СПИРОХЕТЫ
Аргасовые клещи служат переносчиками значительного числа патогенных для позвоночных и человека спирохет рода Borrelia, рассматриваемых в литературе в качестве сильно видоизмененной группы бактерий.
Для иксодид известна лишь передача клещами родов Boophilus и ВЫрі- cephalus evertsi возбудителя спирохетоза крупного рогатого скота Borrelia theileri. Различным сторонам взаимоотношений клещей со спирохетами посвящена обширная литература, библиография которой приводится в ряде последних обзоров (Поспелова-Штром, 1953; Москвин, 1960; Петрищева и Жмаева, 1962; Павловский и Скрынник, 1965; Burgdorfer, 1951; Varma, 1962, и др.). Заболевания людей клещевым спирохетозом, по данным ВОЗ, регистрируются во многих странах. Особенно часты они в Восточной и Центральной Африке (переносчик Ornithodoros moubata и возбудитель Borrelia duttoni), странах Средиземноморья и Ближнего Востока (главный переносчик Ornithodoros erraticus и возбудитель Borrelia hispanica), а также в центральных и западных штатах США, в Мексике и в Южной Америке до северной части Аргентины (несколько видов клещей рода Ornithodoros и спирохет).На территории СССР в республиках Средней Азии клещ Ornithodoros papillipes передает Borrelia sogdiana и в Закавказье клещ Ornithodoros verrucosus—спирохету Borrelia caucasica. Отдельные очаги спирохетозов известны и за пределами этих районов (юг Казахстана, Украины и Северный Кавказ). Второстепенное значение имеют клещи Ornithodoros tartakovskyi, О. nereensis и О. alactagalis, которые передают грызунам спирохет Borrelia latyschevi, В. nereensis и В. агтепіса. Клещи Argas persicus на всем протяжении своего ареала передают возбудителя птичьего спирохетоза Borrelia anserinum, который местами причиняет существенный ущерб птицеводству.
Систематика спирохет рода Borrelia остается весьма запутанной из-за отсутствия надежных морфологических и иммунобиологических критериев вида.
Чаще всего при установлении видовой принадлежности спирохет исходят из вида клеща-переносчика. Метод ксенодиагностики был успешно использован с клещами Ornithodoros turicata, О. parkeri и О, hermsi из Северной Америки, отличающимися высокой специфичностью к определенным видам спирохет (Davis, 1943). На основании многочисленных опытов с И видами клещей-орнитодорин и 5 видами спирохет А. Н. Скрынник (1966) также считает, что нормально клещи могут передавать только свойственные им виды спирохет. Передача «чужих» спирохет наблюдалась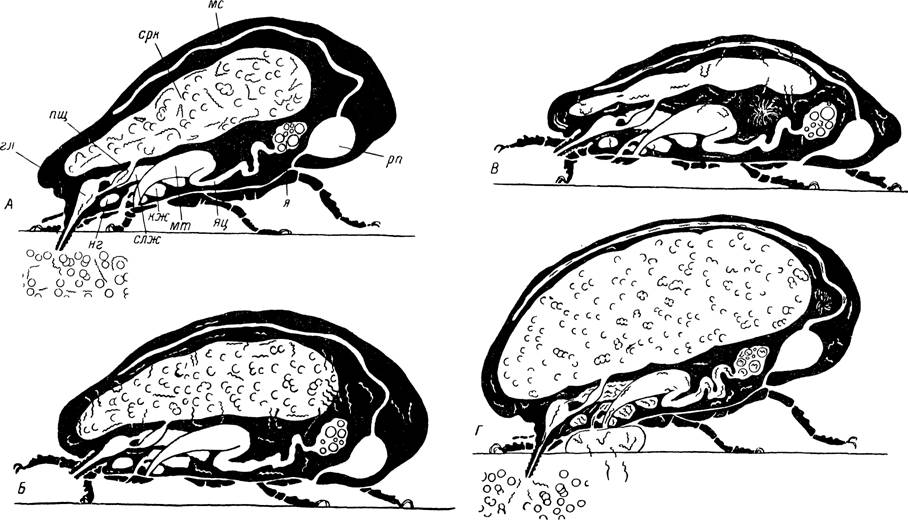
Рис. 464. Циркуляция и передача спирохет Borrelia duttoni клещом Ornithodoros moubata.
А — поглощение инфицированной крови; Б — миграция спирохет из полости кишечника в полость тела; В — размножение спирохет в гемолимфе, заселение ими слюнных и коксальных желез, нервного ганглия, мальпигиевых сосудов и других внутренних органов при почти полном исчезновении из кишечника; Г — питание инфицированного клеща; видно выделение спирохет вместе со слюной и коксальной жидкостью. гл — глотка; пщ — пищевод; срк — средняя кишка; мс — мальпигиев сосуд; рп — ректальный пузырь; я — яичник; яц — яйцевод; мт — матка; слж — слюнная железа; кж — коксальная железа; нг — нервный ганглий (по Burgdorfer, 1951).у немногих особей опытных партий. Несмотря на высокую специфичность отношений клещей и спирохет, этот критерий нельзя переоценивать. Известно, что штаммы Borrelia duttoni, В. hispanica и В. crocidurae удалось успешно культивировать не только в необычных для них видах клещей, но даже во вшах (Haberkorn, 1963), так что эта проблема требует дальнейшего изучения.
В организм клеща спирохеты поступают вместе с кровью инфицированного хозяина. Дальнейшая судьба возбудителя длительное время была предметом дискуссий в связи с гипотезой о существовании у них гранулярных или фильтрующихся стадий развития. Эти взгляды, однако, не получили подтверждения в новейших исследованиях по циркуляции Borrelia duttoni в клещах Ornithodoros moubata (Burgdorfer, 1951), Borrelia sogdiana в Ornithodoros papillipes (Теравский, 1959) и Borrelia an- serinum в Argaspersicus (Никитина, 1964, 1965).
В этих работах приводятся убедительные данные об обнаружении морфологически нормальных спирохет на любых сроках после инфицирования и о размножении их путем простого поперечного деления. Представления о циркуляции и передаче спирохет клещами Ornithodoros moubata суммированы на рис. 464.Поглощенные спирохеты в первые дни после питания обнаруживаются преимущественно в среднем отделе кишечника. Вначале они встречаются по всему содержимому полости кишки, а на более поздних сроках концентрируются на поверхности эпителиальных клеток стенок, проникают между ними в межклеточные пространства и к базальной мембране. Значительное уменьшение количества спирохет в полости кишечника наблюдалось у Ornithodoros moubata через 12—16, ay О. papillipes — через 10—12 и у Argas persicus — через 13—20 дней после заражения. Отдельные спирохеты были обнаружены и на более поздних сроках. В первые дни после инфицирующего кормления у А. persicus наблюдалось даже размножение спирохет в полости кишечника. Исчезновение спирохет из кишечника на более поздних сроках, вероятно, связано с неблагоприятными изменениями его среды по мере переваривания поглощенной крови.
Уже в первые часы или даже минуты после пЪступления инфицированной крови в кишечник спирохеты обнаруживаются в гемолимфе. Проникновение их в полость тела клеща не связано с разрывами стенки кишечника, как считали некоторые исследователи (Аракчеева, 1963), и обеспечивается активными движениями и, возможно, лизирующими факторами самих спирохет. Количество спирохет в гемолимфе в первые дни после заражения увеличивается за счет их миграции из кишечника. На более поздних сроках их содержание в гемолимфе может сильно варьировать. У зараженных голодных клещей присутствие возбудителя в гемолимфе далеко не постоянно (у Argas persicus только у 12—16% особей) и зависит как от видовых особенностей переносчика-возбудителя, так и от физиологического состояния клеща. Питание обычно стимулирует размножение спирохет и количество их в этот период может увеличиваться.
Из гемолимфы спирохеты проникают во многие внутренние органы, причем в распределении их в организме клеща наблюдаются определенные видовые различия.
У Ornithodoros moubata спирохеты Borrelia duttoni чаще всего встречались в нервном ганглии, а затем по частоте обнаружения шли коксальные железы, слюнные железы, органы размножения и мальпигиевы сосуды. Наиболее интенсивное размножение и накопление возбудителя наблюдалось в стенках мальпигиевых сосудов и в нервном ганглии. В то же время, по наблюдениям М. Вармы (Varma, 1962), при заражении нимф этого же вида клеща спирохетами В. turicata последние наиболее часто обнаруживались в слюнных железах. У зараженных Argas persicus возбудитель был обнаружен в нервном ганглии в 67—76% случаев, в слюнных железах — в 71%, в мальпигиевых сосудах — в 41 — 47% й в органах размножения — в 23—27%. Как правило, у видов рода Ornithodoros слюнные железы половозрелых особей заражены спирохетами значительно слабее и реже, чем у нимф. Преимущественную локализацию спирохет в определенных органах клещей связывают с органотропизмом возбудителя к определенным тканям клеща, что было доказано в опытах in vitro (Sarasin, 1959). Нервные ганглии и коксальные железы клещей притягивали к себе значительно большее количество спирохет по сравнению со слюнными железами, мальпигиевыми сосудами и органами размножения. Установлено, что привлекающим фактором служат олигосахариды, выделяемые этими органами (Grim u. Blatter, 1960). Важную роль в распределении спирохет в организме может играть и разная способность их к размножению в различных тканях.Особенности локализации спирохет в организме клещей предопределяют и пути их передачи позвоночным во время питания. Клещ приобретает инфицирующую способность только после того, как спирохеты проникнут в его гемолимфу и из нее в коксальные и слюнные железы. Возбудитель может быть обнаружен в слюне и коксальной жидкости подобных особей, но участие их в передаче инфекции различно у разных видов. У клещей (iOrnithodoros moubata), регулярно выделяющих большое количество коксальной жидкости во время акта питания на теле хозяина, последняя содержит значительное количество спирохет и представляет основной путь передачи инфекции.
У половозрелых особей О. moubata слюнные железы редко содержат спирохеты и передача их с коксальной жидкостью становится единственным путем заражения позвоночных. Напротив, у О. papillipes, Argas persicus и других видов, выделяющих коксальную жидкость по окончании питания и оставлении тела хозяина, передача спирохет возможна только со слюной (Теравский, 1959; Никитина, 1965; Varma, 1962). Передача спирохет с экскретами клещей, по наблюдениям над Ornithodoros moubata (Burgdorfer, 1951), отсутствует, несмотря на сильное инфицирование мальпигиевых сосудов. Маловероятна и возможность непосредственного проникновения спирохет из переднего отдела кишечника в ранку на теле хозяина без предшествующей внутриполостной миграции и заселения слюнных желез, как ошибочно считаетС. Г. Аракчеева (1963).
Аргасовые клещи способны к заражению спирохетами при кормлении на больных животных на всех фазах жизненного цикла и к последующей передаче возбудителя по ходу метаморфоза. Значение отдельных фаз развития клещей в передаче спирохет, как показали опыты Е. Н. Павловского и А. Н. Скрынник (1939, 1952, 1965), неодинаково. Личинки Ornithodoros papillipes передают их лишь в исключительных случаях независимо от того, получили ли они возбудителя трансовариально или при прерывистом кормлении. Легче всего передача спирохет позвоночному происходит на нимфальной фазе. У половозрелых особей иногда наблюдалась утрата способности к передаче. Последнее чаще встречается у многократно питавшихся самок, тогда как длительное голодание (до 10 лет) не приводило к исчезновению спирохет из организма клещей. При кормлении на морских свинках подобные истощенные клещи нормально осуществляли передачу инфекции. Снижение инфицирующей способности отмечено также для имаго О. moubata (Burgdorfer, 1951) и О. turicata (Varma, 1962), причем у этих видов наблюдалось снижение процента заражения спирохетами слюнных желез по сравнению с нимфальной фазой.
Важную роль в циркуляции спирохет в природных очагах играет исключительная способность клещей к трансовариальной передаче спирохет.
Так, у клещей О. papillipes это явление наблюдалось примерно у трети зараженных самок. К трансовариальной передаче были способны и самки дочерних поколений, питавшиеся только на здоровых животных.В результате удалось поддерживать Borrelia sogdiana исключительно за счет циркуляции в организме клещей на протяжении 3 поколений в течение 26 лет (срок наблюдений; Павловский и Скрынник, 1951, 1965). В опытах с Ornithodoros moubata трансовариальная передача прослежена для 5 поколений (Geigy u. Aeschlimann, 1964) и выяснены ее цитологические особенности (Aeschlimann, 1958). Спирохеты были обнаружены у 2— 98% потомства инфицированных самок и из них до 60% передавали их следующему дочернему поколению. Возбудитель внедряется из гемолимфы только в молодые ооциты до образования вокруг них «хориона». После этого они уже не могут быть инфицированы. В ооцитах спирохеты сохраняют нормальную морфологическую форму и располагаются в цитоплазме между желточными шарами. С началом дифференцировки центральной нервной системы они сразу же мигрируют в ее эмбриональную закладку и остаются в ней до окончания эмбриогенеза. Заселение спирохетами других внутренних органов, включая и слюнные железы, происходит уже на личиночной фазе. Частота трансовариальной передачи определяется видовой принадлежностью клещей. Например, у Ornithodoros turicata (Varma, 1962) или Argas persicus (Никитина, 1964) она была значительно реже, чем у Ornithodoros papillipes и О. moubata.
В связи с заражением семенников спирохеты вместе с проспермиями могут проникнуть внутрь сперматофоров и во время оплодотворения поступить в организм незараженной самки. Подобный путь передачи был обнаружен в 1.8% опытов спаривания «чистых» самок с зараженными самцами О. moubata (Wagner-Jevseenko, 1958).
Заражение клещей спирохетами, по-видимому, не отражается на жизнеспособности переносчика. В то же время имеются наблюдения об изменениях вирулентности спирохет при циркуляции их только в клещах. Так, Borrelia duttoni, находившиеся в организме клещей на протяжении 3—5 поколений, теряли инфекциозность для белых мышей (Aeschlimann, 1958; Geigy u. Aeschlimann, 1964). Наоборот, спирохеты того же вида, на протяжении нескольких лет пассировавшиеся через белых мышей, не могли заражать клещей Ornithodoros moubata при инфицирующем кормлении. Заражение в этом случае наблюдалось только после внутриполостного введения возбудителя (Varma, 1962). Таким образом, клещи могут служить резервуарными хозяевами спирохет в природе, но для поддержания достаточной вирулентности штаммов необходима их регулярная циркуляция между восприимчивыми позвоночными и клещами.
Иксодоидные клещи, возможно, связаны и с лептоспирами рода Leptospira, очень близкими к роду Borrelia. Удалось выделить лептоспир из собранных в природе клещей Dermacentor marginatus (Крепкогорская и Ремнецова, 1957), Ornithodoros tartakovskyi (Благодарный, 1959) и некоторых других видов. После экспериментального заражения Leptospira icterohaemorrhagiae сохранялись в клещах Hyalomma asiaticum 22— 27 дней, a Leptospira canicola в Rhipicephalus sanguineus — 36 дней (Со- лошенко, 1959). Этому же автору удалось осуществить передачу возбудителя восприимчивым животным путем кормления на них зараженных клещей. При инфицировании Leptospira ротопа клещей Ornithodoros turicata (Burgdorfer, 1956) возбудитель сохранялся в них 232—518 суток. Кормлением подобных клещей на морских свинках удалось осуществить передачу инфекции и было доказано выделение больших количеств лептоспир с коксальной жидкостью. После поступления в кишечник клещей крови с лептоспирами последние вскоре проникали сквозь его стенки в полость тела. Установлено интенсивное размножение возбудителя в гемолифме и заселение им слюнных и коксальных желез, нервного ганглия, мальпигиевых сосудов и органов размножения. В то же время трансовариальная передача Leptospira ротопа клещами отсутствовала.
Несмотря на случаи успешного лабораторного заражения и последующей передачи возбудителя клещами, редкость находок спонтанно инфицированных клещей не дает оснований рассматривать эту группу членистоногих эпидемиологически важными переносчиками лептоспир.
Еще по теме СПИРОХЕТЫ:
- Методы исследования бледной спирохеты.
- Часть третья Венерические болезни
- 11. 3.2.6. Нейроборрелиоз
- ПЕРСИДСКИЙ КЛЕЩ
- 11.6. Сифилис нервной системы
- ПОСЕЛКОВЫЙ КЛЕЩ
- 1.3.5 Treponema denticola
- 12.4. Краткая характеристика очага бактериологического (биологического) поражения
- Эндемический возвратный тиф
- Ранний врожденный сифилис
- Тиф возвратный вшивый.
- Значение лихорадки
- ОБЩИЕ ВОПРОСЫ ПАРАЗИТОЛОГИИ
- Склерома гортани.
- Глава 2. СИФИЛИС
- Эпидемический возвратный тиф
- СИФИЛИС
- ЧАСТОТА ВЫЯВЛЕНИЯ ИММУНОГЛОБУЛИНОВ М К БОРРЕЛИЯМ У ЛИЦ В РАЗЛИЧНЫХ РЕГИОНАХ РЕСПУБЛИКИ БЕЛАРУСЬ
- КЛЕЩИ И ВОЗБУДИТЕЛИ ТРАНСМИССИВНЫХ ИНФЕКЦИЙ ПОЗВОНОЧНЫХ