Механизм действия Са2+-АТФазы
Рассмотрим основные принципы работы ионных насосов на примере Са2+-АТФазы саркоплазматического ретикулума (SR) клеток
мышц (SR Са2+-АТФаза) (рисунок 64).
Этот фермент составляет 80% белков саркоплазматического ретикулума и играет ключевую роль в реализации сокращения клеток мышц, которое стимулируется ростом концентрации ионов кальция в цитозоле.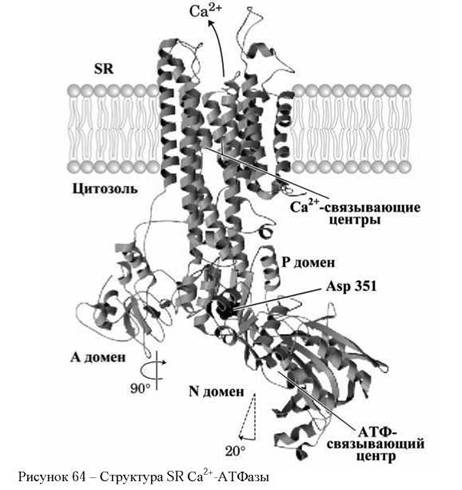
Релаксация мышц происходит вследствие уменьшения концентрации кальция в цитозоле и переноса его в саркоплазматический ретикулум (специализированный "отсек" гладкого эндоплазматического ретикулума для хранения ионов кальция), который осуществляется SR Са2+-АТФазой.
Концентрация ионов Са2+ в цитозоле клеток мускулатуры изменяется от значений 10 7 М (в расслабленном состоянии) до более чем 10 6 М (при сокращении), а в саркоплазматическом ретикулуме концентрация Са2+ может достигать 10 2 М.
Са2+-АТФаза - это 110 кДа полипептид, трансмембранный домен которого состоит из десяти а-спиралей. (1 Дальтон (Да) = 1 а.е.м = = 1,00-10 21 г). Цитоплазматическая часть SR Са2+-АТФазы, по массе составляющая приблизительно половину молекулярной массы белка, состоит из трёх доменов А, N и Р (рисунок 64).
Три цитоплазматических домена SR Са2+-АТФазы имеют различные функции. Нуклеотид-связывающий домен N связывает АТФ, фосфо- рилируемый домен Р принимает фосфатную группу на аспартат Asp 351, активаторный (управляющий) домен А активизирует домен N.
Десять трансмембранных а-спиралей формируют канал для прохождения Са2+ через мембрану. Две из этих спиралей доходят только до середины мембраны и те места, в которых белковая нить становится спиралью, являются центрами связывания ионов кальция.
Пространственное положение домена А управляет сродством ионов Са2+ к этим связывающим центрам и последующим выходом ионов кальция из цитозоля наружу из клетки или в саркоплазматический ретикулум клетки.
В исходном состоянии между центром фосфорилирования и центрами связывания Са2+-ионов достаточно большое расстояние. В процессе одного транспортного цикла домен N наклоняется на 20° влево (рисунок 61), перемещая АТФ-центр к аспартату Asp 351 (аспарагиновая кислота), и домен А поворачивается на 90° вокруг нормали к мембране.
Такие изменения конформации молекулы приводят к перемещению Са2+-связывающих центров сначала на одну сторону мембраны, а затем на другую, изменяя при этом сродство этих центров к ионам Са2+ от высокого на цитоплазматической стороне мембраны до низкого на саркоплазматической стороне мембраны.
Общим для работы всех насосов является
1) фосфорилирование специфического аспартата - Asp 351 в случае SR Са2+-АТФазы;
2) существование как минимум двух различных конформаций, которые мы обозначим и /І .
С учётом фосфорилирования, следовательно, существует минимум четыре конформационных состояния Ех, Д-Р, Д-Р, Д, на основе которых можно построить общую схему работы насосов (рисунок 65).
Реакционный цикл работы насоса состоит из шести этапов.
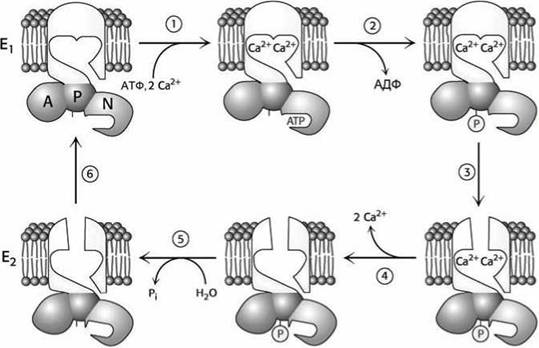
Рисунок 65 - Схема механизма действия SR Са2+-АТФазы: связывание Са2+ (1) и фосфорилирование АТФазы (2) приводит к переносу (3) центров связывания с цитозольной на внешнюю сторону мембраны и высвобождению Са2+ (4); гидролиз фосфоаспартата (5) и конформационный переход, переносящий центры связывания обратно на внутреннюю сторону мембраны (6). возвращает АТФазу в исходное состояние
1. Цикл начинается со связывания АТФ и двух ионов Са2+ с конформацией Д АТФазы.
2. АТФаза переносит фосфатную группу к целевому аспартату.
Кальций уже должен быть связан ферментом, чтобы моглопроходить фосфорилирование. Фосфорилирование смещает кон- формационное равновесие АТФазы в сторону Е2 -конформации.
3. Переход от Д к Ег конформации приводит к "выворачиванию" через мембрану ион-связывающих центров так, что диссоциация ионов кальция будет происходить уже на наружной стороне мембраны.
4. В Е2 -конформации АТФаза имеет низкое сродство к ионам Са2+, что приводит к их высвобождению.
5. Высвобождение ионов Са2+ стимулирует гидролиз фосфоаспартата (дефосфорилирование) и диссоциацию фосфатной группы.
6. АТФаза, лишённая ковалентно связанной фосфатной группы, конформационно нестабильна в Е2 состоянии. Она "выворачивается обратно" в /', -конформацию, завершая реакционный цикл.
Все ионные насосы P-класса независимо то того, какие ионы они перекачивают через мембрану, имеют подобное строение. Во всех таких насосах происходит фосфорилирование аспартата, трансмембранные а-домены всех насосов P-класса имеют приблизительно одинаковый молекулярный вес и одинаковую "конструкцию" из а-спиралей. Всё это говорит о происхождении всех Р-насосов от общего предка, хотя и с течением времени эти насосы эволюционно приспособились транспортировать различные ионы.
Изменение цитозольной концентрации ионов Са2+ играет ключевую роль в механизмах клеточной сигнализации. Для того, чтобы быстро реагировать на сигнал, о котором внутриклеточные системы узнают из скачкообразного роста концентрации кальция в цитозоле, необходимо постоянно (в промежутках между сигналами) поддерживать низкую концентрацию ионов Са2+ в цитозоле (ниже 0,1-0,2 мкМ).
Ионы Са2+ удаляют из цитозоля Са2+-АТФазы плазматической мембраны клетки, строение которых подобно SR Са2+-АТФазе. Активность Са2+-АТФаз плазматической мембраны регулируется кальций-связывающими цитозольными белками кальмодулинами.
При высокой концентрации кальция в цитозоле кальмодулины, вследствие связывания с ионами кальция, изменяют конформацию и "обхватывают" Са2+-АТФазы, что, в свою очередь, индуцирует аллостерическую активацию АТФаз, в результате чего насосы быстро выкачивают ионы из цитозоля.
7.3.
Еще по теме Механизм действия Са2+-АТФазы:
- Механизм токсического действия
- Патогенез посттравматических когнитивных нарушений
- Организация синаптической передачи и ее роль в реализации когнитивных функций
- Нарушение синаптической передачи в патогенезе посттравматических когнитивных расстройств
- ПАТОГЕНЕЗ ГИПЕРТОНИЧЕСКОЙ БОЛЕЗНИ
- Тромбоз
- Гемолитическая анемия
- Механизм действия Са2+-АТФазы
- Механизм действия Na/K-АТФазы
- АВС-транспортёры
- 14.2. ИОНЫ КАЛЬЦИЯ
- СОДЕРЖАНИЕ
- Нарушение функций клеточных структур
- Механизмы нарушения барьерной функции биологических мембран
- Нарушение электрической стабильности липидного слоя