ЦИТОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РАЗМНОЖЕНИЯУ САМЦОВ
Развитие мужской половой системы. У голодных личинок Нуаіотта asiaticum зачатки семенников представлены двумя компактными группами клеток, расположенными между центральной нервной массой и передней стенкой ректального мешка.
Они содержат по нескольку первичных спер- матогониев с крупными сферическими ядрами диаметром 5—6 мк и эпителиальных клеток с овальными ядрами в 4—5 мк. Снаружи зачатки окружены соединительнотканной оболочкой. Зачатки семепроводов и эктодермальные производные полового аппарата на этой стадии отсутствуют.С началом кровососания и во время линьки ядра эпителиальных клеток и первичных сперматогониев начинают делиться. Зачатки семенников удлиняются, причем их передние концы более узкие и состоят в основном из недифференцированных эпителиальных клеток. Они вытягиваются вперед и слепо заканчиваются у боковых стенок образовавшегося к этому времени гиподермального зачатка. Семепроводы на этой стадии представляют сплошные клеточные тяжи, просвет в них еще не развит. В семенниках внутренняя зона занята вторичными сперматогониями, а снаружи и между ними лежат более мелкие овальные ядра эпителиальных клеток.
Во время питания сразу же позади центральной нервной массы на вентральной стороне происходит утолщение гиподермального слоя. Во время линьки этот участок образует небольшое впячивание. Размеры его увеличиваются за счет многочисленных клеточных делений. Образовавшийся зачаток эктодермальных производных половой системы расширяется в заднедорсальном направлении и сужается в месте соединения с гиподермой стенки тела (рис. 424).
У голодных нимф рост и развитие половой системы приостанавливаются и возобновляются только с началом кровососания (рис. 425, 426). Размеры семенников быстро увеличиваются, они приобретают трубчатое строение и к концу линьки достигают размеров, свойственных голодным самцам. Внутри семенников в это время наблюдаются характерные изменения.
Начиная с задних концов вторичные сперматогонии окружаются эпителиальными клетками. В результате клеточных делений образуются сперматоцисты, заполненные первичными сперматоцитами и окруженные клеточной оболочкой. У закончивших линьку самцов задняя половина семенника содержит сперматоциты I порядка, готовые к большому росту, а передняя — вторичные сперматогонии.Длина и диаметр семепроводов значительно увеличиваются, а внутри появляется просвет. Передние слепые концы парных семепроводов лежат бок о бок и дорсально к выросту стенки половой полости. В середине линьки они сливаются в короткий непарный семепровод. Клетки на концах семепровода и выроста стенки половой полости расходятся и между ними устанавливается сообщение.
В эктодермальном половом зачатке голодной нимфы дифференцировка я^елезистых клеток еще не началась (рис. 425) и наблюдается лишь со второй половины линьки. Во время питания передний конец зачатка вытягивается и превращается в толстостенную дорсо-вентрально уплощенную трубку с узким щелевидным просветом (рис. 426). На заднем конце просвет слегка расширяется в небольшую половую полость. До середины линьки в зачатке семеизвергательного канала многочисленны митотические деления. Затем они прекращаются, ядра располагаются в один ряд и занимают середину клеток. Базальные половины клеток содержат крупные вакуоли, а апикальные сильно базофильны. На их поверхности формируется кутикулярная выстилка и к концу линьки высота клеток заметно уменьшается. Задняя половина гиподермального впячивания служит зачатком комплекса придаточных желез, который сильно разрастается на спинную сторону и вперед, образуя главную медиальную лопасть (рис. 427). Несколько позднее от нее в виде выростов формируются остальные лопасти придаточной железы. Клеточные границы в них на этой стадии еще не различимы, ядра лежат в несколько рядов и среди них очень много делящихся. В конце линьки деления прекращаются, а в вентро- латеральных лопастях появляется слой из базальных более крупных ядер.
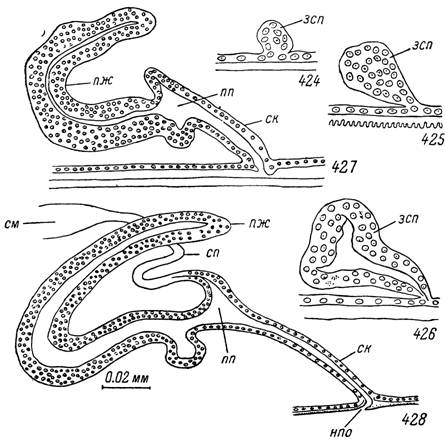
Рис.
424—428. Развитие мужской половой системы Нуаіотта asiaticum. Сагиттальный разрез.424 — напитавшаяся личинка; 425 — голодная нимфа; 426 — напитавшаяся нимфа; 427 — линяющая нимфа; 428 — голодный
самец.
зсп — зачаток семеизвергательного канала и придаточных желез; ск — семеизвергательный канал; пп — половая полость; пж — придаточная железа; сп — семепровод; см — семенник; нпо — наружное половое отверстие.
На этом развитие придаточных желез приостанавливается до начала питания. У Ixodes ricinus развитие мужской половой системы протекает сходным образом, но процессы сперматогенеза и дифференцировки железистой ткани придаточных желез заканчиваются на нимфальной фазе.
У аргасовых клещей зачаток семенника у личинок имеет вид дугообразно изогнутого валика, в котором еще невозможно обнаружить признаков половой дифференцировки (рис. 429). Он образован двумя типами клеток: более крупными первичными половыми клетками с пузыревидным ядром и более мелкими клетками будущих семенных фолликулов. Снаружи зачаток гонады покрыт одним слоем плоских эпителиальных клеток и неклеточной соединительнотканной оболочкой (рис. 432). У напитавшихся личинок размеры зачатка семенника быстро увеличиваются. Он вытягивается своими передними концами к формирующемуся в это же время мешковидному эктодермальному зачатку семеизвергательного канала
и придаточных желез. У нимф I возраста зачатки семепроводов соединяются с эктодермальной закладкой и дальнейшая дифференцировка половой системы растягивается на несколько нимфальных возрастов (рис. 430). Процессы роста и развития ограничены периодами линек и у голодных особей отсутствуют. Дифференцировка придаточных желез заканчивается во время последней линьки.
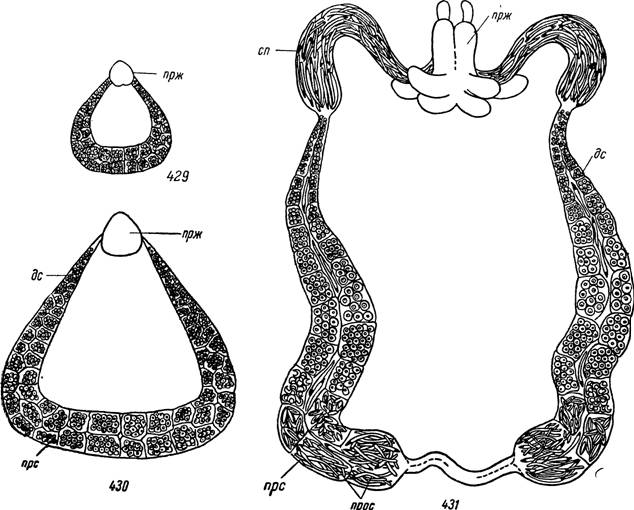
Рис. 429—431. Развитие мужской половой системы Argas persicus.
429 — личинка после питания, в проксимальных сросшихся отделах семенников спер- матоциты в фазе малого роста; 430 — нимфа I возраста, в проксимальных отделах семенников сперматоциты в фазе большого роста; 431 — самец 4—5-дневного возраста, в проксимальных отделах закончен сперматогенез, готовые проспермии выходят из семенников и скапливаются в семенных пузырьках.
прж — придаточные половые железы; сп — семенной пузырек; пре — проксимальный отдел семенника; дс — дистальный отдел семенника; прос — проспермии.
Сперматогенез.
Цитологические особенности сперматогенеза достаточно подробно описаны в литературе (Nordenskiold, 1920; Samson, 1909b; Warren, 1933; Tuzet a. Millot, 1937; Соколов, 1958; Wagner-Jevseenko, 1958; Горощенко, 1960, 1965; Rothschild, 1961; Reger, 1963). В связи с этим мы лишь кратко остановимся на некоторых особенностях этого процесса, необходимых для характеристики физиологического состояния самцов.Уже после питания личинок Hyalomma asiaticum первичные спермато- гонии в зачатке семенника начинают делиться и превращаются во вторичные. У нимф наряду с образованием вторичных сперматогониев многие из них продолжают развитие и проходят 5—6 последовательных делений. В результате внутри сперматоцист содержится по 32—64 сперматоцита
J порядка (рис. 435). Последующие стадии сперматогенеза наблщдаются только с началом имагинального питания.
Сперматогонии имеют небольшие сферические ядра диаметром 4—5 мк, окруженные узким кольцом цитоплазмы. Диаметр сперматоцитов I порядка до начала фазы большого роста составляет 5—8 мк, а после ее окончания — 26—30 мк. Размеры ядер увеличиваются до И—12 мк, а внутри них появляются нуклеолы в 3—4 мк (рис. 436). Большой рост сперматоцитов связан не только с очень значительным увеличением объема цитоплазмы и ядер, но и значительными изменениями во внутренней структуре последних. Ядро сперматоцита на этой стадии обладает большим сходством с зародышевым пузырьком ооцита. Почти сформировавшиеся к началу этой фазы биваленты разрыхляются и способность ядра давать положительную реакцию Фельгена временно исчезает. ДНК хромосом вновь начинает выявляться в самом конце большого роста, когда заканчивается образование бивалентов и исчезает ядрышко. В результате первого деления созревания, которое Ю. Л. Горощенко (1960) считает редукционным, образуются сперматоциты II порядка диаметром 18— 22 мк. Они проходят второе деление созревания (эквационное) и превращаются в сперматиды диаметром 15—17 мк (рис. 437).
Гистогенез спермиев начинается с вытягивания сперматид, достигающих длины 100—110 мк.
В результате этого процесса образуется трубка с терминально расположенным ядром и основной массой цитоплазмы. Из дна трубки в свою очередь формируется внутренний трубчатый вырост. Затем происходит вворачивание трубки проспермия и длина его уменьшается до 50—60 мк. Дальнейшее развитие проспермиев заканчивается уже в выводных путях женской половой системы, где происходит их деваги- нация и превращение в трубчатые спермиофоры длиной в 100—120 мк (рис. 151). Передний конец проспермиев расширен, а задний более узкий и несет небольшое ядро в виде компактного Фельген-положительного тельца (рис. 437). Собственно спермием у клещей служит ядро (Tuzet е. Mil- lot, 1937; Соколов, 1958), так как только оно участвует в оплодотворении. Огромная трубчатая часть спермия осуществляет лишь перенос и защиту .ядра и поэтоку ее называют спермиофорой.У аргасовых клещей процессы сперматогенеза в основных чертах сходны с иксодовыми (рис. 429—431 и 433). Сперматогонии у изученных видов (іOrnithodoros papillipes, О. moubata, Argas persicus, A. reflexus) проходят четыре последовательных деления, так что сперматоцисты содержат по 16 сперматоцитов I порядка. Во время большого роста величина последних увеличивается значительно больше, чем у Hyalomma asiaticum. Размеры ядер достигают 30—38 мк. Объем их увеличивается в 60 раз, а самой клетки — в 400 раз. Таким образом, аргасовые клещи обладают наиболее крупными «спермиями» среди членистоногих. Так, у Ornithodoros papillipes длина спермиофоров в половых путях самок составляет 850— 980 мк (Соколов, 1958).
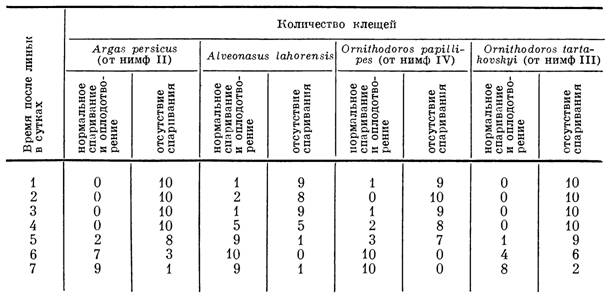
У аргасовых клещей (Балашов и Горощенко, 1960) созревание части сперматоцист заканчивается уже во время последней нимфальной линьки. Однако в большинстве случаев аргазиды становятся способными к спариванию лишь через несколько дней после окончания линьки (табл. 44). Вероятно, требуется некоторое время и для окончания превращения сперматид в проспермии и накопления последних в семенных пузырьках. У самцов О. papillipes (Feldman-Muhsam а. Наѵіѵі, 1967) стерильность недавно перелинявших голодных особей связана с отсутствием проспермиев внутри сперматофоров.
Подобные клещи спариваются с самками, но не способны к оплодотворению. Нормальные сперматофоры образуются у них спустя несколько дней или недель после линьки, и питание значительно ускоряет их созревание.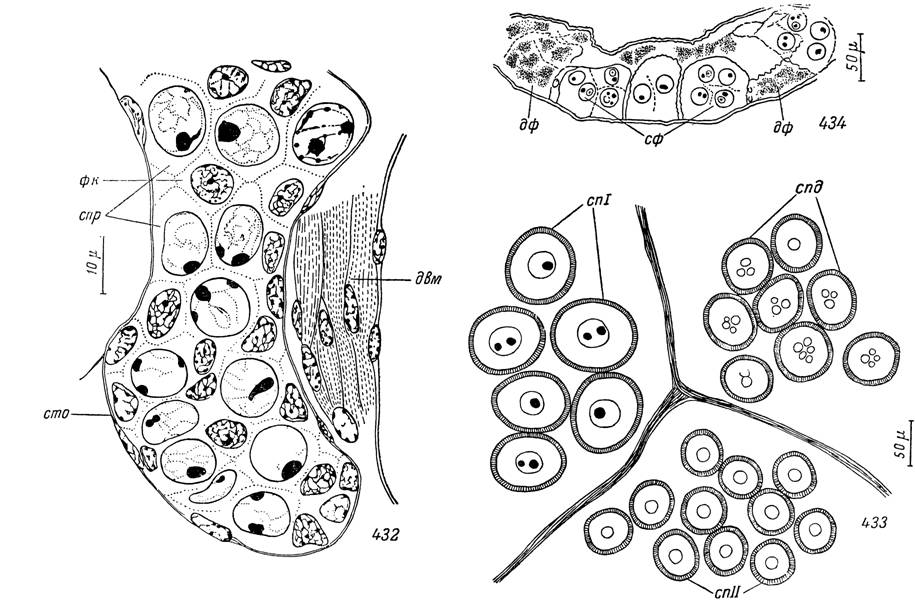
Рис. 432—434. Семенники Argas persicus на разных стадиях жизненного цикла.
432 — участок зачатка семенника личинки; 433 — части трех фолликулов из дистального отдела семенника самца;
434 — участок семенника самца, голодавшего больше двух лет.
спр — сперматогонии; фк — фолликулярные клетки; сто — соединительнотканная оболочка; двм — дорсовентральный мышечный пучок; спі — сперматоциты I порядка в конце роста; спІІ — сперматоциты II порядка; спд — молодые сперматиды; сф — семенной фолликул; дф — дегенерировавший фолликул.Способность самцов аргасовых клещей к спариванию и оплодотворению в первые дни после линьки
В зависимости от количества нимфальных возрастов в жизненном цикле первые сперматоцисты образуются в задних концах семенников уже у напитавшихся личинок или нимф I —II возрастов (рис. 429, 430). Наряду с ростом уже образовавшихся сперматоцитов на нимфальной фазе после каждого питания наблюдается новая волна размножения спер- матогониев, дающих новые порции сперматоцитов. У нимф, готовящихся к линьке в самцов, задние концы семенников содержат молодые сперма- тиды, которые в первые несколько дней после линьки превращаются в зрелые проспермии и выводятся из семенников в семенные пузырьки (рис. 431). В некоторых случаях сперматогенез заканчивался на нимфальной фазе и в семенных пузырьках у только что перелинявших самцов Alveonasus lahorensis и Ornithodoros papillipes в большом количестве обнаруживались готовые проспермии.
Сперматогенез продолжается у самцов в течение нескольких месяцев после линьки, пока не иссякнет запас необходимых для его поддержания веществ и энергии, полученных от нимфальной фазы. Этого запаса, по- видимому, достаточно для того, чтобы большая часть имеющихся в семенниках сперматоцитов закончила свое развитие и превратилась в проспермии.
Голодание вначале вызывает прекращение размножения сперматогоний и остановку образования новых сперматоцитов. Приостановка роста сперматоцитов приводит к прекращению делений созревания. Более длительное голодание приводит к дегенерации крупных сперматоцитов, цитоплазма и ядра которых превращаются в бесструктурную массу. Наиболее стойки к длительному голоданию готовые проспермии, находящиеся в семенных пузырьках, сперматогонии и сперматоциты в, начале роста (рис. 434). Эти элементы сохраняют жизнеспособность несколько лет. После принятия крови в семенниках начинается новая волна сперматогенеза.
У легко переносящих длительное голодание самцов AIveonasus lahorensis и Ornithodoros papillipes подавляющая часть особей сохраняла в наших опытах способность к спариванию и нормальному оплодотворению самок даже после четырехлетнего голодания. У самцов Argas persicus способность к оплодотворению самок утрачивалась черёз 2 года после последнего питания и у Ornithodoros tartakovskyi уже в конце первого года голодания (табл. 45), т. е. несколько раньше гибели клещей от исто-
Способность голодающих самцов аргасовых клещей к спариванию и оплодотворению
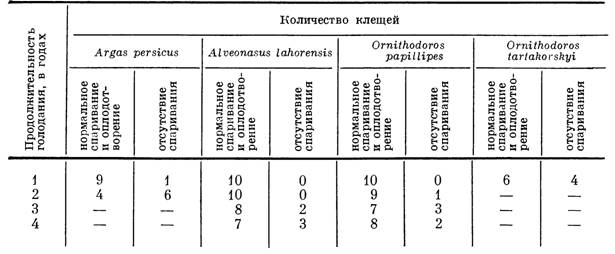
Примечание. Знак минус (—) означает отсутствие жизнеспособных клещей в указанные сроки голодания.
щения. Питание приводило к быстрому (через 2—5 суток) восстановлению нормальной способности к оплодотворению у истощенных особей.
Степень зрелости половых элементов у самцов иксодовых клещей зависит от их физиологического состояния (Балашов, 19566). У Ixodes гі- cinus и /. persulcatus сперматогенез заканчивается еще на стадии линьки. В половых путях недавно перелинявших самцов содержатся зрелые про- спермии, а в сперматоцистах видны все этапы большого роста и спермио- гистогенеза. Самцы способны к оплодотворению, и копуляция может происходить как на теле хозяина, так и в норіах, гнездах и других естественных биотопах с голодными самками. Интересно, что среди Ixodinae широко распространена афагия самцов, так что кровососание уже не играет роли в завершении сперматогенеза (см. стр. 103).
У АтЫуоттіпае голодные самцы не способны к оплодотворению. В их семенниках сперматоциты I порядка останавливаются в развитии перед началом фазы большого роста. Во время кровососания от задних концов семенников распространяется волна большого роста и спермио- гистогенеза. Зрелые проспермии появляются в семепроводах не ранее, чем через 3—5 суток с начала питания, после чего самцы приступают к копуляции. Количество сперматогоний и молодых сперматоцитов постепенно уменьшается, а проспермии растягивают стенки семепроводов, превращающихся в семенные пузырьки.
Строение и функции придаточных желез. Мужские придаточные железы иксодоидных клещей представляют гетерогенные органы, в состав которых входит несколько типов секреторных клеток. Л. Робинсон и Д. Де- видсон (Robinson a. Davidson, 1913—1914) на основании гистологических особенностей описали в придаточных железах Argas persicus два типа железистой ткани — губчатый и гранулярный. Новейшие цитологические и гистохимические исследования (Wagner-Jevseenko, 1958; Till, 1961; Tatchell, 1962b; Балашов, 19646; Chinery, 1965), однако, не подтвердили правомочности классификации лопастей по этому признаку. В зависимости от фазы секреторной активности определенные лопасти или части одной и той же лопасти могут приобретать гранулярную или губчатую структуру. Кроме того, в составе желез выявлено исключительное многообразие типов секреторных клеток, сгруппированных в компактные массы и ограниченных одной лопастью или перемешанных между собой.Наиболее детально изучены тонкое строение и химический состав продуктов секреции придаточных желез самцов Argas persicus (Tatchell, 1962b). Как и у других видов аргазид, развитие входящих в их состав секреторных клеток завершается во время последней линьки и у недавно перелинявших особей в их цитоплазме содержится большое количество секреторных вакуолей. Различия между длительно голодавшими или недавно напитавшимися и спаривавшимися самцами определяются главным образом соотношением числа освободившихся от секрета и заполненных последним клеток.
Передние и задние губчатые лопасти самцов Argas persicus построены из крупных клеток с небольшими ядрами. Цитоплазма имеет вид тонкой сети, занимающей узкие пространства между крупными вакуолями, содержимое которых дает положительные реакции на кислые мукополисахариды (рис. 147).
Непарная вентральная и I пара латеральных лопастей образованы узкими столбчатыми клетками. Ядра последних расположены в базальной зоне, а большую часть клеток заполняют эозинофильные вакуоли диаметром 3—5 мк (рис. 148). В I паре латеральных лопастей вакуоли содержат основные белки, а в непарной вентральной лопасти их содержимое наряду с интенсивным окрашиванием бром-феноловым синим дает положительную реакцию на фенолы и сульфгидрильные группы.
II пара латеральных лопастей содержит два типа железистых клеток. В более крупных клетках с большими сферическими ядрами в процессе секреции происходит отшнуровывание апикальных участков цитоплазмы в виде капель диаметром 4—8 мк (рис. 149). Последние интенсивно окрашиваются гистохимическими реактивами на белки. Цитоплазма более узких и высоких клеток заполнена мукопротеиновыми вакуолями диаметром до 2 мк.
III пара латеральных лопастей образована очень высокими столбчатыми клетками, заполненными белковыми вакуолями диаметром до 5 мк. Содержимое этих вакуолей дает также положительные реакции на присутствие фенолов, дисульфидных групп и полисахаридов. Железистые клетки заднедорсальных лопастей по строению сходны с вышеописанными, но значительно отличаются от них продуктами секреции. В дистальных концах этих лопастей образуется неспецифическая эстераза, в средней части — белки с высоким содержанием триптофана и фенолов и в проксимальных частях — ШИК-положительные вещества.
В главной медиальной лопасти выявлено несколько типов секреторных клеток, которые без определенной закономерности встречаются на всем протяжении лопасти. Клетки с различными секреторными продуктами могут быть перемешаны между собой или собраны в небольшие однородные группы. Морфологически все они довольно однородны. Ядра их лежат в базальной зоне, а цитоплазма заполнена секреторными вакуолями. Часть последних чернится железным гематоксилином, но основная масса интенсивно окрашивается эозином. Гистохимическими реакциями выявлены следующие типы секреторных вакуолей: мукопротеиновые, мукополисахаридные, белковые с высоким содержанием дисульфидных или фенольных групп.
У иксодовых клещей, исключая непитающихся самцов многих видов подсем. Ixodinae, клетки мужских придаточных желез голодных особей не проявляют признаков секреторной активности. Лишь в некоторых клетках главной медиальной лопасти Hyalomma asiaticum на этой стадии были обнаружены эозинофильные вакуоли секрета. Все лопасти придаточных желез голодных самцов этого вида, исключая пару вентро-латеральных, имеют сходное строение. Границы образующих их клеток плохо различимы, объем цитоплазмы невелик и стенки лопастей образованы из массы овальных ядер размером 5—6x3—4 мк, которые тесно прилегают друг к другу и расположены в несколько слоев (рис. 438). Лишь внутренние полости лопастей ограничены узкой цитоплазматической зоной, свободной от ядер. Толщина стенок лопастей на превышает 40—50 мк. В дистальных половинах вентро-латеральных лопастей'уже у голодных особей различимы два типа клеток. Одни более крупные, с ядрами размеров 13—15x7—8 мк, лежат в базальных частях клеток. Между ними вклиниваются более узкие клетки с мелкими ядрами, размером 6—7x4—5 мк, лежащими в центре или в апикальной зоне цитоплазмы (рис. 438).
За первые 3—4 суток питания секреторные клетки придаточных желез самцов Н. asiaticum достигают состояния зрелости. Размеры их увеличиваются в несколько раз главным образом за счет роста объема цитоплазмы, так что толщина стенок лопастей достигает 120—150 мк. Цитоплазма бывает заполнена массой секреторных вакуолей диаметром от 1 до 4—5 мк, содержимое которых выводится в полости лопастей. Многообразие типов секреторных клеток придаточных желез удается выявить только с помощью гистохимических методик, так как на обычных гистологических препаратах клетки с разными продуктами секреции могут выглядеть одинаковыми.
Наиболее гетерогенна по своему клеточному составу главная медиальная лопасть. Высота ее клеток варьирует от 50 до 150 мк; они преимущественно вытянутой формы, с ядрами в базальной и секреторными вакуолями в апикальной половинах (рис. 439). Ядра в соседних клетках могут лежать на разных уровнях и часто могут содержать различные типы секреторных вакуолей. В то же время в одной клетке встречаются вакуоли лишь одинакового химического состава. Гистохимическими методами выявлены следующие типы вакуолей: мукопротеиновые или гликопротеиновые, мукополисахаридные, белковые с высоким содержанием дисуль- фидных групп и липопротеиновые.
Передневентральные лопасти образованы высокими столбчатыми клетками с ядрами на границе с базальной мембраной. Цитоплазма их заполнена мукопротеиновыми вакуолями, так что клеточные границы плохо различимы (рис. 440). Участки лопастей описываемого типа, освободившиеся от секрета, приобретают губчатое строение.
Секреторные клетки непарной вентральной лопасти столбчатые, и их цитоплазма заполнена белковыми вакуолями с высоким содержанием тирозина.
В вентро-латеральных лопастях, сходных по строению со II парой латеральных лопастей Argas persicus, были выявлены два типа клеток. Более крупные клетки со сферическими ядрами в базальной зоне были заполнены очень мелкими вакуолями. Последние концентрировались в апикальных концах клеток, дававших интенсивную положительную реакцию на белки. В процессе секреции наблюдалось отшнуровывание апикальных концов этих клеток в полости лопастей (рис. 441). В более мелких и вытянутых клетках небольшие ядра располагались в центральной зоне, а в цитоплазме были выявлены мукопротеиновые вакуоли. У Наета- physalis spinigera (Chinery, 1965а) в описываемом типе лопастей были обнаружены только клетки с белковым секретом.
Дорсо-латеральные и заднелатеральные пары лопастей отличались одинаковым строением. Стенки их были образованы преимущественно из высоких клеток с белковыми вакуолями. Местами в них встречались также группы из мелких клеток с гомогенной цитоплазмой.
В задневентральных лопастях проксимальные участки были образованы из клеток, заполненных белковыми вакуолями. Последние давали положительное окрашивание по ШИК на полисахариды, а также содержали связанные с белками липоиды и дисульфидные группы. Средняя часть.
лопастей была построена из столбчатых клеток с крупными ядрами и гомогенной цитоплазмой. Они секретировали белки, связанные с липоидами и дисульфидными группами. Их секрет в виде крупных капель поступал в просвет лопастей, вероятно путем отторжения апикальных участков цитоплазмы (рис. 442). Кроме того, в этой же зоне обнаружены узкие и вытянутые клетки, заполненные крупными вакуолями, дающими положительные реакции на белки и полисахариды. Дистальные концы лопастей были образованы из крупных столбчатых клеток с мелкими вакуолями. Последние содержали белки, связанные с полисахаридами и липоидами.
Строение сперматофора. Иксодоидным клещам, как и другим группам наземных хелицеровых, свойственно сперматофорное оплодотворение. Зрелые проспермии клещей переносятся в половые пути самок упакованными в специальные пакеты — сперматофоры.
Сперматофор представляет сферический мешок с узкой шейкой, которая может быть ввернута внутрь шара.
Тонкое строение сперматофора известно только для Argas persicus (Tatchell, 1962b). У этого вида стенка сперматофора представляет двухслойную кожистую оболочку. Наружный слой, толщиной 1 мк, образован из фосфолипопротеинов, а внутренний слой, в 7 мк, — из белков (рис. 151). Снаружи оболочка сперматофора покрыта тонким слоем муцина. Внутри сперматофора находится очень большое количество проспермиев, которые заключены во внутреннем мешке — сперматодозе, стенки которой образованы тонкой мукопротеиновой мембраной. Вершина шейки состоит из муко- протеинов и муцинов. Под ней лежит пара окруженных белковыми оболочками скоплений эстеразы, а на границе с проспермиями расположена зона из секрета III пары латеральных лопастей придаточной железы.
Формирование сперматофора прослежено у нескольких видов арга- совых клещей, включая и Ornithodoros papillipes (Feldman-Muhsam, 1967). В начале спаривания из полового отверстия самца выталкивается наружная кожистая оболочка сперматофора (эктосперматофор). Затем в нее в течение нескольких секунд поступают сперма, семенная жидкость, протеиновые гранулы и симбиотический грибок Adlerocystis sp. После этого в шейку в виде пробки вводится оболочка сперматодозы (эндосперматофор). Выворачивание и расправление сперматодозы и заполнение ее содержимым сперматофора происходит уже после перенесения последнего к влагалищу самки.
Спаривание. Процесс копуляции аргасовых клещей достаточно подробно описан в литературе (Павловский, 1940а; Соколов, 1956; Wagner- Jevseenko, 1958). При спаривании партнеры соединяются своими вентральными поверхностями, причем самцы активно удерживаются на самках с помощью ног. Затем самец вводит свой хоботок через наружное половое отверстие во влагалище и медленными движениями взад и вперед расширяет его просвет. После этого самец выдвигает свой хоботок, подгибает его на вентральную сторону и захватывает сперматофор, выступающий к этому времени из мужского полового отверстия. Сперматофор переносится на хоботке к половому отверстию самки, и его вытянутый носик вводится во влагалище при погружении в последнее хоботка. Процесс спаривания продолжается несколько минут, и самки способны к копуляции в любое время после линьки.
При переносе сперматофора пальцами хелицер делается надрез у конца его шейки. Сквозь него тонкостенная сперматодоза проталкивается во влагалище и сразу же заполняется спермой и семенной жидкостью. Кожистая оболочка сперматофора остается снаружи женского полового отверстия и может сохраняться здесь в течение нескольких дней. Сперматодоза проталкивается в матку перистальтическими сокращениями мускулатуры влагалища и соединительной трубки. В матке может находиться несколько сперматодоз, так как самки аргасовых клещей способны к многократным спариваниям. Проспермии остаются внутри сперматодозы от нескольких дней до нескольких недель или даже месяцев, и в этот период происходит завершение их созревания и превращения в спермиофоры. Из сперматодозы выходят зрелые спермиофоры, активно мигрирующие в яйцеводы.
Детальное описание процесса копуляции иксодовых клещей отсутствует. Как и у аргазид, партнеры во время спаривания соединяются своими вентральными поверхностями и самцы удерживаются в этом положении с помощью ног (рис. 7). Перед переносом сперматофора в женское половое отверстие вводится хоботок самца. Однако сам сперматофор, по наблюдениям над Ixodes holocyclus (Moorhouse, 1966), перемещается к женскому половому отверстию благодаря скользящим движениям самца вдоль вентральной стороны идиосомы самки. Хоботок участвует лишь в подрезании конца шейки и введении ее в женское половое отверстие.
Процесс спаривания занимает от 10—15 мин. до нескольких часов. Самцы Ixodes ricinus, по нашим наблюдениям, могут оставаться прикрепленными к самкам по нескольку суток. Голодные самки АтЫуоттіпае не способны к спариванию, а Ixodinae хотя и могут спариваться, но разрыв сперматодоз и выход спермиофоров в яйцеводы наблюдаются в конце созревания ооцитов. Имеющегося в семенных пузырьках запаса готовых проспермиев достаточно для последовательного оплодотворения нескольких самок. В опытах Г. С. Первомайского (1954) один самец Нуаіотта plumbeum последовательно оплодотворил 25 самок. В наших опытах самцы родов Нуаіотта, Dermacentor и Rhipicephalus во время совместного с самками кормления на кроликах последовательно спаривались с несколькими самками. При одиночном содержании недавно перелинявших самцов Ornithodoros papillipes и Argas persicus с девственными напитавшимися самками 1 самец в течение нескольких дней оплодотворял от 4 до 9 самок.
Еще по теме ЦИТОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РАЗМНОЖЕНИЯУ САМЦОВ:
- Цитологические и гистологические особенности костного мозга и периферической крови у больных МДС
- ЧАСТЬ ДВАДЦАТАЯ РАЗЛИЧНЫЕ СОСТОЯНИЯ ОРГАНОВ РАЗМНОЖЕНИЯ У МУЖЧИ
- Диагностика и особенности цитологической, кольпоскопической картины папилломавирусной инфекции, гистологического метода исследования у беременных. Состояние влагалищного биотопа у женщин с папилломавирусной инфекцией.
- ПИТАНИЕ САМЦОВ
- Органы размножения самцов.
- ОПРЕДЕЛИТЕЛЬ САМЦОВ МОСКИТОВ
- Цитологическая диагностика.
- Цитологическое исследование.
- Аспирационная биопсия и цитологическое исследование
- 3.2. Методики цитологического исследования
- Цитологическое отделение
- Цитологические и кольпоскопические картины у обследованных беременных.
- Цитологическое исследование мокpоты
- Цитологическое исследование отделяемого из соска
- 1.2 Этапы развития цитологического метода в диагностике заболеваний желудка
- Цитологическое исследование мокроты
- Цитологическое исследование.