1.5.1. жХРНРНЙЯХВЕЯЙХЕ ОПНДСЙРШ ЛЕРЮАНКХГЛЮ н2
СВЮЯРБСЧР Б ЯРЮПЕМХХ МЕ РНКЭЙН РЕУ ЙКЕРНЙ, РЙЮМЕИ Х ЯХЯРЕЛ, ЦДЕ НМХ НАПЮГСЧРЯЪ ОПХ ОНБШЬЕММНЛ СПНБМЕ ∆ (он √ юн). вЮЯРЭ РЮЙХУ ОПНДСЙРНБ БШДЕКЪЕРЯЪ ХГ ЩРХУ ЯРПСЙРСП Х ОНОЮДЮЕР Б ЯХЯРЕЛС ЙПНБННАПЮЫЕМХЪ, ЪБКЪЪЯЭ, ОН-БХДХЛНЛС, ЯСЫЕЯРБЕММШЛ ТЮЙРНПНЛ ОНБПЕФДЕМХЪ ЖХПЙСКХПСЧЫХУ Б МЕИ ТНПЛЕММШУ ЩКЕЛЕМРНБ ЙПНБХ.
нОПЕДЕК╦ММШИ БЙКЮД Б ЩРХ ЯДБХЦХ БМНЯЪР ОКЮГЛЮЛЕЛАПЮММШЕ НЙЯХДЮГШ ЯРЮПЕЧЫХУ ЙКЕРНЙ ПЮГКХВМНЦН РХОЮ, Б РНЛ ВХЯКЕ Х МЕ ЪБКЪЧЫХУЯЪ ОПНТЕЯЯХНМЮКЭМШЛХ ТЮЦНЖХ-РЮЛХ. оПНДСЖХПСЕЛШЕ ХЛХ ютй, Б ВЮЯРМНЯРХ н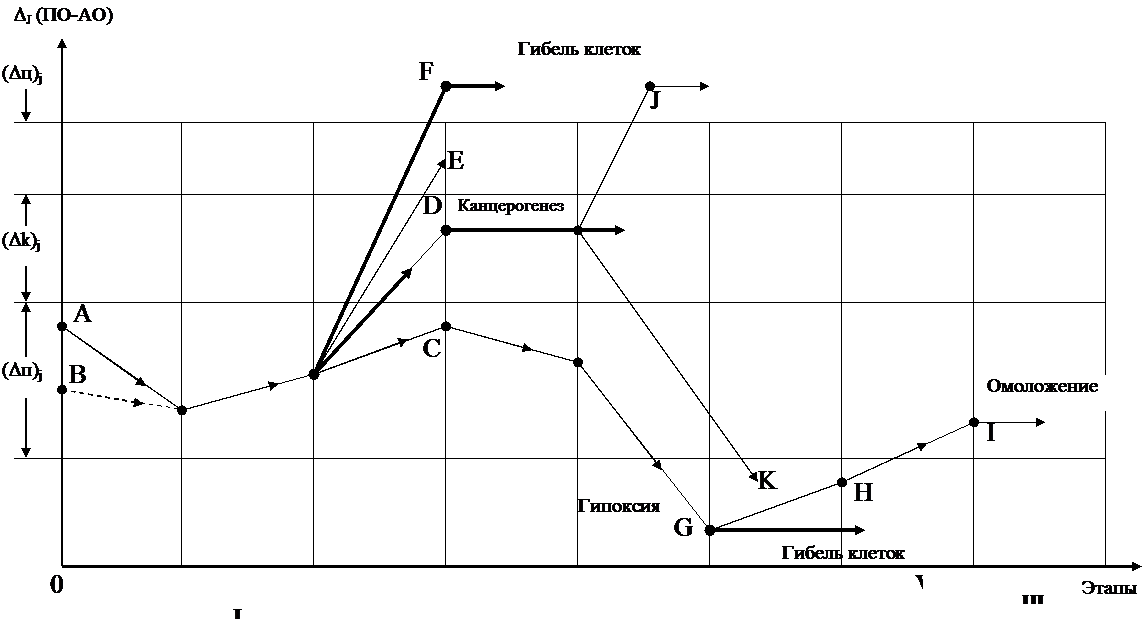 , ЛНЦСР ПЮЯОПНЯРПЮМЪРЭЯЪ МЮ ЯНЯЕДЯРБСЧЫХЕ ЙКЕРЙХ Х ОПНМХЙЮРЭ Б ЖХПЙСКХПСЧЫСЧ ЙПНБЭ, УНРЪ ЩРНЛС Х ОПЕОЪРЯРБСЕР ЙНЩМГХЛ Q Я ЮМРХЯРЮПХРЕКЭМНИ, ОПЕДОНКНФХРЕКЭМН, ЯОНЯНАМНЯРЭЧ (Morre et al., 2000). н РНЛ, ВРН Я БНГПЮЯРНЛ ОПННЙЯХДЮМРМЮЪ ВЮЯРЭ ДХЯАЮКЮМЯЮ ∆ (он √ юн) Б ЙПНБХ ОНБШЬЮЕРЯЪ, МН ЮМРХНЙЯХДЮМРМЮЪ, МЮОПНРХБ, ЯМХФЮЕРЯЪ, ЛНЦКН АШ ЯБХДЕРЕКЭЯРБНБЮРЭ СБЕКХВЕМХЕ Б МЕИ ЙНМЖЕМРПЮЖХХ ЯБНАНДМШУ ПЮДХЙЮКНБ Х ОЕПЕЙХЯЕИ. йНМЙПЕРМШЕ ДЮММШЕ МЮ ЩРНР ЯВ╦Р ОНКСВЕМШ ЙЮЙ Б ЩЙЯОЕПХЛЕМРЕ, РЮЙ Х Б ЙКХМХВЕЯЙХУ СЯКНБХЪУ.
, ЛНЦСР ПЮЯОПНЯРПЮМЪРЭЯЪ МЮ ЯНЯЕДЯРБСЧЫХЕ ЙКЕРЙХ Х ОПНМХЙЮРЭ Б ЖХПЙСКХПСЧЫСЧ ЙПНБЭ, УНРЪ ЩРНЛС Х ОПЕОЪРЯРБСЕР ЙНЩМГХЛ Q Я ЮМРХЯРЮПХРЕКЭМНИ, ОПЕДОНКНФХРЕКЭМН, ЯОНЯНАМНЯРЭЧ (Morre et al., 2000). н РНЛ, ВРН Я БНГПЮЯРНЛ ОПННЙЯХДЮМРМЮЪ ВЮЯРЭ ДХЯАЮКЮМЯЮ ∆ (он √ юн) Б ЙПНБХ ОНБШЬЮЕРЯЪ, МН ЮМРХНЙЯХДЮМРМЮЪ, МЮОПНРХБ, ЯМХФЮЕРЯЪ, ЛНЦКН АШ ЯБХДЕРЕКЭЯРБНБЮРЭ СБЕКХВЕМХЕ Б МЕИ ЙНМЖЕМРПЮЖХХ ЯБНАНДМШУ ПЮДХЙЮКНБ Х ОЕПЕЙХЯЕИ. йНМЙПЕРМШЕ ДЮММШЕ МЮ ЩРНР ЯВ╦Р ОНКСВЕМШ ЙЮЙ Б ЩЙЯОЕПХЛЕМРЕ, РЮЙ Х Б ЙКХМХВЕЯЙХУ СЯКНБХЪУ.оНЙЮГЮРЕКЭМШ, МЮОПХЛЕП, ПЕГСКЭРЮРШ ХЯЯКЕДНБЮМХИ ЯНДЕПФЮМХЪ ЦХДПНОЕПЕЙХЯЕИ КХОХДНБ Х ЮЙРХБМНЯРХ ЮМРХОЕПЕЙХЯМНЦН ТЕПЛЕМРЮ GPX ЯШБНПНРЙХ ЙПНБХ ЙПШЯ КХМХХ Wistar Б БНГПЮЯРЕ 1, 3, 12, 24 Х 32 ЛЕЯ., ЯНДЕПФЮБЬХУЯЪ МЮ НАШВМНИ ДХЕРЕ (ЕЯРЕЯРБЕММНЕ ЯРЮПЕМХЕ), МЮ ЙЮКНПХИМН НЦПЮМХВЕММНЛ ПЮЖХНМЕ (ГЮЛЕДКЕММНЕ ЯРЮПЕМХЕ) Х МЮ ПЮЖХНМЕ Я ДНАЮБКЕМХЕЛ Б РЕВЕМХЕ 2 ЛЕЯ. РХПНЙЯХМЮ (СЯЙН-ПЕММНЕ ЯРЮПЕМХЕ). б ВЮЯРМНЯРХ, СЯРЮМНБКЕМН, ВРН ОПХ ЕЯРЕЯРБЕММНЛ ЯРЮПЕМХХ ФХБНРМШУ СПНБМХ ЦХДПНОЕПЕЙХЯЕИ Х GPX-ЮЙРХБМНЯРЭ ЙНППЕКХПНБЮМШ ЯРПНЦН НРПХЖЮРЕКЭМН; ОПХ ЛНДЕКХПНБЮМХХ ГЮЛЕДКЕММНЦН ЯРЮПЕМХЪ ЙПШЯ GPX-ЮЙРХБ-МНЯРЭ Б ЯШБНПНРЙЕ ЙПНБХ СБЕКХВХБЮКЮЯЭ, Ю ЯНДЕПФЮМХЕ ОЕПЕЙХЯЕИ КХОХДНБ ЯМХФЮКНЯЭ; ЙНМЖЕМРПЮЖХЪ ЦХДПНОЕПЕЙХЯЕИ ОПХ ЛНДЕКХПНБЮМХХ СЯЙНПЕММНЦН ЯРЮПЕМХЪ БНГПЮЯРЮКЮ С ФХБНРМШУ БЯЕУ БНГПЮЯРМШУ ЦПСОО, ОПХВ╦Л ЯМХФЕМХЕ ЮЙРХБМНЯРХ GPX Х ОНБШЬЕМХЕ ЯНДЕПФЮМХЪ ОЕПЕЙХЯЕИ КХОХДНБ Б ЯШБНПНРЙЕ 24-ЛЕЯЪВМШУ ФХБНРМШУ АШКХ ЙЮВЕЯРБЕММН ЯУНДМШ Я ОПНХЯУНДЪЫХЛХ Б МНПЛЕ ОПХ ДЮКЭМЕИЬЕЛ СБЕКХВЕМХХ БНГПЮЯРЮ ЙПШЯ ДН 32 ЛЕЯ.
б ЖЕКНЛ, ХЯУНДЪ ХГ УЮПЮЙРЕПЮ ХГЛЕМЕМХЪ СЙЮГЮММШУ ОНЙЮГЮРЕКЕИ ОПХ ПЮГМШУ ЯЙНПНЯРЪУ ЯРЮПЕМХЪ ОНДНОШРМШУ ФХБНРМШУ, ЮБРНПШ ДЮММНЦН ХЯЯКЕДНБЮМХЪ (мХЙХРВЕМЙН, кЕЛЕЬЙН, 1997) ОНДРБЕПФДЮЧР ЙНМЖЕОЖХЧ Н ЙКЧВЕБНИ ПНКХ ЯБНАНДМНПЮДХЙЮКЭМНЦН НЙХЯКЕМХЪ КХОХДНБ Б ЛЕУЮМХГЛЕ ЯРЮПЕМХЪ.йПНЛЕ ОПЪЛШУ ТЮЙРНБ, ЙНЯБЕММН МЮ СЯХКЕМХЕ ОЕПНЙЯХДЮРХБМШУ ОПНЖЕЯЯНБ СЙЮГШБЮЕР БШЯНЙХИ СПНБЕМЭ УЕЛХКЧЛХМЕЯЖЕМЖХХ ЯШБНПНРЙХ ЙПНБХ С ЯРЮПШУ ФХБНРМШУ Х ОНФХКШУ КЧДЕИ ОН ЯПЮБМЕМХЧ Я РЮЙНБШЛ С ХМДХБХДССЛНБ ЯПЕДМЕЦН БНГПЮЯРЮ (аЮПЮАНИ Х ДП., 1988). яКЕДСЕР РЮЙФЕ ОПХМЪРЭ БН БМХЛЮМХЕ, ВРН Б ОКЮГЛЕ ЙПНБХ КЧДЕИ ЯРЮПВЕЯЙНЦН БНГПЮЯРЮ (>60 КЕР) ГМЮВХРЕКЭМН ЯМХФЕМН ЯНДЕПФЮМХЕ GSH Х ОНБШЬЕМН ЯНДЕПФЮМХЕ GSSG НРМНЯХРЕКЭМН ХУ СПНБМЪ С ЛНКНДШУ ДНМНПНБ (Samiec et al., 1998), Ю ЙНМЖЕМРПЮЖХЪ F2-ХГНОПНЯРЮМНБ Б ОКЮГЛЕ ЙПНБХ ЛНКНДШУ ЙПШЯ Б ~20 ПЮГ МХФЕ, ВЕЛ Б ОКЮГЛЕ ЯРЮПШУ ФХБНРМШУ, Р. Е. МЮКХЖН ОПХГМЮЙХ БШПЮФЕММНЦН ОПХ ЯРЮПЕМХХ НЙХЯКХРЕКЭМНЦН ЯРПЕЯЯЮ (Roberts et al., 2001). оПХЛЕВЮРЕКЕМ Х ДПСЦНИ ТЮЙР: ЯОНЯНАМНЯРЭ ОКЮГЛШ ЙПНБХ ВЕКНБЕЙЮ, ЙЮЙ ОНЙЮГЮК НАЫХИ ЮМРХНЙЯХДЮМРМШИ ЮМЮКХГ ЙПНБХ ДНАПНБНКЭЖЕБ Б БНГПЮЯРЕ 30-80 КЕР, ОПНРХБНЯРНЪРЭ ХНМХГХПСЧЫЕИ ПЮДХЮЖХХ ЯБЪГЮМЮ Я БНГПЮЯРНЛ НАПЮРМНИ ГЮ-БХЯХЛНЯРЭЧ (Lenton et al., 1999).
пЮЯЯЛНРПХЛ, ОПЕФДЕ БЯЕЦН, ЛЕУЮМХГЛ БНГПЮЯРМНЦН ХГЛЕМЕМХЪ ЩПХРПНЖХРНБ Б ЯБЪГХ Я БНГМХЙМНБЕМХЕЛ Х ОНДДЕПФЮМХЕЛ Б МХУ ОЕПНЙЯХДЮРХБМНЦН ЯНЯРНЪМХЪ (ЯЛ. ПХЯ. 5). хЯЯКЕДНБЮМХЪ МЮ ЛНКЕЙСКЪПМНЛ СПНБМЕ ОНЙЮГЮРЕКЕИ онк Х ЮМРХНЙЯХДЮМРМНИ ЯХЯРЕЛШ Б ЩПХРПНЖХРЮПМШУ ЛЕЛАПЮМЮУ КЧДЕИ ОНФХКНЦН Х ЯРЮПВЕЯЙНЦН БНГПЮЯРЮ ОНЙЮГЮКХ, ВРН ЯНДЕПФЮМХЕ ЦХДПНОЕПЕЙХЯЕИ КХОХДНБ Х MDA Б ЩРХУ ЛЕЛАПЮМЮУ ОНБШЬЕМН ЯННРБЕРЯРБЕММН МЮ 9.1 Х 46 % ОН ЯПЮБМЕМХЧ Я РЮЙХЛХ ФЕ ОНЙЮГЮРЕКЪЛХ С КХЖ ЛНКНДНЦН Х ЯПЕДМЕЦН БНГПЮЯРЮ (еУЮКНБ, яНАНКЕБ, 1988). щРХ ДЮММШЕ ОНДРБЕПФДЮЧР ЮЙРХБЮЖХЧ ОПНЖЕЯЯНБ онк Б ЩПХРПНЖХРЮУ ОПХ ЯРЮПЕМХХ НПЦЮМХГЛЮ, ЙНППЕКХПСЧЫСЧ Я ПНЯРНЛ ОПНДСЙРНБ онк Х ЯМХФЕМХЕЛ ЮМРХНЙЯХДЮМРМНИ ЮЙРХБМНЯРХ ЙПНБХ. юБРНПШ ЩРНЦН ХЯЯКЕДНБЮМХЪ НРЛЕВЮЧР РЮЙФЕ, ВРН ╚ЙЮВЕЯРБЕММНЕ ЯНЯРНЪМХЕ ЩПХРПНЖХРЮПМНИ ЛЕЛАПЮМШ ЙНЯБЕММН НРПЮФЮЕР ТСМЙЖХНМЮКЭМСЧ ОНКМНЖЕММНЯРЭ ЙКЕРНВМШУ ЛЕЛАПЮМ НПЦЮМНБ Х ЯХЯРЕЛ╩.
мЕЯЙНКЭЙН ПЮМЭЬЕ Н ГМЮВХРЕКЭМНЛ СБЕКХВЕМХХ Я БНГПЮЯРНЛ ЙНМЖЕМРПЮЖХХ MDA Б ЩПХРПНЖХРЮУ ВЕКНБЕЙЮ АШКН ЯННАЫЕМН Б ДПСЦНИ ПЮАНРЕ (Jozwiak, Jasnowska, 1985), ЦДЕ ДНМНПЮЛХ АШКХ КЧДХ Б БНГПЮЯРЕ НР 4-У ДН 80-РХ КЕР.
мН ГДЕЯЭ ГЮТХЙЯХПНБЮМ РЮЙФЕ Х ПНЯР ЮЙРХБМНЯРХ ЮМРХОЕПЕЙХЯМНЦН ТЕПЛЕМРЮ GPX, ВРН, БЕПНЪРМН, НРПЮФЮЕР МЕЙНРНПСЧ ЮДЮОРЮЖХЧ ЩПХРПНЖХРНБ Й ОНБШЬЕММНЛС СПНБМЧ онк ОПХ НАЫЕЛ БЯ╦-РЮЙХ ЯМХФЕМХХ ЩТТЕЙРХБМНЯРХ ХУ ЮМРХНЙЯХДЮМРМНИ ЯХЯРЕЛШ. тЮЙРШ, ОНДНАМШЕ СЙЮГЮММШЛ, ЯВХРЮЧРЯЪ СФЕ НВЕБХДМШЛХ, Х БПЪД КХ МСФДЮЧРЯЪ Б ОЕПЕОПНБЕПЙЕ. рЕЛ МЕ ЛЕМЕЕ, ХГПЕДЙЮ ОНДРБЕПФДЕМХЕ ХГБЕЯРМНЦН ОПНДНКФЮЕРЯЪ. рЮЙ, юМДН Я ЯНЮБР. (Ando et al., 1995) ОНЙЮГЮК МЮЙНОКЕМХЕ ЦХДПНОЕПЕЙХЯЕИ ТНЯТЮРХДХКУНКХМЮ Х ТНЯТЮРХДХКЩРЮМНКЮЛХМЮ Б ОПНЖЕЯЯЕ ЯРЮПЕМХЪ ЩПХРПНЖХРНБ ВЕКНБЕЙЮ Б ЖХПЙСКЪЖХХ. яБНАНДМШЕ Х ЯБЪГЮММШЕ ТНПЛШ ЛЮКНМНБНЦН, 4-ЦХДПНЙЯХ- Х ДПСЦХУ ЮКЭДЕЦХДНБ РЮЙФЕ ОПЕНАКЮДЮКХ Б ЯРЮПШУ ЩПХРПНЖХРЮУ ОН ЯПЮБМЕМХЧ Я ЛНКНДШЛХ. щРХ ХГЛЕМЕМХЪ ПЮЯЯЛЮРПХБЮЧРЯЪ ЙЮЙ ОПХГМЮЙХ ЯРЮПЕМХЪ ЩПХРПНЖХРНБ Б ЖХПЙСКЪЖХХ. б ЩПХРПНЖХРЮУ КЧДЕИ 20-89-РХ КЕР Я БНГПЮЯРНЛ ЯМХФЮЧРЯЪ ЮЙРХБМНЯРХ Cu,Zn-SOD Х ЦКСРЮРХНМПЕДСЙРЮГШ, ЮЙРХБМНЯРХ ФЕ GPX Х ЙЮРЮКЮГШ МЕ ХГЛЕМЪЧРЯЪ (Andersen et al., 1997).аХЯКНИМЮЪ ОКЮГЛЮРХВЕЯЙЮЪ ЛЕЛАПЮМЮ ЙКЕРНЙ, ЙЮЙ ХГБЕЯРМН, ЮЯХЛЛЕРПХВМЮ: БН БМСРПЕММЕЛ ЛНМНЯКНЕ Е╦ ОПЕНАКЮДЮЧР КЕЦЙННЙХЯКЪЕЛШЕ ТНЯТНКХОХДШ ОН ЯПЮБМЕМХЧ Я ХУ ЯНДЕПФЮМХЕЛ БН БМЕЬМЕЛ ЛНМНЯКНЕ. оН ЩРНИ ОПХВХМЕ ОКЮГЛЮРХВЕЯЙЮЪ ЛЕЛАПЮМЮ АНКЕЕ ВСБЯРБХРЕКЭМЮ Й ПЮГБХРХЧ онк ХЛЕММН ЯН ЯРНПНМШ БМСРПЕММЕЦН ЛНМНЯКНЪ. оПХЛЕМХРЕКЭМН Й ЩПХРПНЖХРЮЛ СЙЮГЮММНЕ НАЯРНЪРЕКЭ-ЯРБН ЛНФЕР АШРЭ ОПХВЮЯРМШЛ Й СЦМЕРЕМХЧ Б МХУ ЩМЕПЦЕРХВЕЯЙНЦН НАЛЕМЮ, Р.Е. Й ОЮДЕМХЧ ХМРЕМЯХБМНЯРХ ЦКХЙНКХГЮ, ОПХ ЯРЮПЕМХХ НПЦЮМХГЛЮ. тНПЛЮКЭМН РЮЙЮЪ БНГЛНФМНЯРЭ ЯБЪГШБЮЕРЯЪ МЮЛХ Я ЦХОНРЕРХВЕЯЙНИ ЯРПСЙРСПНИ ЙНЛОКЕЙЯЮ
 | ||||
| ||||
н ОЮДЕМХХ ЯЙНПНЯРХ ЦКХЙНКХГЮ Б ЩПХРПНЖХРЮУ ВЕКНБЕЙЮ ОПХ ЯРЮПЕМХХ ЯННАЫХКХ, Б ВЮЯРМНЯРХ, йНПЙСЬЙН Я ЯНЮБР. (1989). нМХ ФЕ НРЛЕВЮЧР ОНБШЬЕМХЕ БЕКХВХМШ Пм БМСРПХ ЩПХРПНЖХРНБ Х НАСЯКНБКЕММНЕ ХЛ ЯМХФЕМХЕ ЯПНДЯРБЮ н2 Й ЦЕЛНЦКНАХМС. оПХ РЮЙНЛ ХЯУНДЕ ПНКЭ ЦЕЛНЦКНАХМЮ ЙЮЙ ╚СЯРПНИЯРБЮ╩, БПЕЛЕММН ДЕОНМХПСЧЫЕЦН н2, ОЮДЮЕР, ВРН, ЕЯРЕЯРБЕММН, ОПХБНДХР Й БНГПЮЯРЮМХЧ Пн2 Б ЩПХРПНЖХРЮУ. рЮЙ БНГМХЙЮЧР СЯКНБХЪ Б БХДЕ ОНКНФХРЕКЭМНИ НАПЮРМНИ ЯБЪГХ ДКЪ ОНДДЕПФЮМХЪ Б МХУ ХГАШРНВМНИ ОЕПНЙЯХДЮЖХХ, ХЯРНЫЕМХЪ ЮМРХНЙЯХДЮМРМНИ ЯХЯРЕЛШ ГЮЫХРШ Х ЯМХФЕМХЪ Б ЙПНБХ ЙНКХВЕЯРБЮ ОНКМНЖЕММШУ ЩПХРПНЖХРНБ. оНЯКЕДМЕЕ, Б ЯБНЧ НВЕПЕДЭ, БЛЕЯРЕ Я ДЕТЕЙРНЛ Б ЯБЪГШБЮМХХ н2 ЦЕЛНЦКНАХМНЛ СЛЕМЭЬЮЕР ЙНЩТТХЖХЕМР ХЯОНКЭГНБЮМХЪ н2 Б КЕЦЙХУ Х НАСЯКНБКХБЮЕР ЯПЮБМХРЕКЭМН БШЯНЙНЕ Пн2 Б ЮКЭБЕНКЪПМНЛ БНГДСУЕ (кЮСЩП, яЕПЕДЕМЙН, 1982).
б МЮГБЮММШУ БШЬЕ СЯКНБХЪУ ДНОНКМХРЕКЭМШЛ ТЮЙРНПНЛ, ЯОНЯНАЯРБСЧЫХЛ БНГМХЙМНБЕМХЧ НЙХЯКХРЕКЭМНЦН ЯРПЕЯЯЮ Б ЩПХРПНЖХРЮУ ЛКЕЙНОХРЮЧЫХУ, ЪБКЪЧРЯЪ ОНЯКЕДЯРБХЪ ЩКХЛХМЮЖХХ ЛХРНУНМДПХИ ОПХ ЯНГПЕБЮМХХ ЙКЕРНЙ ЩПХРПНХДМНЦН ПЪДЮ. хЯВЕГМНБЕМХЕ Б МХУ ЛХРНУНМДПХИ √ МЮХАНКЕЕ ЮЙРХБМШУ ОНРПЕАХРЕКЕИ н2, МЕЯНЛМЕММН, ТЮЙР МЕ ЯКСВЮИМШИ. нМ ╚НПЦЮМХГНБЮМ╩ ОПХПНДНИ ХКХ ОПНЯРН ХГ-ГЮ МЕМЮДНАМНЯРХ ХУ Б ЙНМЙПЕРМШУ СЯКНБХЪУ ЯСЫЕЯРБНБЮМХЪ ЙКЕРЙХ, ХКХ ФЕ ДКЪ ЯНГДЮМХЪ ЯХРСЮЖХХ, НАЕЯОЕВХБЮЧЫЕИ ПЕЬЕМХЕ ЙЮЙХУ-РН ДПСЦХУ ЯНБЕПЬЕММН МНБШУ ГЮДЮВ. рЮЙНБШЛХ ЯРЮКХ МЕНАУНДХЛНЯРЭ СЯРПЮМЕМХЪ ЛХРНУНМДПХИ ЙЮЙ ЙНМЙСПЕМРНБ ЦЕЛНЦКНАХМЮ ГЮ н2, МЮЯШЫЕМХЕ ЛНКЕЙСК ЦЕЛНЦКНАХМЮ ЩРХЛ н2, ЙНРНПШИ Б МНБНИ ЯХРСЮЖХХ РЕОЕПЭ РНКЭЙН ДЕОНМХПСЕРЯЪ, Ю МЕ ПЮЯУНДСЕРЯЪ. хМРЕПЕЯМН, ВРН ОПХ ЩКХЛХМЮЖХХ ЩПХРПНЖХРЮПМШУ ЛХРНУНМДПХИ ХГ МХУ Б ЖХРНГНКЭ, ЙЮЙ АШКН ОНЙЮГЮМН МЮ ОПХЛЕПЕ ЯХПХИЯЙНЦН УНЛЪВЙЮ, МЕ БШДЕКЪЕРЯЪ ЖХРНУПНЛ Я (Takano-Ohmuro et al., 1998), ВРН НАШВМН ОПНХЯУНДХР ОПХ ЮОНОРНГЕ ДПСЦХУ РХОНБ ЙКЕРНЙ (ЯЛ.
О.7.1.10).рЮЙХЛ НАПЮГНЛ, ОПХ ЯРЮПЕМХХ ЯЙКЮДШБЮЧРЯЪ СЯКНБХЪ ДКЪ СЯРНИВХБНЦН ОНДДЕПФЮМХЪ Б ЩПХРПНЖХРЮУ ЦХОЕПНЙЯХХ Х ОНЯКЕДСЧЫЕИ НЙХЯКХРЕКЭМНИ ХУ ДЕЯРПСЙЖХХ. б ХГКНФЕММНЛ ЛЕУЮМХГЛЕ МЕЙНРНПШЕ ТЮЙРШ ОНЙЮ МЕ МЮУНДЪР НРПЮФЕМХЕ. яПЕДХ МХУ, МЮОПХЛЕП, РЮЙХЕ ДЮММШЕ (Ramachandran, Abraham, 1989). оПХ ЯРЮПЕМХХ ЩПХРПНЖХРНБ ВЕКНБЕЙЮ ОПНРЕХМЙХМЮГЮ я ОЕПЕУНДХР ХГ ЖХРНГНКЪ Б ЛЕЛАПЮММСЧ ТПЮЙЖХЧ Х ОНБШЬЮЕР СПНБЕМЭ ГЮБХЯХЛНЦН НР МЕ╦ ТНЯТНПХКХПН-БЮМХЪ ЛЕЛАПЮММШУ АЕКЙНБ. оПХВХМШ, ЯЛШЯК Х НРМНЬЕМХЕ ЩРХУ ОПНЖЕЯЯНБ Й ЯРЮПЕМХЧ ЩПХРПНЖХРНБ РПЕАСЧР ЕЫ╦ НЯЛШЯКЕМХЪ.
яЙКЮДШБЮЧЫХЕЯЪ Б ЩПХРПНЖХРЮУ СЯКНБХЪ ЦХОЕПНЙЯХХ Х ОЕПНЙЯХДЮЖХХ НРБЕРЯРБЕММШ МЕ РНКЭЙН ГЮ ДЕЯРПСЙРХБМШЕ ОПНЖЕЯЯШ Б ЯЮЛХУ ЩРХУ ЙКЕРЙЮУ. мЕЦЮРХБМШЕ ОНЯКЕДЯРБХЪ РЮЙНЦН ХУ ЯНЯРНЪМХЪ ОПЕДЯРЮБКЪЧРЯЪ АНКЕЕ ЛЮЯЬРЮА-МШЛХ, ╚БЯЕНПЦЮМХГЛЕММШЛХ╩ Я СВ╦РНЛ РНЦН, ВРН ОНД БКХЪМХЕЛ КХОНОЕПЕЙХЯЕИ ХГ ЛНКЕЙСК ЦЕЛНЦКНАХМЮ, ЙНКХВЕЯРБН ЙНРНПШУ Б НДМНЛ РНКЭЙН ЩПХРПНЖХРЕ ДНЯ-РХЦЮЕР ОПХЛЕПМН 265╥106, ЛНЦСР Б ЛЮЯЯНБНЛ ОНПЪДЙЕ БШЯБНАНФДЮРЭЯЪ ЙНЛОКЕ-ЙЯШ ФЕКЕГЮ. оНЯКЕДМХЕ, ПЮЯОПНЯРПЮМЪЪЯЭ ОН БЯЕЛС ЯРЮПЕЧЫЕЛС НПЦЮМХГЛС Х ОПНМХЙЮЪ Б ДПСЦХЕ ТНПЛЕММШЕ ЩКЕЛЕМРШ ЙПНБХ, Б ЙКЕРЙХ ПЮГКХВМШУ НПЦЮМНБ Х РЙЮМЕИ, ЯОНЯНАМШ ХМРЕМЯХТХЖХПНБЮРЭ Б МХУ ЦЕМЕПЮЖХЧ ЮЙРХБМШУ ЯБНАНДМШУ ПЮДХЙЮКНБ Х онк ЛЕЛАПЮМ. оПХ ЩРНЛ, Б ГЮБХЯХЛНЯРХ НР ОПЕДЬЕЯРБСЧЫЕЦН БМСРПХЙКЕРНВМНЦН ГМЮВЕМХЪ ДХЯАЮКЮМЯЮ ∆ (он √ юн), Б НДМХУ ЙКЕРЙЮУ БНГ-ПЮЯРЮЕР ЯЙНПНЯРЭ ОПНЯРН ЯРЮПЕМХЪ, Б ДПСЦХУ √ ЯЙНПНЯРЭ ХУ ГКНЙЮВЕЯРБЕММНИ РПЮМЯТНПЛЮЖХХ, Ю ЙЮЙХЕ-РН ЙКЕРЙХ АШЯРПЕЕ ОНДБЕПЦЮЧРЯЪ ЮОНОРНГС ХКХ НЙХЯ-КХРЕКЭМНЛС ЖХРНКХГС (ЯЛ. О. 1.6 Х ПХЯ. 7).
й ВХЯКС ТНПЛЕММШУ ЩКЕЛЕМРНБ ЙПНБХ, ОНДБЕПЦЮЧЫХУЯЪ БНГПЮЯРМНЛС ХГЛЕ-МЕМХЧ, НРМНЯЪРЯЪ Х РПНЛАНЖХРШ. я РПЮМЯОНПРНЛ н2 НМХ МЕ ЯБЪГЮМШ, МН ХЛЕЧР ОПЪЛНЕ НРМНЬЕМХЕ Й РПНЛАНЩЛАНКХВЕЯЙХЛ НЯКНФМЕМХЪЛ, ВЮЯРНРЮ ЙНРНПШУ ╚Б ВЕКНБЕВЕЯЙНИ ОНОСКЪЖХХ МЮПЮЯРЮЕР Я ЙЮФДШЛ ОПНФХРШЛ ДЕЯЪРХКЕРХЕЛ╩ (йНПЙСЬЙН, йНБЮКЕМЙН, 1988). с ОПЮЙРХВЕЯЙХ ГДНПНБШУ КЧДЕИ ОНФХКНЦН Х ЯРЮПВЕЯЙНЦН БНГПЮЯРЮ БШЪБКЪЕРЯЪ УПНМХВЕЯЙНЕ БМСРПХЯНЯСДХЯРНЕ ЛХЙПНЯБ╦ПРШ-БЮМХЕ ЙПНБХ, ВЮЯРНРЮ Х ХМРЕМЯХБМНЯРЭ ЙНРНПНЦН СБЕКХВХБЮЕРЯЪ Я БНГПЮЯРНЛ (яСЬЙН, 1994).
ю ХГСВЕМХЕЛ ЛНПТН-ТСМЙЖХНМЮКЭМНЦН ЯНЯРНЪМХЪ РПНЛАНЖХРНБ С ГДНПНБШУ КХЖ 18-79 КЕР СЯРЮМНБКЕМН БНГПЮЯРГЮБХЯХЛНЕ ОНБШЬЕМХЕ ЮЦПЕЦЮЖХНМ-МНИ ЮЙРХБМНЯРХ РПНЛАНЖХРНБ. нМН ОПНЪБКЪЕРЯЪ Б ПНЯРЕ ВХЯКЮ ЮЙРХБХПНБЮММШУ Х ДЕЦПЮМСКХПНБЮММШУ РПНЛАНЖХРНБ ОПХ ЯМХФЕМХХ ЧМШУ Х МЕЮЙРХБХПНБЮММШУ ХУ ТНПЛ, Б СБЕКХВЕМХХ ЯЙНПНЯРХ Х ХМРЕМЯХБМНЯРХ ЮЦПЕЦЮЖХХ, СЯХКЕМХХ ЮДЦЕ-ГХБМНЯРХ. рЮЙХЛ НАПЮГНЛ, ОНЙЮГЮМЮ ГМЮВХЛНЯРЭ ДЮММШУ ОН ЮЦПЕЦЮРН- Х ЮДЦЕГХН-ЛЕРПХХ РПНЛАНЖХРНБ ДКЪ ОНМХЛЮМХЪ ЛЕУЮМХГЛЮ ЮЙРХБЮЖХХ ОПНЖЕЯЯЮ РПНЛАН-НАПЮГНБЮМХЪ ОПХ ЯРЮПЕМХХ (йНПЙСЬЙН Х ДП., 1996). аНКЕЕ РНЦН, ОНБШЬЕМХЕ ЮЦПЕЦЮЖХНММНИ ЮЙРХБМНЯРХ РПНЛАНЖХРНБ ОПХ ЯРЮПЕМХХ ЯНВЕРЮЕРЯЪ ЯН ЯМХФЕМХ-ЕЛ ХУ ЯОНЯНАМНЯРХ Й ДЕОНМХПНБЮМХЧ АХНКНЦХВЕЯЙХ ЮЙРХБМШУ БЕЫЕЯРБ, МЮПСЬЕ-МХЕЛ ХУ ЩМЕПЦЕРХВЕЯЙНЦН НАЕЯОЕВЕМХЪ, Ю НДМХЛ ХГ МЮХАНКЕЕ ГМЮВХЛШУ ЛЕУЮМХГ-ЛНБ ДЕЯРЮАХКХГЮЖХХ ТСМЙЖХНМЮКЭМНЦН ЯНЯРНЪМХЪ РПНЛАНЖХРНБ ОПХ ЯРЮПЕМХХ ЯВХРЮЧР ЛЕЛАПЮММШИ ЛЕУЮМХГЛ (яЮПЙХЯНБ Х ДП., 1998). бЯЕ ЩРХ ХГЛЕМЕМХЪ КЕФЮР Б НЯМНБЕ ЮЙРХБЮЖХХ БМСРПХЯНЯСДХЯРНЦН РПНЛАННАПЮГНБЮМХЪ ОПХ ЯРЮПЕМХХ, ВРН ЯОНЯНАЯРБСЕР ПЮГБХРХЧ ЯЕПДЕВМН-ЯНЯСДХЯРНИ ОЮРНКНЦХХ С КХЖ ОНФХКНЦН БНГПЮЯРЮ (цЮБПХЬ Х ДП., 2001).оНБШЬЕМХЧ ЮДЦЕГХБМШУ Х ЮЦПЕЦЮЖХНММШУ ЯБНИЯРБ РПНЛАНЖХРНБ ЯОНЯНАЯРБСЧР РЕ ФЕ ОПННЙЯХДЮМРМШЕ СЯКНБХЪ Б ЙПНБХ КХЖ ЯРЮПЬХУ БНГПЮЯРМШУ ЦПСОО Х Б ЯЮЛХУ РПНЛАНЖХРЮУ. мЕ ОЕПЕВХЯКЪЪ ХГБЕЯРМШЕ ОН ЩРНИ ВЮЯРХ ПЮАНРШ, НРЛЕРХЛ КХЬЭ ЯКЕДСЧЫХИ ТЮЙР: ОН ЛЕПЕ ЯРЮПЕМХЪ БНГПЮЯРЮЕР ВСБЯРБХРЕКЭМНЯРЭ ДШУЮРЕКЭМНЦН ЙНЛОКЕЙЯЮ I РПНЛАНЖХРНБ ВЕКНБЕЙЮ Й ХМЦХАХРНПЮЛ (ПНРЕМНМС Х ДП.). дЮММШИ ЩТТЕЙР, ПЮЯЯЛЮРПХБЮЕЛШИ ЙЮЙ БНГЛНФМШИ АХНЛЮПЙ╦П ЯРЮПЕМХЪ Х ОНДРБЕПФДЕМХЕ ЛХРНУНМДПХЮКЭМНИ ЕЦН ЙНМЖЕОЖХХ (Pich et al., 1996), ЛНФЕР АШРЭ ЯКЕДЯРБХЕЛ РНЦН, ВРН Б ЯРЮПЕЧЫХУ ЙКЕРЙЮУ ТЕПЛЕМРШ ДШУЮРЕКЭМНИ ЖЕОХ ВЮЯРХВМН ХМЮЙРХБХПСЧРЯЪ (ЯЛ. О. 1.3.1). нВЕБХДМН, ЩРНР ╚ГЮДЕК╩ (╚ОНДЦНРНБКЕММНЯРЭ╩) ОНГБНКЪЕР СФЕ Я ЛЕМЭЬХЛХ СЯХКХЪЛХ ОПНДНКФЮРЭ ОПХМСДХРЕКЭМНЕ ХМЦХАХПНБЮМХЕ ДШУЮРЕКЭМНЦН ЙНЛОКЕЙЯЮ, ВЕЛ Б ЯКСВЮЕ Я ЛНКНДШЛХ ЙКЕРЙЮЛХ, ВРН, ОН-БХДХЛНЛС, Х БНЯОПХМХЛЮЕРЯЪ ЙЮЙ БНГПЮЯРЮМХЕ ВСБЯРБХРЕКЭМНЯРХ. н ОПХВЮЯРМНЯРХ ХГАШРНВМШУ НЙХЯКХРЕКЭМШУ ОПНЖЕЯЯНБ Й ЮЦПЕЦЮЖХХ РПНЛАНЖХРНБ Х РПНЛАННАПЮГНБЮМХЧ СЙЮГШБЮЧР ТЮЙРШ ОПНРХБНДЕИЯРБХЪ ХЛ ЯН ЯРНПНМШ ПЮГКХВМШУ ЮМРХНЙЯХДЮМРНБ, Б РНЛ ВХЯКЕ ЯХМРЕРХВЕЯЙХУ. оНЯКЕДМХЕ (ХНМНК, ЛЕЙЯХДНК Х ДП.) ЩТТЕЙРХБМН ХМЦХАХПСЧР ЮЦПЕЦЮЖХЧ РПНЛАНЖХРНБ ЙЮЙ in vitro, РЮЙ Х ОПХ ББЕДЕМХХ Б НПЦЮМХГЛ (ОНЙЮГЮМН МЮ ОПХЛЕПЕ ЙПШЯ), Б ПЕГСКЭРЮРЕ РПНЛАНЦЕММШИ ОНРЕМЖХЮК ЙПНБХ СЛЕМЭЬЮЕРЯЪ (яОЮЯНБ Х ДП., 1999).
1.5.2. оПХ ЯРЮПЕМХХ НПЦЮМХГЛЮ ХГЛЕМЕМХЪ ОПНХЯУНДЪР Х Б ДПСЦХУ ГБЕМЭЪУ ЯХЯРЕЛШ РПЮМЯОНПРЮ н2. оПЕФДЕ БЯЕЦН, НМХ ГЮРПЮЦХБЮЧР ЯЮЛС ЯНЯСДХЯРСЧ ЯХЯРЕЛС МЮ БЯЕУ Е╦ СПНБМЪУ. йПНБЕМНЯМШЕ ЯНЯСДШ ЯРЮМНБЪРЯЪ ЛЕМЕЕ ЮДЮОРХБМШЛХ, Р.Е. ОНЯРЕОЕММН РЕПЪЧР СОПСЦНЯРЭ, ЩКЮЯРХВМНЯРЭ Х ЯОНЯНАМНЯРЭ ХГЛЕМЪРЭ ЯБНИ ОПНЯБЕР. нАЫЕЕ ПСЯКН ЙЮОХККЪПНБ Х ЮПРЕПХНК ЯСФЮЕРЯЪ, Ю ЙПНБЪМНЕ ДЮБКЕМХЕ Я БНГПЮЯРНЛ СБЕКХВХБЮЕРЯЪ (мЮЦНПМШИ Х ДП., 1963; йНПЙСЬЙН, йНБЮКЕМЙН, 1988; Marin, Rodriguez-Martinez, 1999). оНБШЬЕМХЕ ФЕЯРЙНЯРХ ЯНЯСДХЯРНИ ЯРЕМЙХ УЮПЮЙРЕПХГСЕРЯЪ СБЕКХВЕМХЕЛ НРМНЬЕМХЪ ЙНККЮЦЕМ/ЩКЮЯРХМ Х ДХТТСГМШЛ НРКНФЕМХЕЛ ЙЮКЭЖХЪ Х КХОХДНБ (Robert, 1999). сЛЕЭЬЕМХЕ РЕЙСВЕЯРХ ОКЮГЛЮРХВЕЯЙНИ ЛЕЛАПЮМШ ЙКЕРНЙ ЩМДНРЕКХЪ ЮНПРШ ЯРЮПЕЧЫХУ ЙПШЯ, ЯБЪГШБЮЕЛНЕ Я ХГЛЕМЕМХЕЛ ЯНДЕПФЮМХЪ УНКЕЯРЕПХМЮ Х ОЕПЕЙХЯЕИ КХОХДНБ, БШГШБЮЕР ДХЯТСМЙЖХЧ СЙЮГЮММШУ ЙКЕРНЙ (Hashimoto et al., 1999). аНКЕЕ ЯРЮПШЕ ЦКЮДЙНЛШЬЕВМШЕ ЙКЕ-РЙХ ЯНЯСДНБ СЯЙНПЕММН ОЕПЕУНДЪР НР ЯНЙПЮРХРЕКЭМНЦН Й ЯХМРЕРХВЕЯЙНЛС ТЕМН-РХОС (Lundberg, Crow, 1999).
оПХ ЯРЮПЕМХХ ВЕКНБЕЙЮ СЛЕМЭЬЮЕРЯЪ ЙНКХВЕЯРБН ТСМЙЖХНМХПСЧЫХУ ЙЮОХККЪПНБ МЮ ЕДХМХЖС ОКНЫЮДХ РЙЮМХ, ГЮЛЕДКЪЕРЯЪ ЙПНБНРНЙ МЮ БЯЕУ СВЮЯРЙЮУ ЛХЙПНЖХПЙСКЪРНПМНЦН ПСЯКЮ. б НПЦЮМЕККЮУ Х ЛЮРПХЙЯЕ ЩМДНРЕКХНЖХРНБ ЙЮОХККЪПНБ ЙНФХ БШЪБКЪЧРЯЪ ДЕЯРПСЙРХБМШЕ Х ДХЯРПНТХВЕЯЙХЕ ХГЛЕМЕМХЪ (яЮПЙХЯНБ Х ДП., 1998). оНЯКЕДМХЕ ДНКФМШ, ЕЯРЕЯРБЕММН, ЯЙЮГШБЮРЭЯЪ МЮ ЩТТЕЙРХБМНЯРХ ТСМЙЖХНМХПНБЮМХЪ ЙПНБЕМНЯМНИ ЯХЯРЕЛШ БННАЫЕ, Х МЮ РПЮМЯОНПРЕ н2 Б ВЮЯРМНЯРХ. б ЖЕКНЛ ФЕ, НЯМНБМНИ ОПХВХМНИ ДХЯТСМЙЖХХ ЩМДНРЕКХЪ ЯНЯСДНБ ЪБКЪЕРЯЪ, НВЕБХДМН, ХГАШРНВМЮЪ ЦЕМЕПЮЖХЪ Б МХУ ютй. яННАЫЮЕРЯЪ, МЮОПХЛЕП, ВРН С 12-ЛЕЯЪВМШУ ЙПШЯ Sprague-Dawley НАПЮГНБЮМХЕ н АШКН АНКЕЕ ЩМЕПЦХВМШЛ, ВЕЛ С 3-4-ЛЕЯЪВМШУ ФХБНРМШУ (Hamilton et al., 2001).
мЕЙНРНПШЕ ДПСЦХЕ ХГЛЕМЕМХЪ ЯРЕМНЙ ЯНЯСДНБ, ЯНЯРЮБКЪЧЫХЕ ЯСРЭ ХУ БНГПЮЯРМНИ ОЮРНКНЦХХ, ЯУНДМШ Я ЦХЯРНКНЦХВЕЯЙХЛХ УЮПЮЙРЕПХЯРХЙЮЛХ ЮРЕПНЯЙКЕПНРХВЕЯЙХУ ЯНЯСДНБ (ЯЛ. О. 1.7.1.1), МН БЯ╦ ФЕ ОПЕДЯРЮБКЪЧР ДБЮ ПЮГМШУ ЪБКЕМХЪ. рЕЛ МЕ ЛЕМЕЕ, ЛЕФДС МХЛХ ХЛЕЕРЯЪ ЯБЪГЭ: ЯРЕМЙХ ЯРЮПШУ ЯНЯСДНБ НЯНАН ОПЕДПЮЯОНКНФЕМШ Й ЮРЕПНЯЙКЕПНРХВЕЯЙХЛ ОНБПЕФДЕМХЪЛ, ЪБКЪЧРЯЪ БЮФМШЛХ ОПЕДЬЕЯРБЕММХЙЮЛХ Х СЯЙНПХРЕКЪЛХ ПЮГБХРХЪ ЮРЕПНЯЙКЕПНГЮ (Lundberg, Crow, 1999; Robert, 1999). щРН ЛМЕМХЕ ОНДРБЕПФДЕМН КХРЕПЮРСПМШЛХ Х ЯНАЯРБЕММШЛХ ДЮМ-МШЛХ Х ЮБРНПНБ ПЮАНРШ (рЕПЕЬiМЮ Х ДП., 2000), ХГСВЮБЬХЛХ ПНКЭ БНГПЮЯРЮ Б ПЕЮЙЖХХ ЯНЯСДНБ МЮ ПЮГКХВМШЕ ОНБПЕФДЮЧЫХЕ ТЮЙРНПШ. йЮЙ ОНЙЮГЮМН, ХГЛЕМЕМХЪ, МЮЯРСОЮЧЫХЕ Б ЯРЮПНЯРХ Б ЙКЕРНВМШУ ЩКЕЛЕМРЮУ ЯРЕМЙХ ЯНЯСДНБ МЮ ТСМЙ-ЖХНМЮКЭМНЛ Х ЯРПСЙРСПМНЛ СПНБМЪУ, СБЕКХВХБЮЧР БНЯОПХХЛВХБНЯРЭ ЯНЯСДНБ Й ОНБПЕФДЮЧЫХЛ БКХЪМХЪЛ Х СЯЙНПЪЧР ПЮГБХРХЕ ЮРЕПНЯЙКЕПНГЮ. дКЪ ЯРЮПШУ ЙКЕ-РНЙ ЯНЯСДХЯРНИ ЯРЕМЙХ Б СЯКНБХЪУ БНГМХЙЬЕЦН Б МХУ НРМНЯХРЕКЭМН ОНБШЬЕМ-МНЦН ЙХЯКНПНДМН-ОЕПЕЙХЯМНЦН ЯНЯРНЪМХЪ РЮЙЮЪ ПЕЮЙЖХЪ, Б РНЛ ВХЯКЕ ОПЕДПЮЯОНКНФЕММНЯРЭ Й ПЮГБХРХЧ ЮРЕПНЯЙКЕПНГЮ, ОПЕДЯРЮБКЪЕРЯЪ ЕЯРЕЯРБЕММНИ. йЯРЮРХ, ЯКЕДЯРБХЕЛ РНЦН ФЕ ЯНЯРНЪМХЪ ЪБКЪЕРЯЪ, НВЕБХДМН, Х РНР ТЮЙР, ВРН С ЯРЮПШУ (24-26 ЛЕЯ.) ЙПШЯ ЯНЯСДШ ВСБЯРБХРЕКЭМШ Й ДЕИЯРБХЧ ХНМХГХПСЧЫЕЦН ХГКСВЕМХЪ, Ю С БГПНЯКШУ (8-10 ЛЕЯ.) НМХ НРМНЯХРЕКЭМН ПЕГХЯРЕМРМШ (тПНКЭЙХЯ, яШЙЮКН, 2000).
сВХРШБЮЪ ПЮГКХВХЕ Б ГМЮВЕМХЪУ Пн2 Б ЮПРЕПХЮКЭМНИ Х БЕМНГМНИ ЙПНБХ, КНЦХВМН ОНКЮЦЮРЭ, ВРН ЯРЕМЙХ ЮПРЕПХИ ЯРЮПЕЧР Я АНКЕЕ БШЯНЙНИ ЯЙНПНЯРЭЧ, ВЕЛ ЯРЕМЙХ БЕМ. рЮЙНЕ ЛМЕМХЕ ОНДДЕПФХБЮЕРЯЪ Х ДЮММШЛХ Н РНЛ, ВРН СЙНПНВЕМХЕ ДКХМШ РЕКНЛЕПМНИ дмй, ЯВХРЮЧЫЕЕЯЪ ОН РЕНПХХ РЕКНЛЕПМНЦН ЯРЮПЕМХЪ ЯНЛЮРХВЕЯЙХУ ЙКЕРНЙ ОНЙЮГЮРЕКЕЛ ПЕОКХЙЮРХБМНЦН ЯРЮПЕМХЪ (ЯЛ. О. 1.4.3), ОПНХЯУНДХР Б ЯРЕМЙЮУ ЮПРЕПХИ СЯЙНПЕММН Б ЯПЮБМЕМХХ Я РЮЙНБШЛ Б БЕМЮУ, ОПХВ╦Л ЩРНР ОПНЖЕЯЯ НАЗЪЯМЪЕРЯЪ ДЕИЯРБХЕЛ ЯБНАНДМШУ ПЮДХЙЮКНБ (Chang, Harley, 1995). мЕСДХБХРЕКЭМН РЮЙФЕ, ВРН Б ЯБЪГХ Я ОНБШЬЕММНИ БЕПНЪРМНЯРЭЧ ПЮГБХРХЪ Б ЯНЯСДЮУ НЙХЯКХРЕКЭМНЦН ЯРПЕЯЯЮ ОПХПНДНИ ОПЕДСЯЛНРПЕМШ НОПЕДЕК╦ММШЕ ЛЕПШ ОН ГЮЫХРЕ ХУ НР БНГЛНФМНЦН ОНБПЕФДЕМХЪ. нДМНИ ХГ РЮЙХУ ЛЕП ЛНФМН ЯВХРЮРЭ ХМДСЙЖХЧ ЦЕМЮ ЦЕЛНЙЯХЦЕМЮГШ √ ЯРПЕЯЯНПМНЦН ТЕПЛЕМРЮ, ЙНРНПШИ БШОНКМЪЕР НЙЯХОПНРЕЙРНПМСЧ ТСМЙЖХЧ ОСР╦Л ЦЕМЕПЮЖХХ ХГ ЦЕЛЮ БЮГНДХКЪРЮРНПЮ ян Х АХКХПСАХМЮ, НАКЮДЮЧЫЕЦН ЮМРХНЙЯХДЮМРМШЛХ ЯБНИЯРБЮЛХ (Deramaudt et al., 1998). бННАЫЕ, ЦЕЛНЙЯХЦЕМЮГЮ ОПХГМЮ╦РЯЪ ТЕПЛЕМРНЛ, ХМДСЖХПСЕЛНЛ ОПХ НЙХЯКХРЕКЭМНЛ ЯРПЕЯЯЕ (Dubrovskaya, Wetterhahn, 1998). оНЯКЕДМХИ, МЮОПХЛЕП, БШ-ГШБЮЕР МЮЙНОКЕМХЕ Лпмй ЦЕЛНЙЯХЦЕМЮГШ Б ТХАПНАКЮЯРЮУ ВЕКНБЕЙЮ, РСЬХРЕКЭ ФЕ ЯБНАНДМШУ ПЮДХЙЮКНБ н2 ОПЕДНРБПЮЫЮЕР ЩРНР ЩТТЕЙР (Numasawa et al., 1997).
мЕ ЛЕМЕЕ ОПХБКЕЙЮРЕКЕМ ТЕМНЛЕМ СЯХКЕМХЪ ОПНКХТЕПЮЖХХ ЩМДНРЕКХЮКЭМШУ ЙКЕРНЙ Х ОПНЖЕЯЯЮ ЮМЦХНЦЕМЕГЮ ЛХЙПНЯНЯСДНБ ОПХ ХГАШРНВМНИ ЩЙЯОПЕЯЯХХ ЦЕЛНЙЯХЦЕМЮГШ √ ТЮЙР, ПЮЯЯЛЮРПХБЮЕЛШИ ЙЮЙ СВЮЯРХЕ ЩРНЦН ТЕПЛЕМРЮ Б ОПНКХТЕПЮРХБМНЛ НРБЕРЕ ЩМДНРЕКХЪ МЮ НЙХЯКХРЕКЭМШИ ЯРПЕЯЯ (Deramaudt et al., 1998). бЯЕ ЩРХ ДЮММШЕ ЯБХДЕРЕКЭЯРБСЧР, МЮ МЮЬ БГЦКЪД, Н РНЛ, ВРН Б СЯКНБХЪУ НЙХЯ-КХРЕКЭМНЦН ЯРПЕЯЯЮ ЦЕЛНЙЯХЦЕМЮГЮ БЛЕЯРЕ Я ДПСЦХЛХ ЯНЯРЮБКЪЧЫХЛХ ЮМРХНЙ-ЯХДЮМРМНИ ЯХЯРЕЛШ ОПЕОЪРЯРБСЕР СЯРЮМНБКЕМХЧ Б ЩМДНРЕКХЮКЭМШУ ЙКЕРЙЮУ ХГАШРНВМШУ ГМЮВЕМХИ ДХЯАЮКЮМЯЮ ∆ (он √ юн), ОПХБНДЪЫХУ Й ПЮГБХРХЧ БНГПЮЯРМШУ ОЮРНКНЦХВЕЯЙХУ ХГЛЕМЕМХИ, ЮОНОРНГС ХКХ НЙХЯКХРЕКЭМНЛС ХУ ЖХРН-КХГС. яЙНПЕЕ БЯЕЦН, ЦЕЛНЙЯХЦЕМЮГЮ СВЮЯРБСЕР Б НЦПЮМХВЕМХХ СЙЮГЮММНЦН ДХЯ-АЮКЮМЯЮ ДН СПНБМЕИ, ДНЯРЮРНВМШУ ДКЪ ПЕЮКХГЮЖХХ Б МНПЛЮКЭМШУ ЙКЕРЙЮУ ЩМДНРЕКХЪ НЙХЯКХРЕКЭМНЦН ЛХРНЦЕМЕГЮ. я ДПСЦНИ ЯРНПНМШ, ОНМЪРМН, ОНВЕЛС ЩЙЯОПЕЯЯХЪ ЦЕЛНЙЯХЦЕМЮГШ Б ЙКЕРЙЮУ ЩМДНРЕКХЪ ЯНЯСДНБ ПЕОПЕЯЯХПСЕРЯЪ ОПХ ЦХОНЙЯХХ (Nakayama et al., 2000) √ Б ЩРХУ СЯКНБХЪУ МЕНАУНДХЛНЯРЭ Б ДЮММНЛ ТЕПЛЕМРЕ ОПНЯРН НРОЮДЮЕР.
мЕЯНЛМЕММШИ ХМРЕПЕЯ ОПЕДЯРЮБКЪЧР РЮЙФЕ ДЮММШЕ Н БКХЪМХХ МЕЙНРНПШУ ЙНЛОНМЕМРНБ ОКЮГЛШ ЙПНБХ МЮ ЯНЯРНЪМХЕ Х ТСМЙЖХЧ ЛХРНУНМДПХИ, ОПЕФДЕ БЯЕЦН Б ЙКЕРЙЮУ ЯРЕМЙХ ЯНЯСДНБ. нДМХЛ ХГ РЮЙХУ ЙНЛОНМЕМРНБ ЯВХРЮЧР ЮКЭАС-ЛХМ, БГЮХЛННРМНЬЕМХЪ ЙНРНПНЦН Я ЛХРНУНМДПХЪЛХ НРПЮФЮЧР ХГЛЕМЕМХЪ, ОПН-ХЯУНДЪЫХЕ Б ОПНЖЕЯЯЕ ЯРЮПЕМХЪ Х ЯРПЕЯЯЮ. оНД БКХЪМХЕЛ ОНЯКЕДМХУ СЛЕМЭ-ЬЮЕРЯЪ ЙНКХВЕЯРБН ЛХРНУНМДПХИ, ЮКЭАСЛХМ ФЕ ЙНЯБЕММН ОПНРХБНДЕИЯРБСЕР ЩРНЛС, ХЯОНКЭГСЪ ЯБНЧ РПЮМЯОНПРМСЧ ТСМЙЖХЧ ДКЪ НАЕЯОЕВЕМХЪ МНПЛЮКЭМНИ ФХГМЕДЕЪРЕКЭМНЯРХ ЛХРНУНМДПХИ ОНЯРЮБЙНИ ЙЮКЭЖХЪ, ЛЮЦМХЪ, ФХПМШУ ЙХЯКНР Х МЮАНПЮ МЕНАУНДХЛШУ ЮЛХМНЙХЯКНР. мЮ АНКЭЬНЛ ВХЯКЕ ХЯЯКЕДНБЮМХИ ОНЙЮГЮМН, ВРН НОРХЛЮКЭМНЕ ЯНДЕПФЮМХЕ ЮКЭАСЛХМЮ Б ЯШБНПНРЙЕ ЙПНБХ ДНКФМН АШРЭ > 47 Ц/К ОПХ НРМНЬЕМХХ ЮКЭАСЛХМ/ЦКНАСКХМ, ПЮБМНЛ 2. оПХ ЩРХУ СЯКНБХЪУ ╚МЕ МЮАКЧДЮЕРЯЪ НАЬХПМНИ ЦХАЕКХ ЙКЕРНЙ, ЯСЫЕЯРБЕММШУ ЩМЕПЦЕРХВЕЯЙХУ ОНРЕПЭ Х ХЯРНЫЕМХЪ, ПЮГБХРХЪ НМЙНКНЦХВЕЯЙХУ ГЮАНКЕБЮМХИ ХКХ яохдЮ╩ (Seaton, 1995). нДМЮЙН НРЛЕВЕММЮЪ БШЬЕ ОПННЙЯХДЮМРМЮЪ ЯХРСЮЖХЪ Б ЙПНБХ ЯРЮПЕЧЫЕЦН Х, РЕЛ АНКЕЕ, ОНПЮФ╦ММНЦН БНГПЮЯРМШЛХ АНКЕГМЪЛХ НПЦЮМХГЛЮ (ЯЛ. О. 1.7) НЦПЮМХВХ-БЮЕР, ОН-БХДХЛНЛС, ОНГХРХБМНЕ ДЕИЯРБХЕ ЮКЭАСЛХМЮ БЯКЕДЯРБХЕ ЕЦН НЙХЯКХ-РЕКЭМНЦН ОНБПЕФДЕМХЪ. йЯРЮРХ, ЩРН ОНБПЕФДЕМХЕ ОНДЮБКЪЕРЯЪ ТКЮБНМНХДНЛ ЯХКХАХМХМНЛ, ЙНРНПШИ ХМЦХАХПСЕР РЮЙФЕ БШГШБЮЕЛНЕ ХНМЮЛХ ЛЕДХ онк Х ЦЕЛНКХГ ЩПХРПНЖХРНБ (Filipe et al., 1997).
я БНГПЮЯРНЛ МЕЦЮРХБМШЕ ХГЛЕМЕМХЪ ОПНХЯУНДЪР Х Б ЯЮЛНЛ ЯЕПДЖЕ √ МЮЯНЯЕ ЙПНБННАПЮЫЕМХЪ. нА ЩРНЛ ЯБХДЕРЕКЭЯРБЕР, МЮОПХЛЕП, ОНБШЬЕММЮЪ ВСБ-ЯРБХРЕКЭМНЯРЭ ЯЕПДЖЮ ЯРЮПШУ ЙПШЯ Й ЙЮПДХНРНЙЯХВЕЯЙХЛ ДНГЮЛ ЮДПЕМЮКХМЮ. оНБПЕФДЮЧЫЕЕ ДЕИЯРБХЕ ЕЦН Б ГМЮВХРЕКЭМНИ ЯРЕОЕМХ НОПЕДЕКЪЕРЯЪ МХГЙНИ ЮЙРХБМНЯРЭЧ ЮМРХНЙЯХДЮМРМНИ ЯХЯРЕЛШ Х БШЯНЙНИ √ ОЕПЕЙХЯМНЦН НЙХЯКЕМХЪ Б ЯЕПДЕВМНИ ЛШЬЖЕ ЯРЮПШУ ФХБНРМШУ (лЮПЙНБЮ, лХЯСКЮ, 1992).
нАЕЯОЕВЕМХЕ ЙХЯКНПНДМНЦН ПЕФХЛЮ НПЦЮМХГЛЮ ГЮБХЯХР РЮЙФЕ НР ЯНЯРНЪМХЪ ЯХЯРЕЛШ БМЕЬМЕЦН ДШУЮМХЪ. оПХ ЯРЮПЕМХХ ЯЮЛХ К╦ЦЙХЕ ЯРЮМНБЪРЯЪ ЛЕМЕЕ ОНД-БХФМШЛХ, ЯМХФЮЕРЯЪ ХУ ФХГМЕММЮЪ ╦ЛЙНЯРЭ, ХГЛЕМЪЧРЯЪ Х ДПСЦХЕ ХУ ОЮПЮ-ЛЕРПШ. уНРЪ ДШУЮРЕКЭМШИ НАЗ╦Л Б ЯРЮПНЯРХ СЛЕМЭЬЮЕРЯЪ, МН НДМНБПЕЛЕММН ПЮГБХБЮЧЫЕЕЯЪ СВЮЫЕМХЕ ДШУЮМХЪ ОПХБНДХР Й СБЕКХВЕМХЧ БЕМРХКЪЖХХ К╦ЦЙХУ (ЯЛ. кЮСЩП, яЕПЕДЕМЙН, 1982). щРХ ФЕ ЮБРНПШ НАПЮЫЮЧР БМХЛЮМХЕ МЮ ЯКЕДСЧ-ЫХИ ТЮЙР: ЯМХФЕМХЕ Пн2 Б ЮПРЕПХЮКЭМНИ ЙПНБХ Б ЯРЮПНЯРХ МЕЮДЕЙБЮРМН ЯПЮБ-МХРЕКЭМН БШЯНЙНЛС Пн2 Б ЮКЭБЕНКЪПМНЛ БНГДСУЕ, ВРН ЪБКЪЕРЯЪ ОПХВХМНИ СБЕКХВЕМХЪ ЮКЭБЕНКЪПМН-ЮПРЕПХЮКЭМНЦН ЦПЮДХЕМРЮ ЙХЯКНПНДЮ. б ЖЕКНЛ БНГПЮЯР-МШЕ ХГЛЕМЕМХЪ Б ЙПНБЕМНЯМНИ Х ДШУЮРЕКЭМНИ ЯХЯРЕЛЮУ ЯНГДЮЧР МЕАКЮЦНОПХЪР-МШЕ СЯКНБХЪ ДКЪ НАЕЯОЕВЕМХЪ НПЦЮМХГЛЮ ЙХЯКНПНДНЛ. бНОПНЯ Н РНЛ, ЙЮЙ Б ОНЯКЕДСЧЫЕЛ ЩРН ЛНФЕР НРПЮГХРЭЯЪ МЮ ЯНЯРНЪМХХ НПЦЮМХГЛЮ Х ОПНДНКФЕМХХ ЯРЮПЕМХЪ, НРВЮЯРХ АШК СФЕ ГЮРПНМСР Б ОПЕДШДСЫХУ ОЮПЮЦПЮТЮУ Х АСДЕР ЕЬ╦ НАЯСФДЕМ Б О. 1.6.
пЮЯЯЛНРПЕМХЕ СЯКНБХИ, ОПХБНДЪЫХУ Й НЙХЯКХРЕКЭМШЛ ОНБПЕФДЕМХЪЛ Б ЯХЯРЕЛЮУ ЯМЮАФЕМХЪ НПЦЮМХГЛЮ ЙХЯКНПНДНЛ, ГЮБЕПЬХЛ ЯЯШКЙНИ МЮ ЯКЕДСЧЫХЕ КЧАНОБРМШЕ ЛЮРЕПХЮКШ. нЙЮГШБЮЕРЯЪ, МЮПЪДС Я СОНЛЪМСРШЛХ БШЬЕ КНЙЮКЭМШ-ЛХ ЛЕПЮЛХ ОПНРЕЙЖХХ НР ОЕПНЙЯХЦЕМЮГМНЦН ЯРПЕЯЯЮ С ЛКЕЙНОХРЮЧЫХУ ЯСЫЕЯ-РБСЧР Х НРМНЯХРЕКЭМН ГЮЖЕМРПЮКХГНБЮММШЕ ЛЕУЮМХГЛШ ГЮЫХРШ. нА ЩРНЛ МЮЛ ЯРЮКН ХГБЕЯРМН ХГ ЯРЮРЭХ яЙСКЮВЕБЮ (2001), ЙНРНПШИ ОПХБ╦К ДЮММШЕ КХРЕПЮРСПШ Н м2н2-ЯЕМЯНПЮУ Б МЕИПНЩОХРЕКХЮКЭМШУ РЕКЭЖЮУ К╦ЦЙНЦН (Fu et al., 2000) Х ЙКЕРЙЮУ ЙЮПНРХДМНЦН ЯХМСЯЮ (Wang D. et al., 1996). оЕПБШИ ХГ МХУ НРБЕВЮЕР ГЮ ЯСФЕМХЕ ДШУЮРЕКЭМШУ ОСРЕИ, Ю БРНПНИ √ ЙПНБЕМНЯМШУ ЯНЯСДНБ ОПХ ОНБШЬЕМХХ СПНБМЕИ н2 Х (ХКХ) м2н2 МЕГЮБХЯХЛН НР ОПХВХМШ, БШГБЮБЬЕИ ЩРН ОНБШЬЕМХЕ. мЮГБЮММШЕ ЯХЯРЕЛШ ПЕЦСКЪЖХХ Я НРПХЖЮРЕКЭМНИ НАПЮРМНИ ЯБЪГЭЧ ╚ПЮЯЯВХРЮМШ╩, ОН-БХДХЛНЛС, КХЬЭ МЮ НОПЕДЕК╦ММШИ ДХЮОЮГНМ НРЙКНМЕМХИ НР МНПЛШ Пн2 Х ∆ он √юн) Б ДШУЮРЕКЭМШУ Х ЙПНБЕМНЯМШУ ОСРЪУ. оПХ АНКЕЕ ФЕ ЯХКЭМШУ БЕКХВХМЮУ ЩРХУ НРЙКНМЕМХИ, БНГМХЙЮЧЫХУ ОПХ ЯРЮПЕМХХ Х МЕЙНРНПШУ ОЮРНКНЦХЪУ, НМХ ДЮЧР ЯАНХ, ВРН ДНКФМН ЯОНЯНАЯРБНБЮРЭ МЕЦЮРХБМШЛ ХГЛЕМЕМХЪЛ Б ДШУЮРЕКЭМНИ Х ЙПНБЕМНЯМШУ ЯХЯРЕЛЮУ (ЯЛ. БШЬЕ).
еЫЕ ОН РЕЛЕ 1.5.1. жХРНРНЙЯХВЕЯЙХЕ ОПНДСЙРШ ЛЕРЮАНКХГЛЮ н2:
- нРПЮБКЪЧЫХЕ Х БШЯНЙНРНЙЯХВМШЕ БЕЫЕЯРБЮ ЖХРНРНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- йКЧВЕБШЕ ТСМЙЖХХ НПЦЮМХГЛЮ ВЕКНБЕЙЮ, ОНГХРХБМНЕ БНГДЕИЯРБХЕ МЮ ЙНРНПШЕ ОНГБНКЪЕР НРМНЯХРЭ ОПНДСЙРШ Й ЙЮРЕЦНПХХ ОПНДСЙРНБ ТСМЙЖХНМЮКЭМНЦН ОХРЮМХЪ:
- 1.6.рНЙЯХВМШЕ УХЛХВЕЯЙХЕ БЕЫЕЯРБЮ ЖХРНРНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ
- 5.3 нОПЕДЕКЕМХЕ ЖХРНРНЙЯХВЕЯЙНИ ЮЙРХБМНЯРХ ЛЮЙПНТЮЦНБ ОН НРМНЬЕМХЧ Й ЬРЮЛЛЮЛ C. neoformans ПЮГМНИ БХПСКЕМРМНЯРХ
- 3.2.9 нОПЕДЕКЕМХЕ ЖХРНРНЙЯХВЕЯЙНИ ЮЙРХБМНЯРХ ОЕПХРНМЕЮКЭМШУ ЛЮЙПНТЮЦНБ
- 5.2.5. оПЕОЮПЮРШ, НЙЮГШБЮЧЫХЕ БНГДЕИЯРБХЕ МЮ ЖХРНРНЙЯХВЕЯЙСЧ ЮЙРХБМНЯРЭ Х ХМРЕПТЕПНМНОПНДСЙЖХЧ
- рЕЛЮ ╧ 5 рНЙЯХВМШЕ УХЛХВЕЯЙХЕ БЕЫЕЯРБЮ ЖХРНРНЙЯХВЕЯЙНЦН ДЕИЯРБХЪ.
- бРНПХВМШИ ЛЕРЮАНКХГЛ ЦПХАНБ
- лЕРЮАНКХГЛ ЯТХМЦНКХОХДНБ
- лЕРЮАНКХГЛ ЯСАЯРПЮРНБ ОПХ ЬНЙЕ.
- 5. юЛХМНЙХЯКНРШ: ЛЕРЮАНКХГЛ Х ТСМЙЖХХ