3.1. Морфология миндалевидного тела головного мозга крыс контрольных групп
В соответствии с целью и задачами работы вначале была изучена топография и морфология МТ головного мозга крыс контрольных групп.
При исследовании топографии МТ нами, в соответствие с рекомендациями С.А.
Чепурнова и Н.Е. Чепурновой [191], были выделены базо-латеральный и кортико-медиальный отделы, которые включали следующие ядра. Базо-латеральный отдел состоял из 2-х ядер: базального и латерального. Кортико-медиальный отдел включал 3 ядра: кортикальное, медиальное и центральное (рис. 3.1).



|
|
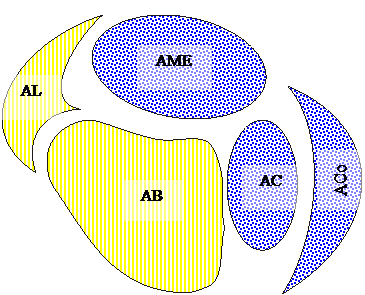
Рис. 3.1. Схема ядер миндалевидного тела головного мозга (по С.А. Чепурнову и Н.Е. Чепурновой, 1981; с изменениями):
BL – базо-латеральный отдел: АВ – базальное ядро ; AL – латеральное ядро.
; AL – латеральное ядро.
КМ – кортико-медиальный отдел: АСо – кортикальное ядро; АМЕ – медиальное ядро; АС – центральное ядро.
Все цифровые данные, полученные при изучении МТ головного мозга контрольных животных 2-х возрастных групп, приведены в таблице 3.1.
Таблица 3.1
Морфометрические характеристики клеток ядер основных отделов миндалевидного тела головного мозга крыс разного возраста контрольных групп (М ± m) (мкм)
| Возраст крыс | Отдел МТ | Изучаемый параметр | Сроки эксперимента (сутки) | |||
| 7 | 15 | 30 | 60 | |||
| I | BL | Sкл | 59,67±0,03* | 45,71±0,03* | 57,92±0,07* | 59,11±0,2* |
| Sяд | 4,11±0,01 | 4,04±0,01 | 3,66±0,02 | 3,73±0,07 | ||
| ЯЦО | 0,08±0,001* | 0,11±0,002* | 0,08±0,003* | 0,07±0,01* | ||
| Н | 2142,05±2,38* | 2088,17±2,09 | 2077,96±6,36 | 2000±29,44 | ||
| Г | 1913,04±4,25 | 2445,16±2,74 | 2241,93± 13,62* | 2431,09± 63,99 | ||
| С | 788,21±2,81* | 1202,76±2,09* | 1044,35± 10,72* | 1032,26± 9,55* | ||
| Г/Н | 0,89±0,052* | 1,17±0,03 | 1,08±0,16 | 1,22±0,79 | ||
| С/Н | 0,37±0,04 | 0,58±0,02* | 0,5±0,14 | 0,52±0,66 | ||
| С/Г | 0,41±0,02 | 0,49±0,02* | 0,47±0,06* | 0,42±0,29* | ||
| КМ | Sкл | 66,16±0,04 | 57,14±0,03* | 61,22±0,07 | 66,3±0,28 | |
| Sяд | 4±0,01 | 4,04±0,01 | 4,31±0,02 | 3,94±0,07 | ||
| ЯЦО | 0,08±0,002 | 0,11±0,002* | 0,08±0,003 | 0,07±0,009 | ||
| Н | 2159,48±2,25 | 3380,38±2,42* | 2650,54±6,62 | 2274,19±16,2 | ||
| Г | 2610,99±1,84 | 2018,85±1,94 | 2044,23±12,41 | 2361,78±5,85 | ||
| С | 1736,99±1,61* | 2051,29±1,92* | 1551,09±2,11 | 1612,9±6,75* | ||
| Г/Н | 1,21±0,06 | 0,6±0,01 | 0,77±0,14 | 1,04±0,42 | ||
| С/Н | 0,8±0,05 | 0,61±0,03 | 0,59±0,12* | 0,71±0,32 | ||
| С/Г | 0,67±0,03* | 1,02±0,02* | 0,76±0,05* | 0,68±0,1* | ||
| II | BL | Sкл | 75,71±0,1* | 64,95±0,08* | 73,54±0,11* | 72,9±0,08* |
| Sяд | 4,16±0,02 | 3,93±0,02 | 3,64±0,02 | 3,62±0,02 | ||
| ЯЦО | 0,06±0,003* | 0,07±0,003* | 0,06±0,003* | 0,06±0,002* | ||
| Н | 1526,82±2,35* | 1811,76±4,65 | 1561,24±2,89 | 1423,94±2,77 | ||
| Г | 1657,26±2,39 | 2361,13±3,09 | 2236,67±3,64* | 1894,92±3,85 | ||
| С | 809,14±3,11* | 829,09±3,39* | 791,68±3,61* | 687,6±3,09* | ||
| Г/Н | 1,09±0,08* | 1,3±0,1 | 1,43±0,11 | 1,33±0,11 | ||
| С/Н | 0,53±0,07 | 0,46±0,07* | 0,51±0,08 | 0,48±0,08 | ||
| С/Г | 0,49±0,06 | 0,35±0,06* | 0,35±0,07* | 0,36±0,06* | ||
| КМ | Sкл | 63,6±0,11 | 67,26±0,08* | 63,6±0,1 | 70,92±0,08 | |
| Sяд | 4,05±0,03 | 4,76±0,03 | 4,23±0,04 | 3,95±0,02 | ||
| ЯЦО | 0,07±0,005 | 0,08±0,004* | 0,08±0,006 | 0,06±0,002 | ||
| Н | 1526,82±4 | 1911,1±4,83* | 1561,24±4,83 | 1410,14±2,89 | ||
Продолж.
табл. 3.1| Возраст крыс | Отдел МТ | Изучаемый параметр | Сроки эксперимента (сутки) | |||
| 7 | 15 | 30 | 60 | |||
| II | КМ | Г | 790,32±3,62 | 2437,85±3,31 | 2361,29±4,84 | 1658,99±2,63 |
| С | 265,48±3,44* | 848,82±2,21* | 909,68±3,33 | 668,2±2,12* | ||
| Г/Н | 0,52±0,13 | 1,28±0,16 | 1,51±0,07 | 1,28±0,08 | ||
| С/Н | 0,17±0,08 | 0,44±0,07 | 0,58±0,05* | 0,47±0,05 | ||
| С/Г | 0,34±0,06* | 0,35±0,05* | 0,39±0,05* | 0,4±0,05* | ||
Примечание: возраст крыс: I – неполовозрелые, II – пубертатный период;
отделы МТ: BL – базо-латеральный, KM – кортико-медиальный;
обозначения изучаемых параметров клеток представлены на стр. 38-39;
* – статистически достоверные изменения показателей.
Базо-латеральный отдел МТ. В 2-х возрастных группах животных чаще других определяются нейроны средних размеров треугольной и вытянуто-овальной формы. Ядра располагаются в центральной части нейрона, несколько в сторону от его геометрического центра. У ядер четко выражена оболочка. Содержимое ядра имеет мелкозернистую структуру. Внутри ядра определяется обычно одно темное ядрышко. В цитоплазме нервных клеток выявлялось крупнозернистое хроматофильное вещество, равномерно расположенное. В аксоне и его конусовидном основании хроматофильное вещество отсутствует. Глиальные элементы располагаются диффузно (рис. 3.2). При исследовании данного отдела МТ головного мозга крыс пубертатного периода нами визуально не было выявлено существенных морфологических отличий в строении изучаемой структуры, по сравнению с неполовозрелыми животными. При морфометрическом исследовании установлено, что нейроны 2-х ядер данного отдела исследуемых возрастных групп расположены компактно, на расстоянии 2-11 мкм друг от друга.
Площадь клеток составляла в среднем 55,6 ± 0,08 мкм2 для неполовозрелых животных и 71,78 ± 0,09 мкм2 – для половозрелых. Статистически достоверные изменения изучаемых параметров клеток наблюдались лишь в значениях показателей площади сечения нейрона в
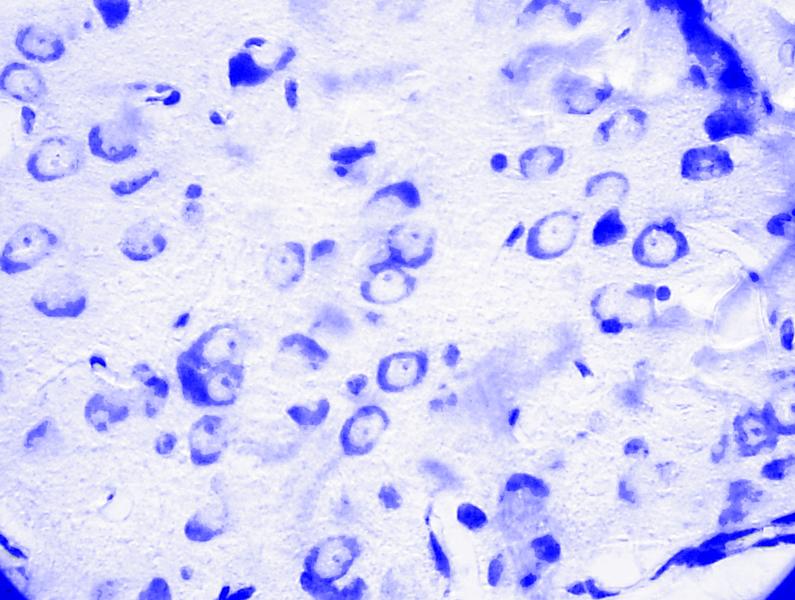
Рис. 3.2. Микрофотография. Латеральное ядро базо-латерального отдела миндалевидного тела головного мозга половозрелой крысы на 7 сутки. Контрольная группа. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
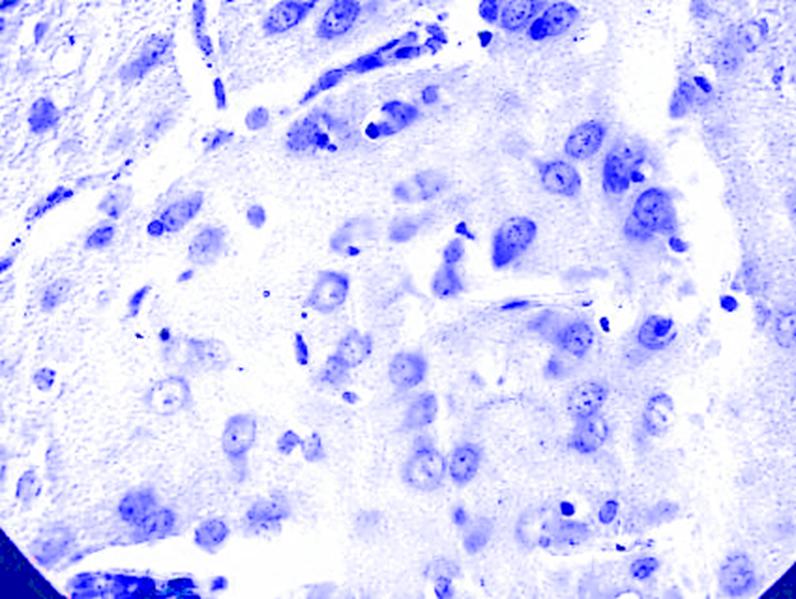
Рис. 3.3. Микрофотография. Медиальное ядро кортико-медиального отдела миндалевидного тела головного мозга неполовозрелой крысы на 7 сутки. Контрольная группа. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
сторону увеличения последних с возрастом, а показателей ядерно- цитоплазматического отношения, плотности сателлитной глии и интерглиального коэффициента в сторону уменьшения (табл. 3.1).
Кортико-медиальный отдел МТ. В 2-х возрастных группах животных в указанном отделе чаще других определяются нейроны средних размеров. Состоит из клеток вытянуто-овальной формы, равномерно распределенных в объеме исследуемого отдела. В клетках – округлое ядро средней величины располагается в центральной части нейрона, несколько в сторону от его геометрического центра. Одиночные ядрышки расположены в центре ядра. У ядер четко выражена оболочка. Содержимое ядра имеет мелкозернистую структуру. Крупнозернистое хроматофильное вещество, равномерно расположено в соме клетки. Глиальные элементы располагаются диффузно (рис. 3.3).
Нейроны 3-х ядер МТ 2-х возрастных групп расположены на расстоянии 7-16 мкм друг от друга. При микроскопической морфометрии нами было выявлено, что площадь клетки составляет 62,71 ± 0,11 мкм2 для неполовозрелых животных и 66,35 ± 0,09 мкм2 – для половозрелых.
При морфометрическом исследовании данного отдела нами не выявлены существенные морфологические отличия в строении клеток 3-х ядер данного отдела МТ животных разных возрастных периодов. Лишь в значении показателей плотности сателлитной глии и интерглиального коэффициента наблюдалось их статистически достоверное уменьшение у крыс пубертатного возраста по сравнению с таковым у неполовозрелых животных. Изменения других 7 изучаемых показателей не имели статистически достоверных различий, то есть были практически одинаковы (табл.3.1).При сравнении использованных нами в работе 9-и цифровых показателей клеток и ядер в целом 2-х разных отделов МТ между собой статистически достоверные изменения установлены только по показателям плотности общей глии и сателлитной глии в сторону увеличения их в кортико-медиальном отделе в 2-х возрастных группах.
Электронная микроскопия. Данному материалу было уделено больше внимания, так как в имеющейся литературе сведения по МТ головного мозга человека и животных крайне ограничены и скудны. Электронно-микроскопический анализ образцов клеток ядер изучаемых отделов МТ свидетельствует об их сложной микроскопической организации, представленной нервными и глиальными клетками с их многочисленными отростками, формирующими нейропиль, пронизанный сетью терминальных сосудов.
Нейроны содержали крупное овальное ядро с равномерно распределенными гранулами хроматина и электронноплотным ядрышком, локализованным чаще в центре ядра. Ядерные мембраны имели ровный контур, перинуклеарное пространство узкое, немногочисленные ядерные поры были хорошо выражены. Наружная кариолемма содержала рибосомы, наблюдалась органичная связь ее с канальцами гранулярной эндоплазматической сети (ЭПС), представленной короткими цистернами. В цитоплазме были расположены также свободные рибосомы и немногочисленные гранулы гликогена. Агранулярная ЭПС была представлена мелкими везикулами и локализована в перикарионе вблизи комплекса Гольджи и топологически связана с цистернами гранулярной сети.
Аппарат Гольджи был расположен вблизи ядра и образован уплощенными цистернами в комбинации с разнообразными пузырьками. Цитоплазма нейронов содержала также значительное количество лизосом 0,3-0,5 мкм в диаметре округлой или овальной формы с мелкозернистым содержимым средней электронной плотности, окруженных одиночной элементарной мембраной (рис. 3.4).Митохондрии нервных клеток широко варьировали по своей форме и размерам. В перикарионе они имели округлую или овальную форму, в отростках – палочкообразную и вытянутую с диаметром 0,1 мкм. В цитоплазме нейронов митохондрии располагались хаотично, связь этих органелл с канальцами гранулярной ЭПС не наблюдалась (рис. 3.4, 3.5). Матрикс митохондрий умеренной электронной плотности, наружная мембрана гладкая,
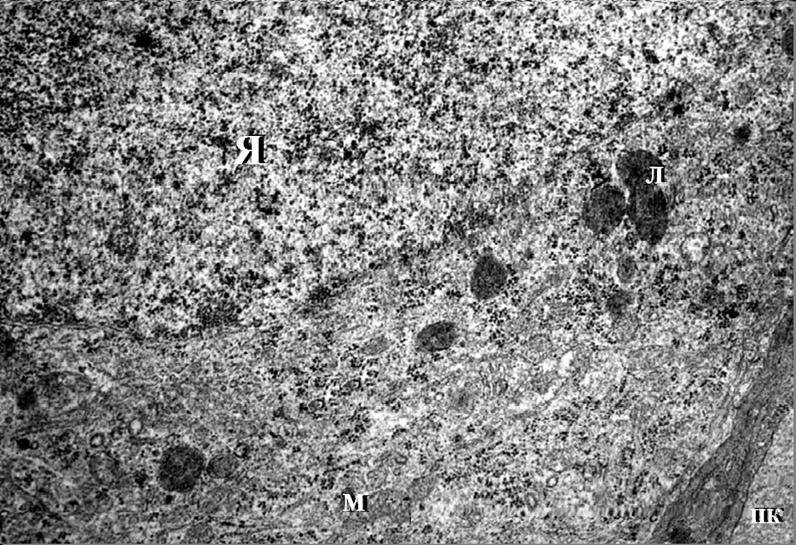
Рис.3.4. Электронная микрофотография. Ультраструктура нейрона кортикального ядра кортико-медиального отдела миндалевидного тела головного мозга неполовозрелой крысы. Контрольная группа. Увеличение 15000.
Я – ядро;
М – митохондрии;
ПК – просвет капилляра;
Л – лизосомы.
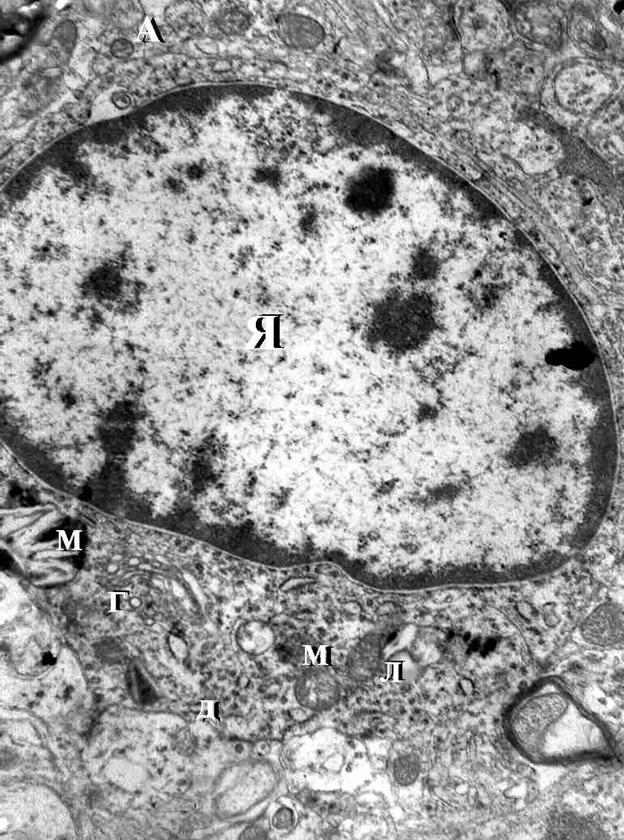
Рис.3.5. Электронная микрофотография. Ультраструктура клетки базального ядра базо-латерального отдела миндалевидного тела головного мозга крысы пубертатного возраста. Контрольная группа. Увеличение 8000.
Я – ядро;
М – митохондрии;
Л – лизосомы;
Г- аппарат Гольджи;
А – аксон;
Д – дендрит.
внутренняя формировала кристы поперечной ориентации. В отростках нейронов удлиненные митохондрии иногда содержали продольно ориентированные кристы. В перикарионе нейронов между органеллами наблюдались микротрубочки и нейрофиламенты, образующие рыхлые пучки, проходящие в начальные сегменты аксонов и дендритов.
Нейропиль был образован тонкими немиелинизированными терминалями аксонов и дендритов, многие из которых образуют между собой и телами нейронов аксодендритические и аксосоматические контакты.
В нейропиле наблюдались отростки олиго- и астроглии, миелинизированные аксоны, тела клеток микроглии.Глиальные клетки различались, как своими морфологическими характеристиками, так и локализацией относительно других клеточных элементов МТ. При этом клетки астроглии были равномерно распределены среди нервных клеток, образуя расширения, лежащие на поверхности кровеносных сосудов («концевые ножки»). Астроциты имели округлое или овальное ядро с кариоплазмой равномерной плотности. Ядерная оболочка иногда образовывала глубокие складки. Цитоплазма относительно бедна органеллами, расположенными в основном в перикарионе. Свободных рибосом мало, ЭПС была представлена короткими цистернами. Митохондрии разнообразны по форме, в отростках они вытянуты по длине. Отличительной чертой астроцитов являлось наличие в цитоплазме их тел и отростков фибрилл. Крупные отростки содержали те же органеллы, что и перикарион. В более тонких отростках большинство органелл отсутствовали. Здесь наблюдались лишь единичные рибосомы, гранулы гликогена, микротрубочки, пучки фибрилл. Очень часто тончайшие отростки этих клеток приобретали форму пластинок, где расстояние между плазматическими мембранами составляло несколько сот ангстрем. Такие пластинчатые отростки нередко лежали вокруг синапсов. Отростки астроцитарной глии формировали целый ряд специализированных контактов друг с другом: плотные контакты и точечные сращения. Вокруг кровеносных сосудов концевые ножки астроцитов образовывали сравнительно плотный слой, принимая участие в структуре и функционировании гематоэнцефалического барьера. В олигодендроцитах был хорошо развит аппарат Гольджи, цистерны и везикулы которого повсеместно встречались в цитоплазме перикариона. Митохондрии многочисленные небольших размеров с поперечными кристами. Цитоплазма олигодендроцитов содержала включения: тельца либо с гомогенным содержимым, либо состоящие из гранул, филаментов, мембранных структур. Клетки микроглии имели небольшие размеры (4-5 мкм), сосредоточены вокруг сосудов и нейронов и контактировали с ними. Электронноплотное ядро занимало почти весь объем этих клеток. Цитоплазма образовывала узкий слой вокруг ядра и содержала митохондрии, цистерны гранулярной ЭПС и рибосомы. Олигодендроциты, имеющие умеренную электронную плотность, часто являлись сателлитами нейронов. Ядра их отличались полиморфизмом – от округлых до вытянутых ядер неправильной формы с мелкими глыбками гетерохроматина. Перинуклеарное пространство расширено. Ядро часто расположено эксцентрично. В цитоплазме была хорошо развита гранулярная ЭПС, цистерны которой локализировались концентрически вокруг ядра. Цитоплазматический матрикс отличался высокой электронной плотностью.
Для более глубокого изучения морфологии различных отделов МТ головного мозга крыс и с целью дальнейшего использования полученных данных определялись корреляционные взаимосвязи между ядрами, а затем и отделами МТ в контрольных группах животных. В итоге корреляционная плеяда для каждой контрольной группы включает 2 отдела МТ головного мозга и 9 изучаемых характеристик клеток. Анализ корреляционной плеяды показал следующее.
1. Наиболее сильная связь (0,7 ≤ r ≤ 1,0) в обеих возрастных группах контрольных животных наблюдалась между обоими отделами МТ по следующим 5 показателям: площадь сечения нейрона; площадь сечения ядра; плотность нейронов на 1 мм2; плотность общей глии на 1 мм2; плотность сателлитной глии на 1 мм2.
2. По остальным 4 показателям коэффициенты корреляции составляют среднюю степень связи (0,3 ≤ r ≤0,7) между отделами МТ.
Таким образом, можно отметить, что изучаемые отделы МТ головного мозга крыс представляют собой в контроле довольно устойчивую систему с высоким уровнем взаимосвязей ее компонентов между собой по их основным характеристикам.
Еще по теме 3.1. Морфология миндалевидного тела головного мозга крыс контрольных групп:
- 3.4. Морфологические преобразования миндалевидного тела головного мозга крыс после хронической интоксикации фенобарбитоном с коррекцией силибором
- 3.3. Морфологические преобразования миндалевидного тела головного мозга крыс неполовозрелого возраста после хронической интоксикации бензоналом
- 3.5. Однофакторный дисперсионный анализ хронического влияния барбитуратов на ядра основных отделов миндалевидного тела головного мозга крыс в различные возрастные периоды
- 3.2. Морфологические преобразования миндалевидного тела головного мозга крыс после хронической интоксикации фенобарбитоном в постнатальном периоде
- 1.2. Морфофункциональные преобразования миндалевидного тела головного мозга человека и животных после интоксикации барбитуратами
- 4.3. Структурные изменения головного мозга у детей с церебральными параличами по результатам компьютерной томографии головного мозга.
- Уровень эндогенной интоксикации и активность процессов свободнорадикального окисления в контрольной группе и группе больных миомой матки
- 3.4. Особенности психоэмоционального статуса у военнослужащих-женщин с ожирением и в контрольной группе в период беременности 3.4.1. Оценка качества жизни и уровня тревожности в исследуемых группах
- 3 группа (контрольная).
- 3 группа (контрольная).
- 1.1 Результаты применения МР-диффузии в контрольной группе пациентов
- 3 группа (контрольная).
- Клиническая характеристика основной и контрольной групп
- 3 группа (контрольная).