Мужская половая система
Мужская половая система Ixodoidea состоит из двух трубчатых семенников, полностью разделенных у АтЫуоттіпае, частично сливающихся вдоль медиальной поверхности у Ixodes ricinus и соединяющихся задними концами у аргасовых клещей, парных семепроводов, семеизвергательного канала и комплекса придаточных желез (рис.
142)Семенники. Парные семенники самцов Hyalomma asiaticum расположены по бокам тела от заднего края центральной нервной массы до уровня концов стигмальных пластинок. В задней трети они изгибаются внутрь и заметно утолщаются (рис. 142).
У Argas persicus, Alueonasus lahorensis ж Ornithodoros papillipes семенники сливаются своими концами и образуют подковообразную гонаду. У старых особей место соединения представляет тонкий соединительнотканный тяж, лишенный половых элементов. Ряд исследователей (СЬгі- stophers, 1906; Robinson a. Davidson, 1913—1914; Roshdy, 1961a) неправильно называет его семенником, а настоящие семенники считает семенными пузырями. На самом деле описываемый центральный участок гонады несомненно образуется в связи с исчерпанием в нем половых элементов. У нимф и молодых самцов семенники соединяются непосредственно своими толстыми концевыми участками (рис. 430—431).
По своему строению семенники Ixodoidea относятся к типу трубчатых гонад. Снаружи их окружает тонкая соединительнотканная оболочка, а толстая стенка вокруг небольшого центрального просвета состоит из сперматоцист различной стадии зрелости. Каждая из сперматоцист обладает собственной тонкой стенкой эпителиального происхождения (рис. 435).
Половые элементы созревают в строго определенном направлении сзади-наперед. Впервые задне-передний градиент сперматогенеза был описан у аргасовых клещей (Горощенко, 1960; Wagner-Jevseenko, 1958). По нашим наблюдениям, он свойствен и всем изученным видам иксодовых клещей.
В вытянутом семеннике Hyalomma asiaticum и Ixodes ricinus от его переднего конца каудально располагаются зоны первичных спермато- гониев, вторичных сперматогониев, сперматоцитов I порядка на стадиях малого и затем большого роста, сперматоцитов II порядка, сперматид на различных стадиях спермиогистогенеза и проспермиев.
Зрелые проспермии передвигаются по центральному просвету от заднего конца в направлении семепроводов. Они лишены способности к активным самостоятельным движениям, и их проталкивание вперед облегчается перистальтическими сокращениями стенок семенника.Выводные пути. Семепроводы представлены короткими парными трубками, которые соединяются перед впадением в непарную половую полость — atrium genitale. Последняя эктодермального происхождения и выстлана тонкой кутикулой. Половая полость служит местом соединения основных частей полового аппарата. В нее открываются придаточные половые железы, а в передневентральном направлении она переходит в семеизвергательный канал (рис. 428).
Свободные от проспермиев или слабо растянутые последними семе- проводы представляют тонкие трубки диаметром в 25—40 мк, стенки которых образованы внутренним слоем столбчатых или кубических эпителиальных клеток высотой в 15—20 мк с небольшими сферическими ядрами диаметром в 5—6 мк. Снаружи эпителиальный слой окружен тонкой соединительнотканной оболочкой с одиночными тонкими мышечными волокнами (рис. 145). При заполнении проспермиями стенки семепроводов могут очень сильно растягиваться и превращаются в тонкую мембрану (толщиной в 2—3 мк). Клеточные границы в мембране неразличимы, а сильно уплощенные ядра лежат на значительном расстоянии друг от друга. В описываемом состоянии семепроводы часто называют семенными пузырьками.
Семеизвергательный канал образован короткой дорсо-вентрально уплощенной трубкой, идущей от половой полости в передневентральном направлении. Под центральной нервной массой канал открывается наружным половым отверстием. Последнее спереди прикрыто выдающейся назад лопастью кутикулы вентральной стенки тела — апроном.
Внутри семеизвергательный канал покрыт кутикулой. В передней половине канала на дорсальной стороне кутикула склеротизована, а на вентральной — гибкая и собрана в складки. В гибких участках она состоит из эпикутикулы и слоистой эндокутикулы. В склеротизованных частях к ним добавляется мезокутикула.
В задней части канала мезоку- тикула утончается и продолжается до половой полости тремя узкими ветвями — медиальной и боковыми. Эпителий семеизвергательного канала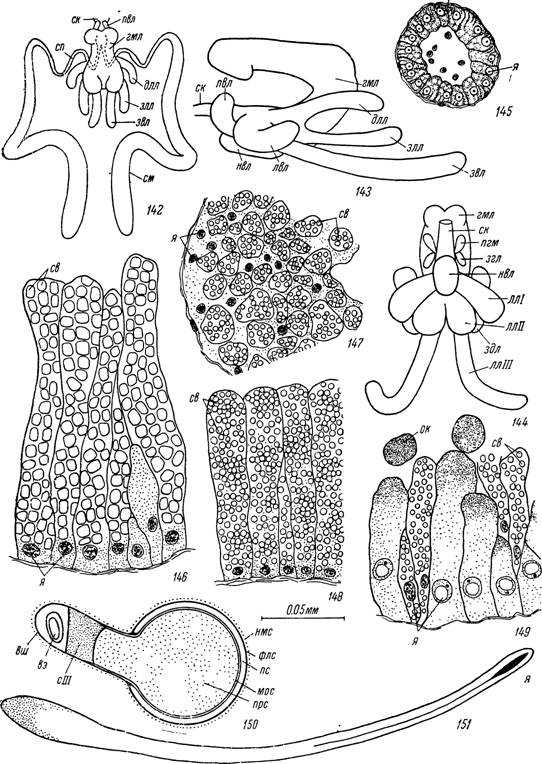
Рис. 142—151. Строение половой системы самцов иксодоидных клещей. 142 — половая система Hyalomma asiaticum, вид сверху; 143 — придаточная железа Н. asiaticum, вид сбоку; 144 — придаточная железа Ornithodoros papillipes, вид снизу.; 145 — поперечный срез семепровода Hyalomma asiaticum,; Argas persicus: 146 — участок стенки главной медиальной лопасти, 147 — то же передней губчатой лопасти, 148 — то же I пары латеральных лопастей, 149 — то же II пары латеральных лопастей и 150 — сперматофор на продольном разрезе (Tatchell, 1962b); 151 — зрелый спермиофор Hyalomma asiaticum.
ск — семеизвергательный канал; гмл — главная медиальная лопасть придаточной железы; пел — передневентральная лопасть; лвл — вентро-латеральная лопасть; пел — непарная вентральная лопасть; длм — дорсо-латеральная лопасть; злл — заднелатеральная лопасть; звл — задневентральная лопасть; пгм — передняя губчатая лопасть; згл — задняя губчатая лопасть; ллі — первая пара латеральных лопастей; ллІІ — вторая пара латеральный лопастей; ллІІІ — третья пара латеральных лопастей; здл — заднедорсальная лопасть; сп — семепровод; см — семенник; се — секреторные вакуоли; ок — отторгающиеся апикальные концы секреторных клеток; я — клеточные ядра; нмс — наружный му- циновый слой; флс — фосфолипопротеиновый слой оболочки спе рматофора; пс — протеиновый слой оболочки сперматофора; мос — мукопротеиновая оболочка сперматодозы; пре — проспермии; вш — вершина шейки сперматофора; вэ — вакуоль с эстеразой; dll — секрет III пары латеральных лопастей.образован кубическими или плоскими клетками, строение которых у половозрелых особей не меняется.
Придаточные железы. Многолопастная мужская придаточная железа прикрывает сверху семеизвергательный канал, располагаясь позади от мозга и под отростками средней кишки.
Она сильно расчленена, причем форма и размеры отдельных ее лопастей значительно варьируют в пределах над семейства.У аргасовых клещей анатомические особенности придаточной половой железы описаны для Argas persicus (Robinson a. Davidson, 1913— 1914; Tatchell, 1962b), A. vespertilionis (Roshdy, 1961a), A. boueti (Roshdy,
1962) , A. transgariepinus (Roshdy, 1963), A. brumpti (Roshdy, 1966) и Ornithodoros moubata (Wagner-Jevseenko, 1958). Нами исследовано ее строение у клещей А г gas persicus, Alveonasus lahorensis и Ornithodoros papil- lipes. У всех изученных видов были обнаружены одни и те же лопасти, и различия были связаны лишь с их взаимным расположением и соотношением размеров. У самцов Ornithodoros papillipes наиболее массивна непарная главная медиальная лопасть придаточной железы, передний конец которой непосредственно переходит в стенки семеизвергательного канала. Дистальная половина этой лопасти изгибается на спинную сторону и вперед, прикрывая сверху проксимальную часть лопасти и семеизвергательный канал (рис. 144).
Сразу же позади места соединения железы с семеизвергательным каналом с боков главной медиальной лопасти отходят небольшие парные передняя и задняя губчатые лопасти, а снизу — непарная вентральная лопасть. Далее назад с боков отходят три пары латеральных лопастей и парная заднедорсальная лопасть. Третья пара латеральных лопастей продолжается дальше остальных в сторону заднего конца тела и отличается молочно-белой окраской, тогда как другие лопасти полупрозрачные. В работе О. Вагнер-Евсеенко (Wagner-Jevseenko, 1958) лопасти придаточной железы обозначены буквами латинского алфавита от А до Н, из которых А отвечает главной медиальной лопасти, В — заднедорсальной паре, С, D и Е — III—I парам латеральных лопастей, F — непарной вентральной лопасти, a G и Н — задней и передней парам губчатых лопастей.
У иксодовых клещей анатомические особенности мужских придаточных желез достаточно подробно описаны для Hyalomma anatolicum (Yalvac, 1939а), Н. asiaticum (Балашов, 19646), Dermacentor andersoni (Douglas, 1943), Rhipicephalus appendiculatus (Till, 1961) и Haemaphy- salis spinigera (Chinery, 1965a).
Как и у аргазид, наиболее массивна главная медиальная лопасть, которая может раздваиваться на дистальном конце и давать парные боковые выросты в месте соединения ее дорсальной и вентральной ветвей. У Н. asiaticum (рис. 142, 143) размеры вентральной ветви главной медиальной лопасти немногим меньше, чем дорсальной ветви, так что в сагиттальном сечении орган напоминает букву V, направленную углом назад. От переднего конца вентральной ветви главной медиальной лопасти до ее изгиба на спинную сторону отходят следующие лопасти: передневентральные, вентро-латеральные, непарная вентральная, дорсо-латеральные, заднелатеральные и задневентральные. У голодных самцов наиболее массивны, не считая главной медиальной лопасти, задневентральная пара лопастей, а у напитавшихся — заднелатеральные. Последние могут достигать уровня заднего конца ректального мешка.Гомологизация лопастей придаточных желез у аргасовых и иксодовых клещей затруднительна в связи с различным клеточным составом и характером секреции в анатомически сходных частях желез у представите- телей этих двух семейств. Цитологические особенности и секреторная активность этих органов подробно рассматриваются в разделе «Размножение» (см. стр. 243).
Еще по теме Мужская половая система:
- Мужская половая система
- Мужская половая система
- Опухоли мужских половых органов
- ГЛАВА 11 Клиническая патология мужской и женской репродуктивных систем
- Женская половая система
- Женская половая система
- Женская половая система
- Половая система
- 2.1. Эмбриогенез и дифференцировка органов репродуктивно-половой системы
- Половая (гендерная) идентичность и половое формирование
- СПЭ потерпевших по делам о половой неприкосновенности и половой свободе личности
- Методы мужской контрацепции
- Параграф седьмой. Способы получить мужское потомство
- Параграф двадцать пятый. Об анатомии мышц мужского члена
- Клинические аспекты женского и мужского бесплодия
- Микробиологические метаболические особенности лиц мужского и женского пола
- Нарушения половых хромосом