроме того, по поверхности идиосомы голодных клещей проходит несколько более глубоких борозд, каждая из которых имеет свое наименование. Во время питания борозды идиосомы частично или полностью расправляются. Растяжимую часть дорсальной поверхности идиосомы позади скутума называют аллоскутумом.
У самцов аллоскутум отсутствует, и спинной щиток, называемый конскутумом, занимает всю дорсальную поверхность тела. Кроме краевой зоны растяжимая кутикула занимает у них межкоксальную область и узкие пространства между вентральными щитками (табл. IX, 5). В ней отсутствует правильная система микроскладок, а имеются лишь более глубокие и редкие борозды. Соответственно и способность к растяжению у описываемого типа кутикулы ограничена.
Фовеа. Сразу позади спинного щитка у нимф и самок на аллоскутуме и у самцов на конскутуме симметрично лежат два округлых участка из ямчато-мор- щинистой кутикулы — фовеа (табл. VIII, 2). Внутри и по краям фовеа находятся узкие щелевидные отверстия. Предполагается, что этими отверстиями открываются протоки расположенных под кутикулой желез, которые у самок вырабатывают половые феромоны (Sonenshine et al., 1983).
Генитальные отверстия. Вентральная поверхность идиосомы занята в передней трети сросшимися с ней массивными первыми члениками ног — коксами (табл. XIII, 5 4). По средней линии на уровне III кокс у половозрелых особей располагается гоперечное половое отверстие (табл. VIII, 5), прикрытое у самцов снаружи тонкой :клеротизованной пластинкой — апроном (табл. VIII, 4\ XI, 2).
Анальное отверстие. В задней трети тела лежит анальное отверстие (табл. IX,
XL 5). Оно окружено широким кольцом околоанальной складки из эластичной, ;гбранной в систему папиллообразных складок кутикулы. Внутри него заключен анальный клапан из твердого кутикулярного кольца и подвижно сочлененных с -им створок. Последние способны замыкаться и размыкаться. При размыкании створок между ними выворачивается часть задней кишки. На створках симмет- гично расположены у личинок 1 пара, у нимф и имаго несколько пар тонких, гтсчггепенно заостряющихся к концам волосков.
Хетом. Большая часть хет на поверхности идиосомы простые, по-видимому ;ыполняющие механорецепторные функции. Подобные хеты имеют типичное ча- п^видное сочленение. У Amblyomminae (табл. IX, 7) хеты гладкостенные, и толщина их может быть одинаковой по всей ее длине или же они слегка сужаются і дистальному концу. Конец хеты закругленный. У Ixodinae хеты вооружены зубчиками и чешуйками, форма и размеры которых отличаются у разных видов табл. X, 5, б). Кроме описанного выше типа по краю идиосомы и в зоне фестонов «стречаются более редкие, так называемые погруженные хеты. Они углублены в -длшу кутикулы, меньше размером и сильнее сужаются к своему дистальному к:нцу.
Форма, число и взаимное расположение хет идиосомы у личинок клещей постоянны для каждого вида (ортотрихия) и являются важными таксономическими дензнаками. У нимф, особенно у имаго, количество хет увеличивается во много гдз по сравнению с личинками, а постоянство их расположения в значительной степени утрачивается (неотрихия).
Кутикулярные поры. Кроме хет на поверхности идиосомы встречается боль- _»? количество отверстий, различающихся размерами и формой. Большая часть ~\. по-видимому, является открывающимися наружу протоками дермальных желез. Некоторые из этих отверстий связаны с сенсорными образованиями в толще или -_?д поверхностью кутикулы.
Наиболее характерны почти округлые крупные поры, окруженные снаружи ва- -йком. Внутри этих пор видна слегка выступающая наружу дужка. Подобные :~зерстия, особенно многочисленные в области фестонов (табл. IX, 5), встречается по всей идиосоме. Медиально от каждой коксы симметрично располагается ~о отверстию подобного типа (табл. XIII, 5, 4). Этот тип отверстий, очевидно, стелставляет собой какие-то погруженные сенсиллы, широко распространенные у членистоногих. Крупные кутикулярные отверстия второго типа также имеют ок- гуглую или овальную форму, но часто суживаются вдоль одной из осей, лишены -дружного ободка и внутри их не видна дуговидная структура (табл. XI, 7). Полными отверстиями, вероятно, открываются наружу дермальные железы II типа, пдиболее многочисленны мелкие отверстия стреловидной или треугольной формы, где полагающиеся правильными рядами в растяжимых участках идиосомы (табл. IX,
Множество мелких щелевидных отверстий располагается вдоль границы кокс : кутикулой идиосомы (Атлас..., 1979; Walker et al., 1996а, 1996b).
Стигмальные пластинки. Вентролатерально сразу позади кокс IV пары ног у -ѵмф и имаго расположены стигмальные пластинки, или перитремы, на которых -крываются дыхательные отверстия, или стигмы. Форма и размеры стигмальных -.-астинок у разных видов варьируют от округлой или овальной до вытянутой с гоосокаудальным отростком (табл. XI, 4\ XII, 7). Перитрема — сложное образование из твердой склеротизованной кутикулы. Наружный край ее гладкий и об- газ>ет так называемую хитиновую каемку. Ближе к переднему краю перитремы -ілодится щелевидное дыхательное отверстие, ведущее в обширную атриальную полость, которая в свою очередь сообщается с главными стволами трахейной системы. Снаружи дыхательное отверстие частично прикрыто гладким кутикуляр- ным бугорком — макулой. Вокруг дыхательного отверстия располагается узкая зона гладкой кутикулы, а за ней к краям поверхность перитремы пронизана множеством круглых отверстий (табл. XI, 5, 6). Эти отверстия сложной системой внугрикутикулярных каналов сообщаются с хеморецепторными сенсиллами, открываясь в сотовидную массу бокаловидных полостей в толще пластинки (табл. XII, 2). Дыхальца клещей обладают эффективным замыкательным аппаратом, обеспечивающим чередование периодов активного газообмена с длительными паузами (Fielden et al., 1994).
Ноги. К вентральной стороне идиосомы причленяются 4, а у личинок 3 пары ходильных ног. Каждая нога образована 6 члениками (кокса, вертлуг, бедро, коленный членик, голень и лапка), соединенных между собой высокоподвижными сочленениями из эластичной кутикулы. Лишь коксы полностью срослись с кутикулой идиосомы.
Зубцевидные выросты на коксах и вертлугах ног вместе с кутикулярными выростами основания гнатосомы обеспечивают фиксацию клеща на теле хозяина (дабл. XIII, 2—4). Это достигается зажиманием волос при подгибании гнатосомы и ограниченных смещениях выростов кокс. У некоторых видов родов Ixodes и Haemaphysalis волосы зажимаются между передними выступами кокс пары ног и выростами членика пальп (Балашов, 1989а).
Членики ног построены из твердой, почти полностью склеротизованной кути- к\лы. от поверхности которой отходят многочисленные остроконечные трихоидные хеты обычного строения, большая часть которых выполняет тактильные функции. Кроме них на лапках I пары ног обнаружены сложно устроенные сенсорные образования — органы Галлера (табл. XXVI, 7; XXX, 2) и одиночные волоско- видные хеморецепторные сенсиллы.
Лапка заканчивается двумя сильно развитыми коготками и расположенной между ними подушечкой (табл. XIII, 7), выполняющей функции присоски. Поверхность подушечки, спадаясь, собирается в складки эластичной кутикулы. Вероятно, с подушечкой связан и выводной проток находящейся внутри лапки тарзальной железы.
Покровы
Тело иксодовых клещей покрыто плотными покровами, выполняющими роль наружного скелета и благодаря высокой водонепроницаемости препятствующими испарению воды. Строение покровов иксодид подробно описано для Boophilus (Beadle, 1974), Я. asiaticum (Атлас..., 1979), I. persulcatus (Таежный клещ..., 1985) и Я. dromedarii (El Shoura, 1988а). У всех этих видов строение покровов было сходным. В состав покровов входит слой гиподермальных клеток (гиподерма) и выделяемая ими кутикула, которая имеет сложное строение* Внутренняя часть кутикулы, образующая основную ее массу и лежащая непосредственно над гиподермой, носит название прокутикулы.
Прокутикула построена из связанных между собой молекул протеинов и хитина. Биохимическими исследованиями установлено, что для прокутикулы иксодовых клещей характерны относительно низкое содержание хитина (около 4 %) и своеобразный аминокислотный состав белков. Эти особенности кутикулы, возможно, обусловливают исключительную способность некоторых участков покровов к растяжению. В зависимости от степени склеротизации и способности к растяжению различают три вида прокутикулы: эндокутикулу, мезокутикулу и экзокутикулу. Эндокутикула — несклеротизованная растяжимая прокутикула; экзокутикула — полностью задубленная; в мезокутикуле процессы склеротизации еще не завершены. В тех случаях, когда в состав кутикулы входят все три вида прокутикулы, эндокутикула располагается над слоем гиподермальных клеток, мезокутикула — над ней, а экзокутикула образует внешний, наиболее удаленный от гиподермы слой.
Из эндокутикулы построена растяжимая часть идиосомы недавно перелинявших клещей. В дальнейшем, на стадии послелиночного доразвития происходит частичная склеротизация прокутикулы в этой части покровов и превращение ее в ме- зокутикулу. Толщина эндо- и мезокутикулы голодных самок таежного клеща составляет соответственно 3—4 и 15—18 мкм, а у Я. asiaticum они вдвое толще. У нимф толщина прокутикулы меньше, чем у взрослых, а у личинок она тоньше, чем у нимф, и не превышает 2—8 мкм. На электронограммах мезокутикула (табл. XIV, 1—3) имеет тонкогранулярную структуру и обладает умеренной электронной плотностью. Она пронизана слегка извилистыми поровыми каналами диаметром 100—200 нм, в просвете которых иногда содержится гранулярный или фиброзный материал. Эндокутикула имеет выраженную ламеллярную структуру. Каждая ламелла состоит из тонких микрофибрилл, образующих сложный рисунок; толщина ламеллы 1.5—2 мкм. Поровые каналы этой части кутикулы имеют дугообразную форму.
Только из экзокутикулы образована кутикула твердых (нерастяжимых) покровов — спинного щитка, вентральных щитков самца, анального клапана, перитрем, гнатосомы, конечностей. Экзокутикула не окрашивается без специальной предварительной обработки и имеет на срезах янтарно-желтый оттенок. На электронограммах она выглядит гомогенной, но несколько более электронно-плотной по сравнению с мезокутикулой.
Поверхность кутикулы таежного клеща образует регулярные складки (табл. IX, 7; X, 1, 2) и, так же как у других наземных членистоногих, покрыта эпикутикулой, которая при исследовании в световом микроскопе выглядит однородной. В отличие от прокутикулы для эпикутикулы характерно отсутствие хитина. Толщина этого слоя, по данным световой микроскопии, составляет 1—4 мкм.
Строение эпикутикулы иксодид одинаково во всех частях покровов независимо от степени их склеротизации. По данным электронной микроскопии, она имеет сложное строение и состоит из четырех слоев, различающихся по толщине и электронной плотности (рис. 4). Самый внешний из слоев эпикутикулы, восковой, имеет толщину около 100—200 нм. Для него характерна умеренная электронная плотность, и иногда в нем выявляется горизонтальная слоистость. Лежащий на нижней границе воскового очень тонкий (2—4 нм) электронно-плотный слой, по- видимому, образован мономолекулярным слоем липидов. Следующий, кутикулино- вый, слой отличается от остальных частей эпикутикулы чрезвычайно высокой электронной плотностью, его толщина составляет 15—30 нм. Этот слой первым откладывается при образовании кутикулы во время линьки. Под кутикулиновым слоем располагается протеиновая эпикутикула, близкая по структуре и электронной плотности к прокутикуле, с которой граничит. Толщина этой части эпикутикулы 150—300 нм.
Гиподерма представлена одним слоем клеток. Клетки голодных особей имеют уплощенную форму, их высота составляет от 1—2 мкм у личинок до 5—10 мкм у взрослых. Ультраструктура гиподермальных клеток голодных особей (табл. XIV, 2) свидетельствует об отсутствии в них активных синтетических и секреторных процессов. В цитоплазме слабо развиты элементы гранулярного эндоплазматического ретикулума и содержится лишь незначительное количество свободных рибосом.
Строение покровов значительно изменяется в период многодневного питания, во время которого происходят рост и растяжение кутикулы. В течение большей части периода питания наблюдается увеличение толщины кутикулы, причем отмечен рост как эндокутикулы, так и мезокутикулы.
У самок таежного клеща толщина эндокутикулы увеличивается с 3—4 до 17— 30 мкм, а мезокутикулы — с 15—18 до 30—32 мкм. Одновременно отмечается и возрастание сухой массы кутикулы. В конце периода питания рост кутикулы
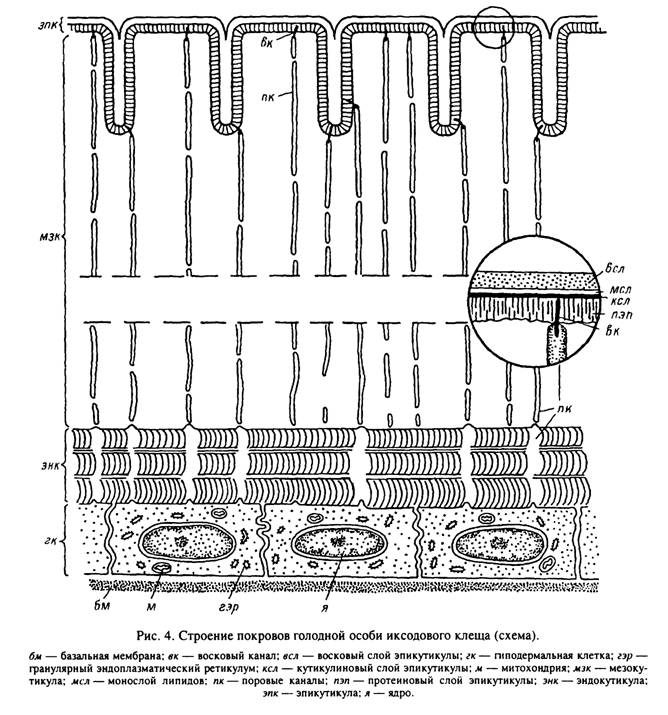
прекращается и происходит ее растяжение, при этом толщина мезокутикулы снижается до 10—12, а эндокутикулы — до 9—J0 мкм. О растяжении кутикулы свидетельствуют увеличение диаметра поровых каналов и изменения в ультраструктуре кутикулярных ламелл. Растяжение сопровождается расправлением борозд и регулярных складок на поверхности растяжимой кутикулы идиосомы.
Рост кутикулы обеспечивается секреторной активностью гиподермы, в строении которой в это время наблюдаются значительные изменения. Высота клеток увеличивается в 3—4 раза по сравнению с периодом голодания и достигает 30— 35 мкм, а форма клеток становится столбчатой. В конце периода питания толщина гиподермального слоя вновь несколько уменьшается, что связано с интенсивным растяжением покровов, происходящим в это время. С началом питания увеличиваются и размеры ядер и ядрышек. В гиподерм аль ном слое часто можно наблюдать делящиеся клетки. Цитоплазма становится базофильной, в ней выявляются РНК и щелочная фосфатаза. Повышается электронная плотность цитоплазмы, в ней появляются многочисленные цистерны гранулярного эндоплазматического ретикулума и комплексы Гольджи, а в апикальных частях клетки, граничащих с кутикулой (табл. XIV, 4), — секреторные пузырьки нескольких видов. На поверхности клеток, обращенной к кутикуле, возрастает количество микроворсинок, по ультраструктуре значительно отличающихся от микроворсинок голодных особей. Они, по-видимому, принимают участие в образовании и ориентации хитиново-протеиновых фибрилл прокутикулы.
Гистологические изменения, происходящие в период линьки, однотипны у всех иксодид. В отличие от других кровососов у иксодид процессы роста покровов и внутренних органов, связанные с питанием, переходят в процессы подготовки к линьке. Рост продолжается и в течение нескольких дней, которые отделяют момент отпадения напитавшегося клеща от начала линочных изменений. В этот период гиподермальные клетки продолжают активно секретировать. Вероятно, одновременно происходит изменение химического состава покровов и усиление их скле- ротизации, благодаря чему возрастают их твердость и хрупкость.
Перед началом линьки рост кутикулы прекращается. Далее происходит отделение кутикулы от гиподермы- (или аполизис), которое, с одной стороны, представляет следствие митотической активности гиподермы, а с другой — является результатом растворения нижних ламелл эндокутикулы под действием секрета гиподермы. Пространство между гиподермой и кутикулой, возникающее в результате их отделения друг от друга, носит название экзувиальной полости. Ее образование начинается с переднего конца тела. Экзувиальная полость заполнена линочной жидкостью. Содержимое экзувиальной полости в процессе формирования новой кутикулы частично всасывается гиподермальными клетками.
Откладка кутикулы следующей фазы развития начинается с послойного образования эпикутикулы. Первым откладывается кутикулиновый слой. Вслед за кути- кулиновым слоем откладывается протеиновая эпикутикула. В то же время начинается формирование кутикулярных складок. Первым шагом к образованию поверхностного рисунка является формирование на апикальной поверхности гиподермы, покрытой двумя слоями эпикутикулы, регулярных углублений. Микроворсинки в промежутках между ними значительно удлиняются. Над концами и вокруг микроворсинок происходит откладка материала прокутикулы. По мере образования прокутикулы внутри складок на месте микроворсинок формируются поровые каналы. В период откладки основной массы прокутикулы микроворсинки укорачиваются до своей обычной длины (около 0.5 мкм), а в образовании поровых каналов принимают участие цитоплазматические выросты на поверхности гиподермальных клеток. Формирование эпикутикулы завершается уже после сбрасывания линочной шкурки, когда на поверхности кутикулы появляется восковой слой. Откладка различных компонентов кутикулы сопровождается активной секреторной деятельностью гиподермы.
В начале периода линьки — во время откладки кутикули нового слоя и протеиновой эпикутикулы наблюдается скопление в базальных частях гиподермальных клеток больших количеств гликогена. Как показали электронно-микроскопические исследования, скопления гликогена занимают практически всю подъядерную зону клеток, не оставляя места для других видов включений и органоидов. К моменту сбрасывания старой кутикулы количество гликогена значительно уменьшается.
Активные синтетические процессы продолжаются в гиподермальных клетках и после окончания линьки. В период последи ночного доразвития происходит скле- ротизация покровов и на этом завершается формирование эпикутикулы.
Помимо обычных гиподермальных клеток в состав покровов иксодовых клещей входят специализированные железистые клетки, связанные с выводным протоком, открывающимся на поверхность кутикулы. Эти образования получили название дермальных желез. Дермальные железы встречаются по всей поверхности растяжимой кутикулы идиосомы, на спинном щитке и на основании гнатосомы. Они открываются на поверхность сложно устроенными кутикулярными протоками, которые у желез
Рис. 5. Строение дермальной железы I типа Ixodes persulcatus.
пж
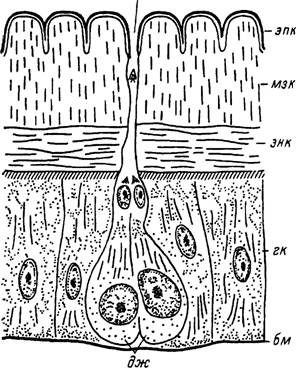
бм — базальная мембрана; гк — гнподермальная клетка; дж — секреторные клетки дермальной железы; мзк — мезо- кугнкула; пж — проток дермальной железы; энк — эндокутикула; эпк — эпнкугнкула.
I типа иногда называют копьевидными сенсиллами и у желез II типа — стреловидными.
У /. persulcatus обнаружены только дермальные железы I типа. Они состоят из двух железистых клеток и нескольких клеток, образующих проток (рис. 5). У голодных особей железистые клетки практически неотличимы от окружающих клеток гиподермы. Активизация их секреторной деятельности происходит с началом питания. В это время железистые клетки и их ядра увеличиваются в объеме. Наблюдается усиление базофи- лии цитоплазмы. В апикальной части клетки обычно появляется крупная вакуоль, которая, по данным электронной микроскопии, представляет обширную инвагинацию поверхностной мембраны железистой клетки, сообщающуюся с протоком железы. Поверхность инвагинации несет многочисленные микроворсинки. В ультраструктуре цитоплазмы также происходят значительные изменения: резко увеличивается количество элементов гранулярного эндоплазматического ретикулума, появляются развитые комплексы Гольджи и многочисленные секреторные вакуоли. Железистые клетки, очевидно, продолжают активно секретировать и у клещей, закончивших питание, однако с началом линочных изменений происходит их дегенерация, хотя ядра железистых клеток сохраняются вплоть до начала откладки эпикутикулы.
У Я. asiaticum (Атлас..., 1979), R. appendiculatus (Walker et al., 1996a, 1996b) и других амблиомин кроме описанных выше желез I типа имеются также более крупные железы II типа. Они также состоят из 2 железистых клеток, которые уже у голодных особей значительно крупнее гиподермальных (рис. 6). С началом кровососания размеры их быстро увеличиваются и достигают у напитавшихся самок 200—250 и у нимф 100 мкм. После отпадения напитавшихся особей размеры желез продолжают некоторое время увеличиваться и достигают 300—400 мкм при диаметре ядер 40—80 мкм. Апикальная часть каждой из железистых клеток заполняется секреторными вакуолями, содержимое которых изливается в обширную полость^ сообщающуюся с выводным протоком железы. Именно у напитавшихся особей секрет этих желез изливается на поверхность покровов в виде мелких капелек жидкости у отверстий их стреловидных протоков. Затем начинается их быстрая дегенерация, полость спадается и железа исчезает.
Функции дермальных желез как таежного клеща, так и других иксодовых клещей остаются неясными. Некоторые исследователи (Лагутенко, 1969) описывали их как железисто-чувствительные органы — копьевидные сенсиллы и приписывали секрету функцию смазки, предохраняющей чувствительные волоски. Однако дальнейшие исследования не подтвердили этого предположения. Дермальные железы образуют белковые и липоидные секреты, часть из которых участвует в феромонной коммуникации клещей (Walker et al., 1996b).
Специализированным типом дермальных желез являются так называемые фо- веальные железы, подробно исследованные у Я. dromedarii (Sonenshine et al.,
сен
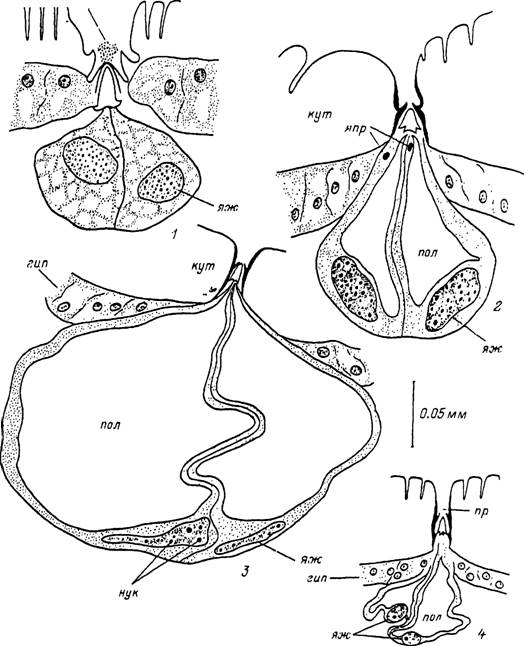
Рис. 6. Строение дермальной железы II типа Hyalomma asiaticum.
; — напитавшаяся нимфа на стадии формирования кутикулы; 2 — голодная самка; 3 — самка в середине стадии питання; л — напитавшаяся самка вскоре после отпадения, гип — гиподерма; кут — кутикула; нук — нуклеола; пол — внутренняя полость железы; пр — проток дермальной железы; сек — секрет; яж — ядра железистых клеток; япр — ядра клеток протока.
1983) . На поверхности фовеа открываются выводные протоки нескольких десятков подобных желез. Каждая железа включает клетки протока и 3 железистые клетки. У голодных самок фовеальные железы находятся в неактивном состоянии. С началом питания размеры железистых клеток увеличиваются в несколько раз и цитоплазма их заполняется секреторными включениями .рис. 7). Предполагают, что продуктом их секреции служит летучий половой феромон — 2,6-дихлорфенол, выделяемый через отверстия в кутикуле фовеа питающихся самок.
Особые дермальные железы обнаружены и в тарзальных члениках каждой из 4 пар ног Я. asiaticum. Они образованы из многих клеток однослойного эпителия, окружающих мешковидную центральную полость железы. Полость железы открывается наружу выстланным кутикулой протоком. Клетки стенок железы заполнены секреторными гранулами.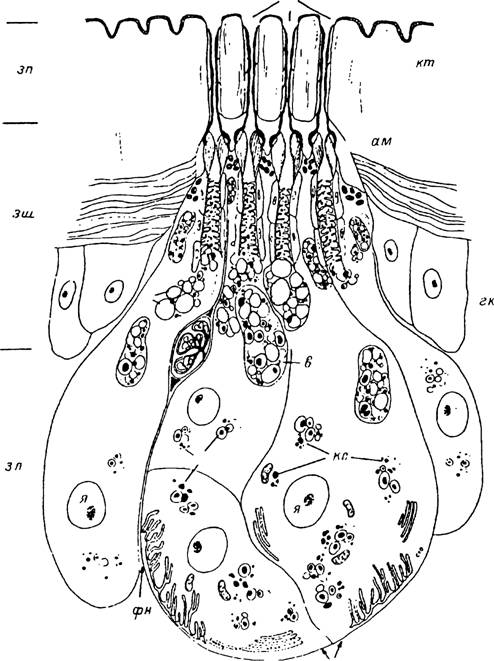
СК
Рис. 7. Фовеальная феромонная железа самки иксодового клеща (по: Sonenshine, 1985, с изменениями).
им — ампулы; в — вакуоли с секретом; гк — гнподермальные клетки; зп — зона протоков; зш — зона шейки; зл — зона секреторных лопастей; кт — кутикула; кс — капли секрета; п — прото* фовеальной железы; ск — секреторные клетки; фн — фовеальный нерв; фп — фовеальные поры:!— ядра секреторных клеток.
Специализированная группа дермальных желез расположена под поровыми полями. Протоки этих желез отрываются на поровых полях, а их секрет участвует в формировании водонепроницаемого покрытия откладываемых яиц (Kakuda et al., 1995а).
Еще по теме Внешнее строение:
- Г.Д. Жабоедов и др.. Офтальмология: учебник— K.: ВСИ “Медицина”,2011. — 448 с., 2011
- клинико-анатомические формы омертвения
- Раннее сексуальное развитие
- Современные представления о синдроме дисплазиисоединительной ткани
- Тема занятия. БОЛЕЗНИ СИСТЕМЫ КРОВЕТВОРЕНИЯ. АНЕМИИ
- 1.3.6 Chlamydia trachomatis
- Болезнь Меньера
- Н. Н. Зайко. Патологическая физиология, 2015
- ТИП ЧЛЕНИСТОНОГИХ (ARTHROPODA)
- КЛЕТОЧНЫЙ И ТКАНЕВОЙ УРОВНИ АДАПТАЦИЙ У ПОЙКИЛОТЕРМНЫХ ОРГАНИЗМОВ