СТАДНЫЕ И АТРЕГАПИОПНЫЕ РЕАКЦИИ
Подражательное поведение и возникающие на его основе явления, однако, не исчерпывают всех реакций животных на себе подобных и являются скорее лишь частной и весьма специализированной формой врожденной физиологической реакции.
И тем не менее есть все основания думать, что именно на основе этого подражательного поведения, фиксирующего расстояние между особями, и создается объединение животных в группы — агрегации (Слоним, 1965). Само же существование животных в группе приводит к ряду физиологических изменений в организме каждого члена группы. Лучше других при этом выявляются реакции общего обмена веществ и эндокринные, главным образом со стороны коры надпочечников.Изменения газообмена в группе лучше всего выявляются в состоянии ПОЛНОГО 'Мышечного покоя. Если взять группу лабораторных белых крыс и поместить их в условия низкой температуры, то они образуют скопление: садятся друг на друга, постоянно меняют положение (крайние особи протискиваются внутрь группы) (Hill, 1913). Отмечено, что при групповом скоплении газообмен у них резко снижен. Это явление тогда же было истолковано Хиллом как проявление терморегуляции, как следствие ограничения теплоотдачи в связи с тем, что суммарная поверхность тела животных, собранных вместе, значительно меньше отдающей тепло поверхности животных, посаженных врозь. Это, если можно так выразиться,— коллективная терморегуляция. Однако уже расчеты Хилла обнаружили недостаточность такого объяснения, так как уменьшение поверхности тела было меньше, нежели соответственное падение теплообразования. Хилл считал, что здесь имеет место снижение двигательной активности животных.
Хорошо выраженный двигательный аффект в группе был недавно обнаружен Ю. Кескпайком и А. Ф. Давыдовым (1966). Оказалось, что с развитием терморегуляции у птенцов появляется хорошо выраженное «скучивание» при умеренном охлаждении, когда родители покидают гнездо.
На рисунке 83 показаны позы птенцов и их реакция через 20 мин на изменение температуры камеры. Птенцы в возрасте четырех суток «скучиваются» при температуре 20° С и отодвигаются друг от друга при температуре 33° С. В первый день после вылупления скучивание наблюдается даже при температурах 34 и 35° С. Двигательная реакция на охлаждение появляется ранее, чем формируется терморегуляция. Последняя как бы за-меняет при определенных температурах реакцию окучивания. На рисунке 84 показаны те же отношения у птенцов разного возраста по наблюдениям, произведенным через 10 мин после вылета самки из гнезда.
Явление скучивания (агрегации) широко распространено и среди пойкилотермных организмов: массовые скопления лягушек и жаб, насекомых во время зимовки, массовые скопления моллюсков (АПее, 1931; Понугаева,
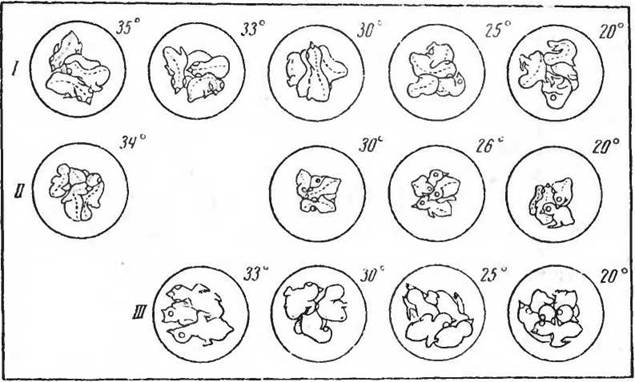
Рис. 83. Положение птенцов дрозда в гнезде при различной температуре среды (°С) через 15—20 мин после экспозиции в камере. I, II — 1-е сутки; /// — 4-е сутки
1953). Для таких скоплений характерно резкое суммарное понижение обмена веществ в покое, достигающее у представителей отдельных видов 40% и более (Понугаева, 1949, 1952, 1953, I960). Факт этот был обнаружен у моллюсков, рыб и лягушек, т. е. у животных, не обладающих терморегуляцией. Интересно, что у моллюсков понижение обмена веществ можно обнаружить не только при помещении их в группу живых моллюсков, но и среди вываренных раковин. Очевидно, это связано не только с химическими раздражителями, особенно интенсивными в водной среде.
М. Штефан (1957), а затем К. А. Алексеева (1959) на рыбах'подтвердили данные Хилла, Элли и А. Г. Пону- 391
гаевой. Было установлено, что удаление переднего мозга рыб влечет за собой выпадение стадного влияния на газообмен. Последний оказался неодинаково выраженным у разных видов рыб. У водных организмов исследователи неоднократно обнаруживали химические вещества, выделяющиеся в окружающую среду и могущие действовать на обмен веществ (телергоны) (Киршен- блат, 1968).
(Имеются многочисленные указания на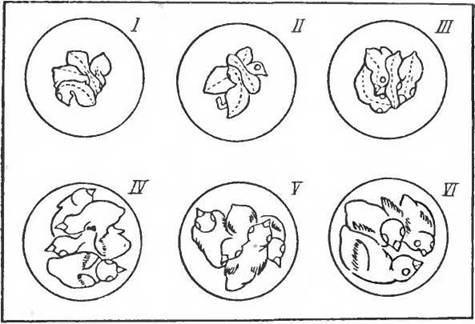
Рис. 84. Положение птенцов различного возраста в гнезде через 10 мин после вылета самки: / — односуточные; II— двухсуточные; III — трехсуточные; IV — четырехсуточные; V — пятнеуточные; VI — шестисуточные
вещества, стимулирующие и задерживающие рост растений, действующие через корневую систему.)
Особенно ярко изменение обмена веществ в стаде или группе проявляется у млекопитающих со стадным или колониальным образом жизни. Для получения «стадного» эффекта непосредственный термический контакт животных друг с другом не обязателен (Понугаева, 1960). Например, обезьяны собираются на ночь (особенно при низких температурах среды) в большие группы (Слоним и Щербакова, 1949). Снижение газообмена в группе обезьян можно наблюдать, если посадить животных в различные камеры. Контакт животных обеспечивается с по- 392
мощью стадных звуков, и обмен веществ в такой разделенной группе снижается.
У сусликов, живущих колониями, можно наблюдать яркий «стадный» эффект при отсаживании их в общую газообменную камеру. Физиологический механизм рефлекторного влияния на обмен веществ в группе подобных организмов подвергается тщательному изучению с целью определения роли различных анализаторов и сроков возникновения стадных рефлексов. В онтогенезе стадные рефлексы (изменения газообмена) возникают у грызунов сравнительно поздно (на 30-й день). Можно считать установленным, что этих рефлексов нет ни в момент рождения животного, ни в последующий период гнездового развития (Понугаева, I960). Понижения газообмена при групповом скоплении не наблюдается у представителей ряда млекопитающих, ведущих во взрослом состоянии одиночный образ жизни. Помещение таких животных, как серый хомячок (Mesocricetus griseus), незокия (Nesokia indica), ушастый с?к (Hemiechinus auritus), в среду себе подобных вызывает у них повышение газообмена (Понугаева, 1960).
В таблице 41 приведены основные величины изменений газообмена.
Изменения обмена веществ в группе и во время спячки не исчезают. Исследования, проведенные в пещере «Адзаба» возле Сухуми, в Саблинской пещере под Ленинградом на различных видах летучих мышей, обнаружили снижение обмена веществ в группе и в состоянии дневного сна у летучих мышей летом и во время зимней спячки (Понугаева, 1949; Понугаева и Слоним, 1953). Последний факт представляет большой интерес, так как поясняет значение скоплений животных в их зимних убежищах во время спячки. В этих условиях скучивание животных обеспечивает сниженный энергетический расход, а следовательно, лучшие условия выживания во время прекращения питания в зимний период. Большие зимние скопления грызунов в копнах сена и соломы (полевая мышь — Apodemus agrarius, курганчиковая мышь — Mus musculus hortulanus и др.) также способствуют лучшему выживанию в этих резервациях. Ограничение газообмена в группе мелких видов может достигать 50% по сравнению с одиночными особями.
Изменение газообмена у изолированных животных и в группе
(по Понугаевой, 1960)
| Отряд | Жив >тное | Температура камеры CC | Потребление O2 (м.г,кгіч) | Обмен веществ (в °Л от Гб- мена одиночек) | Вероятность случайности (%) | ||
| у изолированных ЖИВОТНЫХ | в группе | ||||||
| Руко крылые | Подковонос малый (в покое) (Rhinolop- hus hipposidcros) . . | 23 | 1169 | 838 | 72 | ||
| Ночница большая (в покое) (Myotis myotis)............................ | 14 | 4366 | 2364 | 54 | |||
| Длиннокрыл обыкновенный (в покое) (Miniopterus schrei- bersi).................. ....... | 23 | 1846 | 1084 | 59 | |||
| Насеко- моя тные | Еж ушастый (Не- miechinus auritus) . . | 16 | 1698 | 2280 | 134 | 95 | |
| Грызуны | Полевка обыкно венная (Microtus аг- valis) | 20 | 5689 | 4196 | 73,8 | 99 | |
| Полевка общест венная (Microtus so- cialis) | 25 | 6218 | 3756 | 60,4 | 99 | ||
| Полевка рыжая (Clethrionomys glareolus) | 22 | 2366 | 2771 | 117 | 99 | ||
| Крыса пасюк (серая) (Rattus norvegi- cus) | 20 | 4615 | 2529 | 54,8 | 99 | ||
| Крыса пасюк альбинос (Rattus norvegi- cus var. albino . . . | 20 | 3413 | 2704 | 79,2 | 95 | ||
| Незокия (Nesokia indica) | 16 | 1390 | 1375 | 98,5 | 95 | ||
| Мышь домовая | (20 | 4412 | 4061 | 92 | 96 | ||
| (Mus musculis) . . | (20 | 4173 | 4127 | 98,8 | 95 | ||
| Мышь белая . . . | 22 | 8168 | 6309 | 77,2 | 99 | ||
| Соня лесная (Dyro- mys nitedula) . . . . | 25 | 3767 | 3166 | 84 | 99 | ||
| Отряд | Жипотное | Температура камеры (rC) | Потребление O2 (мл/кг,ч) | Обмен веществ (в % от об мена одиночек) | Вероятность случайности (%) | ||
| у изолированных ЖИВОТНЫХ | в группе | ||||||
| Пеструшка степная (Lagurus lagurus) . . | 22 | 7537 | 6802 | 90 | 95 | ||
| Хомячок серый {Cricetulus migrato- rius) | 23 | 3079 | 3728 | 121 | 98 | ||
| Хомяк крысовид- | US | 1568 | 1609 | 102,2 | 95 | ||
| ный {Cricetulus triton) | 118 | 1846 | 1452 | 78 | 99 | ||
| Песчанка мало- азийская {Meriones | 125 | 2504 | 1540 | 61,5 | 99 | ||
| tristrami.......................... | 1 25 | 3789 | 2153 | 65,5 | — | ||
| Суслик тонкопалый {Spermophilopsis lep- | /20 | 1062 | 1040 | 97,9 | 95 | ||
| todactylus)...................... | 120 | 888 | 787 | 89,2 | 95 | ||
| Суслик малый {Сі- | 125 | 2807 | 1484 | 52,8 | 99 | ||
| tellus pygmaeus) . . | 125 | 2648 | 1476 | 55,7 | 99 | ||
| Хищники | Норка {Mustela lut- | / 13 | 2972 | 2336 | 78,6 | 98 | |
| reola) ....... | 1 із | 3360 | 2356 | 70,1 | 99 | ||
| Кошка домашняя . | 20 | 1066 | 751 | 70,5 | 99 | ||
| Обезья- | Зеленая мартышка | ||||||
| мы | (Cercopithecus griseo- viridis) | 28 | 579 | 632 | 108,9 | 99 | |
| Павиан-гамадрил {Papio hamadrylas) | 24 | 689 | 475 | 68,9 | 99 | ||
Однако описанные выше рефлекторные изменения газообмена проявляются далеко не во всяком физиологическом состоянии. Наибольший эффект удается наблюдать на группах, состоящих из особей одного пола, или на разнополых особях вне периодов спаривания (гона). В периоды спаривания «стадный» эффект резко уменьшается или исчезает, заменяясь значительным повышением обмена при совместном пребывании однополых
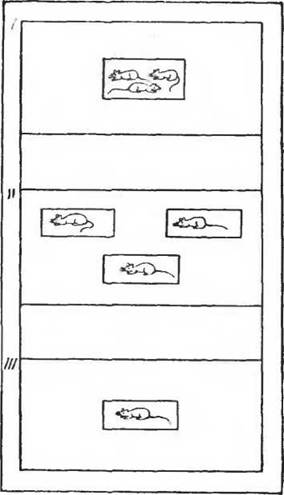
Рис. 85. Схема постановки опытов с изучением стадных рефлексов на газообмен. I—группа; II—разделенная группа; III — одиночка
особей, что особенно ярко проявляется у малого суслика в весеннее время.
Специальной формой эксперимента, когда животные помещались в отдельные клеточки в общей камере, удалось установить наличие стадного эффекта. Однако когда часть животных была заменена источниками теплового излучения равной интенсивности, то стадный эффект оставшегося в камере животного исчез. Следовательно, изменения газообмена не могут быть сведены к процессам ограничения теплоотдачи, т. е. к процессам терморегуляции (рис. 85).
Физиологические сдвиги, происходящие в группе себе подобных организмов, не ограничиваются только изменением газообмена. Например, в группе овец можно наблюдать учащение пульса и дыхания при удалении других овец. У взрослых овец этот эффект выражен ярче, чем у ягнят (Понугаева и Трубицына, 1955). Так, частота сердечных сокращений у новорожденного ягненка, оставшегося без овцы-матки на 30—40 лшн, увеличивалась на 5 ударов у серой каракульской породы, на 12 у черных и на 14 у гибридов. В момент удаления матки у серых пульс учащался на 17, у черных на 13 и у гибридов на 2 удара; при приближении матки у серых пульс повышался на 3 удара, у черных на 9, у гибридов архарокаракуль на 12. Изменения дыхания были незначительны. На 5-й день после рождения характер изменения пульса и дыхания остался тем же (изучалась реакция только у гибридов). Так, у находящегося с овцой-маткой ягненка частота пульса была 218 ударов, во время ухода матки частота пульса увеличилась до 256 ударов. Когда ягненок остался один, пульс понизился до 200 ударов, при приближении овцы-матки увеличился до 238.
Анализаторы, раздражение которых вызывает описанные эффекты, по-видимому, достаточно разнообразны и до сих пор у грызунов изучены мало; вероятно,
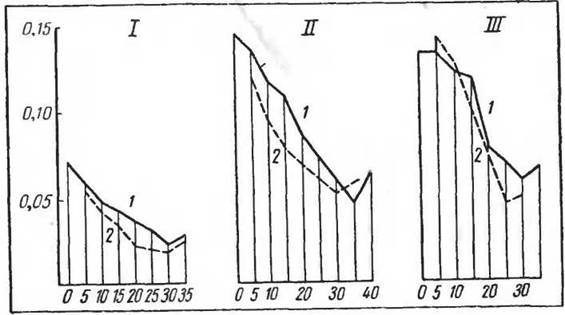
Рис. 8G. Стадные изменения обмена веществ при различной температуре внешней среды (по Понугаевой, I960)./ — песчанка; II— общественная полевка; III — обыкновенная полевка:
/ — одиночные животные, 2 — животные в группе. По оси ординат — потребление Ог (мл'миніг)' по оси абсцисс — температура среды
все анализаторы принимают участие при изменении реакции газообмена в группе. Установлена роль зрительного, обонятельного и, по-видимому, звукового анализаторов. Опыт показывает, что если этих животных рассадить по отдельным камерам, имеющим прозрачные стенки, так, чтобы общение осуществлялось только за счет звукового и зрительного анализаторов, а обонятельный будет выключен, то в первое время стадная реакция исчезает, но через некоторое время вновь обнаруживается. Очевидно, нарушенный динамический стереотип через некоторое время восстанавливается на основе других анализаторов. Однако здесь имеют место большие видовые различия.
Чрезвычайно важен вопрос о роли температурного анализатора в изменениях обмена веществ в стаде. Из рисунка 86 видно, что эти изменения наиболее ярко вы-
ражаются при низких температурах среды, при средних они уменьшаются, а при высоких сравнительно невелики, но не исчезают полностью. Следовательно, стадный эффект имеет много общего с «групповой» терморегуляцией, но не аналогичен ей.
Стадные изменения обмена веществ обнаруживаются не только у бодрствующего (гомойотермного) животного, и во время зимней спячки. Так, у спящей группы летучих мышей газообмен оказывался пониженным по сравнению с одиночной особью в том же состоянии, при той же температуре среды и температуре тела.
Имеются немногочисленные данные о физиологических механизмах, лежащих в основе наблюдаемых изменений газообмена. Роль щитовидной ?келезы в проявлении стадного эффекта у белых крыс изучалась путем подавления ее функции при введении в организм р-ме- тилтиоурацила — 25,35 и 50 мг ежедневно. Во всех случаях наблюдалось понижение газообмена в зависимости от дозы. Однако на фоне этого сниженного уровня наблюдался отчетливо выраженный стадный эффект (около 15% снижения обмена в группе). Кроме того, были поставлены исследования с полной одномоментной экстирпацией щитовидной железы. В ряде случаев наблюдалось увеличение стадного эффекта на фоне выключения (полного и частичного) функции щитовидной железы. Особенно яркий эффект получен при очень низкой температуре среды (7°С в летний период). Здесь стадный эффект почти отсутствовал у нормальных крыс и составлял около 15% снижения обмена у крыс, получавших метилтиоурацил. Таким образом, в отличие от специфического динамического действия пищи, выпадающего при угнетении функции щитовидной железы, стадный эффект при этом сохраняется и даже становится более значительным.
Снижение газообмена в 70% случаев сопровождается снижением величины электрической активности скелетной мускулатуры. Есть все основания предполагать, что в основе наблюдаемых изменений газообмена лежат сдвиги в состоянии мышечного тонуса, изменения возбуждения мышечных волокон скелетной мускулатуры. Каковы при этом изменения в других органах и в частности во внутренних органах и в центральной нервной системе, должны показать будущие исследования.
Стадные и групповые реакции связаны с эндокринными системами. В группе животных постоянно действуют факторы, которые ^ерез нервную систему вызывают изменения в функции эндокринных желез, особенно гипо- физарно-адренокортикальной и репродуктивной систем (Christian, 1963). В основе этих явлений лежит реакция стресса, т. е. неспецифических стадий адаптации, особенно ярко выраженных при экспозиции животных в группе на сравнительно короткое время. В опытах Кристиана подсаживание полевки Microtus agrestis в клетку, где уже жила пара полевок этого же вида, приводило к борьбе, покусам животных и к значительной гипертрофии надпочечников и селезенки, атрофии тимуса, увеличению межпозвоночных хрящей, т. е. к явлениям стресса, что было ярче выражено у подсаженных особей, нежели у тех, которые жили здесь ранее.
При сопоставлении числа рубцов от укусов (в процессе борьбы) с перечисленными выше морфофнзиологиче- скнми изменениями в организме; никакой связи между этими явлениями установить не удалось (Clarke, 1953; Chitty a. oth., 1956; Christian, 1959). Таким образом, физиологические изменения в группе не связаны пи с мышечной деятельностью, пи с травмами, которые животные получают в процессе борьбы.
Физиологические механизмы, лежащие в основе явлений стресса, возникающего в этих условиях, были тщательно изучены и описаны Кристианом (Christian, 1963). Они заключаются в возбуждении гипофизо-адренокорти- кальной системы, усилении распада белков, подавлении роста костей, соединительной ткани, продукции коллагена, в подавлении митотической активности, что связано и со снижением выделения гормона роста. Повышение активности коры надпочечников (выделение кортикои- дов) может оказывать угнетающее влияние и на щитовидную железу. Однако, по-видимому, снижение обмена веществ в группе связано не только с этим угнетением тиреоидальной функции, а и с изменением состояния скелетной мускулатуры и особенно с двигательным поведением животного.
Изучение ориентировочно-исследовательской реакции по «поведению на открытой площадке» (open field behaviour) показало, что изолированное содержание белых крыс ведет к снижению ориентировочно-исследова-
тельской активности по сравнению с белыми крысами, содержащимися в группе. Ориентировочно-исследовательская активность как у изолированных, так и у содержащихся в группе животных несколько выше в зимний период (январь—февраль), чем в весенне-летний.
Меняется в зависимости от сезона и активность гипофизарно-надпочечниковой системы. Однако направленность этих изменений различна у «групповых» и изолированных животных (Старыгин, 1969). В зимнее время уровень кортикостерона в крови у содержащихся в группе животных в 2—2,5 раза выше, чем у изолированных. С наступлением весны активность гипофизарно-надпочечниковой системы у изолированных животных повышается, а у групповых несколько снижается. В результате в мае уровень кортикостерона у групповых животных оказывается в 2 раза ниже, чем у изолированных.
Еще по теме СТАДНЫЕ И АТРЕГАПИОПНЫЕ РЕАКЦИИ:
- Сравнительный анализ больных ОГ с положительной реакцией Манту на 2 ТЕ и 5 ТЕ с больными подгруппы основной группы с отрицательной реакцией Манту на 2 ТЕ и положительной РМ на 5 ТЕ
- Зрачковые реакции
- Общие реакции организма на повреждение
- 9. Психогенные реакции в детском возрасте (К. Левикова).
- VI. Классификация аллергических реакций.
- Гиперсенситивные реакции
- Поствакцинальные реакции и их купирование
- 23. Патологические реакции и их виды.
- Специфические аллергические реакции
- Физиологические реакции на стресс
- 1. Ориентировочная реакция
- Реакция гемолиза
- Глава 11. Анафилактические реакции
- Аллергические реакции IV типа
- Реакция связывания комплемента
- Компенсаторные реакции при гипоксии
- Реакция с липоидным антигеном (VDRL).