1.5.3. Нарушения в системе иммунитета при старении
прослеживаются на медиаторном, клеточном и молекулярном уровнях, затрагивают все главные её компоненты – стволовые клетки, Т- и В-лимфоциты, макрофаги. Однако, наи-более важные возрастные иммунологические изменения связаны с ослабле-нием Т-клеточного компонента иммунной системы, с инволюцией тимуса и снижением способности системы формировать функционально полноценные Т-клетки (Иванов, Иванова, 1987; Ershler, 1988; Асфандиярова и др., 1996; Antonaci et al., 1996; Ptak, Szczepanik, 1998; Chakravarti, Abraham, 1999).
Уменьшение числа митотически компетентных Т-клеток по мере старения связывается также с существованием известного предела Хейфлика и у клеток иммунной системы, следствием чего являются снижение клеточного иммунитета и повышение смертности от инфекционных заболеваний в пожилом возрасте (Effros, 1998). Атрофируются также селезёнка и лимфатические узлы, падает содержание периферических эффекторных клеток и увеличивается число незрелых лимфоцитов вследствие задержки дифференцировки (Донцов, 1989).Данные такого рода продолжают пополняться всё новыми исследованиями. Показано, например, что у 26-месячных крыс Sprague-Dawley относительное содержание лимфоцитов в периферической крови и селезёнке снижено по сравнению с таковым у крыс в возрасте 3-х и 13-ти мес. Уменьшенными также были уровни Т-хелперов, естественных киллеров (NK) и общее содержание Т-клеток (Flaherty et al., 1997). О снижении с возрастом подопытных животных (крыс Wistar) литической активности NK-клеток сообщили Даусон с соавт. (Dawson et al., 1997). Ими же было отмечено, что недостаток витамина А в рационе по сравнению с нормой снижает уровень и процентное содержание указанных киллеров в периферической крови, а избыток витамина А – увели-чивает. Этот интересный факт естественно связать с окислительным поврежде-нием самих эффекторных клеток при дефиците антиоксидантов или неэффек-тивности антиоксидантной системы и защитой их от повреждения в обратном случае (см.
ниже).Сходные факты приведены в работе, где изучали влияние различных анти-оксидантов на киллерную активность мононуклеарных клеток из лимфоузлов, селезёнки, тимуса и перитонеальных лимфоцитов мышей BALB/c в возрасте 8-72 недель. Тиоприлин, N-ацетилцистеин, аскорбиновая кислота и витамин Е повышали киллерную активность у мышей всех возрастных групп, но более эффективными из них оказались первые два. Особенно благоприятным было действие антиоксидантов для старых животных, у которых с возрастом падает естественная кисллерная активность (Ferrandez et al., 1999). Те же, по существу, эффекты наблюдаются и в процессе старения людей. У пожилых лиц снижа-ются общий уровень Т-лимфоцитов, их субпопуляции CD4+ и особенно CD8+, а также содержание NK-клеток. У них же ослаблена фагоцитарная функция моноцитов/макрофагов (Борисова и др., 1999).
Популярной остаётся точка зрения, что ключевым в иммуностарении является процесс, регулирующий инволюцию и атрофию тимуса, вскоре после начала которых содержание тимусного гормона в сыворотке начинает уменьшаться с возрастом. Такое мнение косвенно подтверждается интересным экспериментом, проведённым Куликовым с соавт. (1997), с использованием разработанного ими способа аллотрансплантации тимуса и костного мозга в иммунопривилегированные области организма. Пересадив ткани от молодых крыс стареющим животным, они показали замедление возрастной инволюции тимуса, увеличение в нём и костном мозге митотического индекса, а также активацию синтеза макромолекул в 1.5-2.1 раза в головном мозге, печени и селезёнке за счёт повышения активности ДНК-зависимой РНК-полимеразы.
Изучение возрастных изменений первичных и вторичных органов имму-нитета у мышей C57BL/6 в возрасте от 1 дня до 1 года показало, что до определённого момента жизни и роста животных относительный вес тимуса, селезёнки и подмышечных лимфоузлов у них снижается, однако, позже инволюции подвергается только тимус. При этом отмечается корреляция между снижением массы тимуса и уменьшением лимфоидных клеток в головном мозге (Dominguez-Gerpe, Rey-Mendez, 1998).
Каковы же механизмы увеличения числа дефектных лимфоцитов при возрастной инволюции органов иммунной системы? Почему снижаются пролиферативная активность, эффекторная и хелперная функции Т-клеток, нарушаются супрессорная их функция, соотношение различных видов Т-клеток? Чёткого ответа на эти вопросы пока нет.
Полагая, что старение стволовых клеток иммунной системы, самих лимфоцитов, как и всех других типов клеток, начинается с деградации митохондрий и создания условий для пероксидативного стресса (см. п. 1.3), обсудим с этих позиций возможные механизмы возрастного Т-клеточного иммунодефицита.Как известно, повреждение мембран – наиболее ранний и основной воз-растной дефект лимфоцитов. Для старых этих клеток характерно увеличение насыщенности и микровязкости липидов (Садовникова, 1986). Одним из результатов модификации липидного состава мембран лимфоцитов может быть подавление клеточных реакций иммунитета, а уменьшение содержания ненасыщенных фосфолипидов и обогащение мембран холестерином снижает цитолитическую активность эффекторных Т-клеток (Schlager et al., 1983). В этом отношении примечательны материалы, полученные Черкашиным и соавт. (1988) в эксперименте. Изучая у 83 лиц пожилого и старческого возраста состояние ПОЛ в цельной крови, плазме, эритроцитах и лимфоцитах во взаи-мосвязи с показателями системного иммунитета, они установили следующее. Снижение показателей клеточного иммунитета совпадает с увеличением концентрации MDA, диеновых конъюгатов во всех исследованных средах (по сравнению с контролем – 32 здоровых человека среднего возраста) на фоне снижения антиоксидантной активности. Наиболее четко показатели клеточного иммунитета коррелируют с уровнем продуктов ПОЛ в лимфоцитах. Содер-жание в них MDA увеличилось на 40 %, диеновых конъюгатов – на 31.7 %, а количество GSH уменьшилось на 35 %. Авторы, естественно, связали эти данные с дестабилизирующим действием продуктов ПОЛ на функцию клеточных мембран, угнетением ими клеточной реакции иммунитета.
Липидной пероксидации подвергаются и внутренние мембраны лимфоцитов, в том числе однослойная липопротеидная мембрана лизосом. На это указывает высокая активность катепсинов в лимфоцитах пожилых людей после окислительного стресса, хотя исходная активность этих протеолитических ферментов у пожилых и молодых была одинаковой (Garcia-Arumi et al., 1998).
Вероятно, при окислительном стрессе высвобождение катепсинов из лизосом лимфоцитов происходит у пожилых (по сравнению с молодыми) сравнительно легко, ввиду наличия в их крови и в самих форменных её элементах проок-сидантных условий, т.е. вследствие «подготовленности» и приближенности мембран лимфоцитов к состоянию избыточной пероксидации. Некоторое усиление такого состояния с помощью окислительного стресса должно приводить к дестабилизации мембраны лизосом и соответственно к заметному повышению уровня и активности эндопептидаз, в частности катепсинов, в гиалоплазме. Вероятность же подобного исхода в лимфоцитах, взятых у молодых людей, должна быть значительно более низкой.Принципиально повышенный уровень ПОЛ в мембранах активированных эффекторных клеток (естественных Т-киллеров, микро- и макрофагов) заложен уже самим существованием в них природного механизма «кислородного взрыва» в качестве мощного оружия обезвреживания антигенов. Правда, и соб-ственная безопасность их от возможного окислительного повреждения представлена соответственно более надёжной антиоксидантной системой. Однако в случае «срыва» последней по тем или иным причинам указанное повреждение неизбежно. При старении эффекторных клеток и их предшественников серь-ёзным таким срывом является выход в первую очередь из строя основной, «митохондриальной» ступени антиоксидантной системы.
Активации пероксидативных процессов в плазматической мембране того же лимфоцита или макрофага способствует также сам провоцирующий ПОЛ акт взаимодействия их с антигеном, лектином, медиатором, с другой клеткой, приводящий к разрыхлению структуры липидной матрицы и модификации мембраны. В процессе ПОЛ плазматическая мембрана теряет легкоокисляемые ненасыщенные фосфолипиды фосфатидилсерин и фосфатидилинозит, а трудноокисляемые сфингомиелин и фосфатидилхолин, наоборот, накапливаются в ней (Садовникова, 1986). Если в здоровых физиологических условиях ПОЛ выполняет определённые полезные для клети функции (участие в стимуляции митогенеза, обновление липидов мембран и др.), то при возрастном повышении в ней дисбаланса ∆ (ПО – АО) усиленное переокисление оказывает уже негативное действие.
Прежде всего, падает чувствительность рецепторов и ферментов, встроенных в плазматическую мембрану.Как известно, ключевые ферменты протеинкиназа С (PKC) и аденилатциклаза в составе двух разных фундаментальных механизмов формирования и передачи универсальных сигналов с плазматической мембраны во внутриклеточные структуры являются соответственно фосфатидилсерин- и фосфатидил-инозит-зависимыми. Активация Na+,K+-АТРазы и, вероятно, ещё ряда других ферментов в клеточной мембране зависит от фосфатидилинозитного микроокружения (Abdel-Latif, 1983). Кроме того, некоторые ферменты – щелочная фосфатаза (Noda et al., 1987), орнитиндекарбоксилаза (Mustelin et al., 1987) заякорены в плазматической мембране с помощью фосфоинозитидов. По нашему мнению, в названных случаях легкоокисляемые фосфолипиды выполняют в норме регуляторную функцию, «выключая» или «включая» соответствующие зависимые от них ферменты при повышении ПОЛ мембраны в физио-логически допустимых ещё пределах. Если иметь в виду аденилатциклазу, то такое выключение её в эффекторных клетках должно снижать уровни сАМР и сАМР-зависимого митохондриального дыхания и, следовательно, создавать внутриклеточную гипероксию. Этот механизм, по-видимому, задействован природой для реализации циклически повторяющегося «окислительного взрыва» при активации Т-киллеров и макрофагов в качестве одного из основных защитных средств в системе клеточного иммунитета.
При устойчивом пероксидативном стрессе как движущей силы старения, воздействующей в том числе на фосфолипидные связи с мембраной и микро-окружение указанных выше ферментов, нормальное функционирование последних будет, очевидно, непрерывно нарушаться и в значительной степени блокироваться. Например, активность PKC в этой ситуации должна быть постоянно низкой, а не периодически, как в норме. В таком состоянии угнета-ются клеточные реакции иммунитета, снижаются уровни пролиферативной и естественной цитотоксической активности эффекторных клеток. По этим же «кислородно-перекисным» причинам стареющие клетки иммунной системы могут подвергаться апоптозу (см.
п.7.1.1) и приводить, в частности, к возрастным лимфопении и ослаблению Т-клеточных функций. Так, было сообщено (Aggarwal, Gupta, 1998), что у здоровых людей 65-95 лет по сравнению с молодыми (20-29 лет) на CD4+ и CD8+ Т-лимфоцитах периферической крови повышена экспрессия молекул Fas и Fas-лиганда. В этих же лимфоцитах снижена экспрессия антиапоптозного белка bcl-2 и увеличена экспрессия проапоптозного белка bax. У пожилых людей указанные клетки были более чувствительны к индукции апоптоза in vitro под действием антител против антигена Fas. Высокая чувствительность Т-лимфоцитов показана и к апоптозу, вызыва-емому TNF-α, и с этим фактом связывается дефицит этих клеток у лиц пожи-лого возраста (Aggarwal et al., 1999).Небезынтересны также материалы обзора о возрастных изменениях про-цессов преобразования сигналов в Т-клетках, которые связываются с ослабле-нием активности или экспрессии молекул, выполняющих соответствующие функции Hirokawa, 1999). Считают, например, что ослабление активности может быть сопряжено с нарушением строения указанных молекул в результате окислительного стресса. При взаимодействии Т-клеток с антигенами имеет место возрастзависимое изменение фосфорилирования белков тирозинкина-зами. Это, в свою очередь, ведёт к изменению содержания разных фосфопротеидов, причастных к преобразованию сигналов.
По некоторым данным, с возрастом в лимфоцитах человека повышается частота мутаций. Для выявления причин этого сдвига проведён анализ связи его с уровнем АФК, состоянием антиоксидантной системы и активностью репарационных систем. Результаты исследований показали, что с возрастом падает эффективность репарации ДНК. Это, скорее всего, и приводит к росту частоты мутаций, но снижения активности антиоксидантной защиты не происходит (Barnett, King, 1995). Данная информация согласуется также с материалами, по которым в лимфоцитах старых организмов устанавливается окислительный стресс, обусловленный недостаточностью митохондриального дыхания. Показано, в частности, что лимфоциты селезёнки старых мышей обладали более низкими скоростями дыхания и пониженным мембранным потенциалом митохондрий, чем лимфоциты молодых животных. При инкубации тех и других с липофильной краской (указывается её тип) первые были чувствительнее вторых к ингибированию ею транспорта электронов по дыхательной цепи (Rottenberg, Wu, 1997).
Названные выше изменения происходят, по-видимому, во всех типах Т-клеток, в том числе и в специализированных Т-регулирующих, существование которых предсказывается теоретическим моделированием закономерностей функционирования системы межклеточной регуляции клеточного роста разнотипных соматических тканей. Согласно новой иммунной теории старения (Донцов, 1998), такие Т-регуляторы реагируют «в сингенной смешанной культуре лимфоцитов» на собственные растущие клетки организма. Снижение с возрастом функции таких специализированных Т-клеток может, считает автор, являться центральным механизмом старения самообновляющихся соматических тканей организма, определять снижение ростового потенциала последних при старении.
1.5.4. Наряду с иммунокомпетентными клетками возрастным изменениям, как отмечалось выше, подвергаются органы, выполняющие иммунобиологическую функцию. К числу таких органов относится селезёнка, которая продуцирует лимфоциты и антитела, воспроизводит генерацию фагоцитирующих клеток. В частности, в клетках селезёнки старых мышей уменьшаются активности PKC и сАМР-зависимой протеинкиназы (PKA), причём эти возрастные изменения более выражены в мембранной фракции клеток (Blumenthal, Malkinson, 1989). Данные факты сходны с упомянутыми выше для эффекторных клеток периферической крови. Кроме того, у старых мышей выявлено выраженное снижение уровня цитотоксической активности клеток селезёнки при сохранении уровня цитостатической активности (Ванько и др., 1988). Это может быть связано с тем, что первая из них определяется не просто созданием гипероксии, но и способностью фагоцитов и Т-киллеров продуцировать при этом АФК как конкретных участников «окислительного взрыва». Между тем, активность NADPH-оксидазы, ответственной за производство О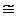 в плазматической мем-бране указанных клеток, с возрастом должна снижаться, поскольку активирующая её липидозависимая PKC сама инактивируется при устойчиво под-держиваемой гипероксии, но этих условий, по-видимому, достаточно для проявления цитостатического действия.
в плазматической мем-бране указанных клеток, с возрастом должна снижаться, поскольку активирующая её липидозависимая PKC сама инактивируется при устойчиво под-держиваемой гипероксии, но этих условий, по-видимому, достаточно для проявления цитостатического действия.
О протекании указанных негативных процессов в иммунокомпетентных клетках свидетельствуют различные данные, а частности следующие. Спектрофотометрическим методом с использованием цитохрома с определяли генерацию О моноцитами, выделенными из крови здоровых людей в возрасте 25-75 лет. Моноциты активировали форболовым эфиром (форболмиристатацетатом), стимулирующим NADPH-оксидазу. Оказалось, что указанная генерация ослабевала с возрастом, причём у женщин и мужчин соответственно на 70 и 45 % (Alvarez, Santa, 1996). У пожилых людей, стариков 80 лет и старше хемилюминесценция нейтрофилов, по уровню которой судили о дисфункции последних в части выделения активного О2, была значительно ниже, чем у взрослых здоровых лиц (Aoki, 1994).
Наибольшие же возрастные изменения происходят в другом органе иммун- ной системы – тимусе, через который проходят и где формируются Т-клетки. Эти изменения могут проявляться как в снижении гормональной активности тимуса и пролиферативных потенций Т-клеток, так и в нарушении их эффек-торных и иммунорегуляторных функций. В результате создаётся предраспо-ложение организма к инфекциям, злокачественным опухолям и аутоиммунным заболеваниям, а с недостаточностью тимусного гормона связывают наблюда-емый с возрастом дефект Т-клеток, незавершённость их дифференцировки, уменьшение числа эффекторных клеток (см. выше; Antonaci et al., 1996; Block, Ziesche, 1998; Ptak, Szczepanik, 1998). Так, по давним ещё сведениям (Reddy, Goh, 1979), число циркулирующих Т-клеток у 60-70-летних людей составляет только приблизительно 70 % уровня, характерного для молодых.
Любопытно, что гормоны тимуса оказывают также выраженное стабилизирующее действие на интенсивность ПОЛ в лимфоцитах периферической крови. Это было показано путём тимэктомии, которая вызывала в организме животных усиление переокисления фосфолипидов, накопление продуктов ПОЛ и изменение структурных компонентов биомембран. Введение тимэктомированным животным в ранние сроки тимозина приводило к восстановлению нормального уровня ПОЛ. Такое же влияние гормоны тимуса оказывали на интенсивность ПОЛ в субклеточных органеллах печени крыс (Мамутов, 1991). Очевидно, при возрастной инволюции тимуса его ПОЛ-ингибирующее действие должно ослабляться, способствуя повышению дисбаланса ∆ (ПО – АО) в клетках и тканях организма и, следовательно, их старению. В этом аспекте можно лишь отчасти согласиться с мнением Бернета (Burnet, 1970), что клетки тимуса и тимус-зависимые клетки определяют темп старения, а изменения в них ответственны за аутоиммунные и другие возрастные заболевания. Главным объективным фактором гетерохронного старения клеток, тканей и систем живого организма является всё же сама гипероксическая среда его обитания, в условиях которой одним из самых чувствительных органов постулируется тимус. Элиминируя первым, он, естественно, способствует деградации и всех других связанных с ним, но медленнее стареющих частей организма.
Относительно причин и механизма процесса инволюции и атрофии тимуса, начинающегося у человека и животных, как считают (George, Ritter, 1996), c момента рождения, можно сказать, что какие-либо принципиальные резуль-таты их исследования пока не получены. В процессе возрастной инволюции тимуса происходит морфологическая перестройка её сосудистого русла, в частности, уменьшение плотности микрососудов в паренхиме и капсуле. Эти данные выявлены при исследовании 104 тимусов людей в возрасте от новорожденных до 80 лет (Бабкина, 1996). Представляет интерес предположение о жаберном происхождении тимуса (Hirokawa, 1`980). Как известно, в ходе эволюции жабры у млекопитающих подверглись инволюции, и тот же в принципе процесс элиминации, возможно, отражён и унаследован их дальним «родственником» тимусом.
Конкретно же в основе механизма атрофии тимуса, предположительно, могут находиться следующие процессы. Прежде всего, мы распространяем идею «митохондриального» старения также на тимоциты и другие типы клеток в тимусе. Более того, полагаем, что этот процесс протекает в тимусе интен-сивнее, чем в других органах и тканях, и, по существу, является основным пусковым механизмом окислительной деградации или апоптоза его клеток. Может быть, процесс освобождения от митохондрий здесь осуществляется по «сценарию», сходному с таковым при созревании ретикулоцитов (молодых эритроцитов). В последних реализуется пероксидативный путь разрушения митохондрий с участием липооксигеназы (Rapoport, Schewe, 1986; Schnurr et al., 1996), который запрограммирован природой в качестве естественного этапа развития ретикулоцитов в зрелые эритроциты. В процессе адаптации живых организмов к гипероксическим условиям внешней среды эритроциты при-обрели способность накапливать О2 и насыщать им молекулы гемоглобина, обеспечивая тем самым клетки организма активным участником выгодного в энергетическом отношении процесса окислительного фосфорилирования. При наличии же множества конкурирующих за О2 митохондрий и соответственно низком рО2 в эритроцитах указанная крайне важная функция была бы невоз-можна или затруднена (Лю, Шайхутдинов, 1991).
Возвращаясь к липооксигеназе, отметим, что позднее было сформулировано более общее положение об участии её в программированной деградации разных внутриклеточных органелл в процессе дифференцировки ретикулоцитов и ряда других специализированных типов клеток (Van Leyen et al., 1998). Перед началом деградации органелл липооксигеназа интегрируется в их мемб-раны и увеличивает проницаемость последних. Тем самым она способствует высвобождению содержимого органелл, а их белки становятся доступными для действия протеаз. Однако применительно к деградации митохондрий в клетках тимуса данное представление, с нашей точки зрения, несостоятельно. Действи-тельно, в тимусных клетках складывается совсем иная ситуация: в отсутствии в них какого-либо О2-депонирующего устройства деградация митохондрий может привести лишь к апоптозу или окислительному разрушению этих клеток и тимуса в целом. Вероятно, поэтому инволюционирующий тимус содержит содержит многочисленные макрофаги, но малое количество тимоцитов (Hirokawa, 1980). Поскольку митохондрии слишком активны в образовании О и представляют опасность для клетки, то, по мнению Скулачева (1997), на митохондриальном уровне возможна реализация механизма, подобного апоптозу. На этот случай предложен даже термин «митоптоз». Не исключено, что такой механизм выбраковки митохондрий существует и в клетках тимуса. Результатом же его действия, по нашим представлениям, будет не защита этих клеток, а ускоренная их гибель по кислородно-перекисному механизму апоптоза (см. главу 7). В пользу важности феномена апоптоза для атрофии тимуса говорит тот факт, что у трансгенных мышей с дефектом апоптоза наступает ингибирование инволюции тимуса (см. Семенков, Артемьева, 1999).
Изложенные выше факты и положения о возможных причинах возникновения с возрастом Т-клеточного иммунодефицита обобщены в виде двух функциональных схем на рис.6.
Отмечая ослабление иммунной системы при старении человека и других животных, следует ещё раз подчеркнуть, что оно сопровождается такими же
 |  | ||||||
 | |||||||
| |||||||
нарушениями и иммунологическими сдвигами, которые характерны в случаях недоразвития у них Т-клеточного компонента указанной системы (Fernandez et al., 1980). С точки зрения кислородно-перекисной концепции, принципиально важными являются также данные о способности ингибиторов свободных радикалов, антиоксидантов и ингибиторов образования таких радикалов по липооксигеназному и циклооксигеназному путям подавлять митогенез, киллерную активность и фагоцитоз (Садовникова, 1986). Это свидетельствует, с одной стороны, о наличии в механизме указанных столь разных биологических феноменов общих моментов, а с другой, – о необходимости в соответствующих иммунокомпетентных клетках определённого, но разного уровня дисбаланса ∆ (ПО – АО). Свободные радикалы выполняют в этих процессах роль истинных вторичных посредников.
Обращает на себя внимание ещё один немаловажный факт, указывающий на избыточный апоптоз зрелых Т-лимфоцитов при иммунном старении. Активация мононуклеаров крови старых (> 80 лет) людей фиксированными на пластинке моноклональными антителами приводила к развитию межнуклеосомной деградации ДНК и снижению численности клеток в культуре. Эти сдвиги были менее заметными при активации клеток крови молодых (< 35 лет) при том, что уровень онкобелка bcl-2, считающегося ингибитором апоптоза, в старых и молодых клетках был одинаков в процессе их активации. Авторы (Phelouzat et al., 1996) заключили, что усиление апоптоза активированных Т-лимфоцитов – характерный признак иммунологического старения, первичную основу кото-рого, как и самого апоптоза (см. главу 7), составляет, по нашему мнению, устойчивый окислительный стресс.
В целом известные на сегодня иммунные теории старения не содержат, как считают некоторые исследователи (Никитин, 1982 и др.), главного критерия полноценности теорий онтогенеза – принципа первичности изменений, поэтому они могут иметь лишь определённое «вспомогательное» значение для понимания некоторых сторон процесса старения. Нам представляется, что этому принципу соответствует изложенное здесь «митохондриально-пероксида-тивное» старение клеток иммунной системы. Во всяком случае, в пользу такого механизма свидетельствуют уязвимость иммунокомпетентных клеток к физиологическим концентрациям радикалов в связи с возрастным уменьшением активности антиоксидантных ферментов (Садовникова, 1986) и, наоборот, стимуляция иммунологических реакций при старении ловушками свободных радикалов, антиоксидантами-геропротекторами (Walford, 1982).
Вопрос о старении иммунной системы намного, естественно, сложнее, чем тот, который в ограниченных рамках обсуждался выше. Действительно, как отмечает Бутенко (1998), возрастные изменения иммунитета связаны с наруше-ниями регуляции на уровне иммуно-нейро-эндокрийных взаимодействий, на уровне взаимодействий органов и клеточных популяций внутри самой иммун-ной системы и, наконец, на уровне внутриклеточных сигнальных механизмов. В этом плане многое предстоит ещё познать, но, в конечном счёте, все или большинство нарушений на указанных уровнях, как представляется нам, опре-деляются прежде всего внутриклеточными изменениями, изначально нося-щими пероксидативный характер.
1.5.5. Судя по публикациям,
Еще по теме 1.5.3. Нарушения в системе иммунитета при старении:
- Изменения систем при старении.
- 1 . 2 . 4 . Локальный иммунитет влагалища и его нарушения при хроническом рецидивирующем кандидозе гениталий и рецидивирующем бактери- альном вагинозе
- 1.5. Старение систем организма
- Роль нарушений иммунитета в возникновении опухолей
- 4.2.4. Основные алгоритмы нарушений в иммунной системе при ВИД.
- Паллиативный уход при нарушениях дыхательной системы
- Нарушение функций органов и систем при шоке
- Изменения в организме при старении
- Помощь при нарушениях в работе мочевыводящей системы
- 4.2 Синтез биотехнической системы диагностики микроциркуляторных нарушений при ревматических заболеваниях
- 1.6. Некоторые «парадоксальные» феномены при старении