14.3. Осцилляции ВНУТРИКЛЕТОЧНОГО КАЛЬЦИЯ
Взаимодействие между Са2+-каналами плазматической и внутренних мембран клетки, кальциевыми насосами, а также кальций-связываю- щими белками, локализованными как в мембранах, так и в цитоплазме клетки, приводит к так называемым осцилляциям Са2+, то есть периодическим флуктуациям его концентрации в цитоплазме (рисунок 143).
В невозбудимых клетках основным триггером кальциевых осцилляций является инозитол трисфосфат, который образуется из фосфатидилинозитол бисфосфата при активации фосфолипазы С гормонами или факторами роста (рисунок 139). Инозитол трисфосфат способен диффундировать от плазматической мембраны, где он образуется, до мембран эндоплазматического ретикулума за десятки секунд. Количество и концентрация образующегося IP3 достаточно высоки, чтобы связаться со всеми соответствующими рецепторами, однако выход Са2+ происходит только в так называемых горячих участках.
Горячими называются участки переменной локализации в клетке, возникающие, как пузырьки в закипающей воде, в разных местах мембраны эндоплазматического ретикулума за счёт высокой локальной концентрации Са2+, инозитол трисфосфата или его рецептора.
Согласно существующим в настоящее время представлениям, вышедший в горячем участке в цитоплазму Са2+ диффундирует вдоль ретикулума и повышает в его мембранах чувствительность рецептора к инозитол трисфосфату, а также облегчает открывание канала, тем самым вызывая перемещение фронта Са2+-волны.
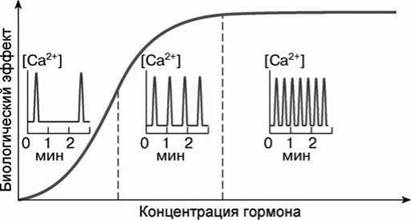
Рисунок 143 - Зависимость частоты флуктуации концентрации ионов Са2+ в цитоплазме и величины биологического эффекта, вызываемого ионами Са2+. от концентрации гормона
Локальное повышение концентрации Са2+ в этом участке мембраны выше порогового значения приводит к инактивации кальциевых каналов, в результате чего горячая точка гасится, а диффузия Са2+ генерирует новые точки выброса Са2+ из ретикулума.
В гашении горячих точек участвуют также кальциевые насосы, транспортирующие Са2+ из цитозоля в эндоплазматический ретикулум или межклеточное пространство.В возбудимых тканях основной вход Са2+ внутрь клетки происходит через потенциалочувствительные Са2+-каналы, которые функционально сопряжены с рианодиновыми рецепторами. В участке сопряжения этих двух типов каналов выход Са2+ приводит к распространению Са'волны возбуждения рианодиновых рецепторов вдоль мембран ретикулума. За фронтом Са2+-волны происходит снижение уровня Са2+ вследствие того, что в ретикулуме в этом участке уже нет кальция, поэтому Са2+ не выходит через каналы, а Са2+-насосы, напротив, выкачивают Са2+ из цитоплазмы. В возбудимых клетках частоту осцилляции Са2+ могут повышать как 1Рз, так и циклическая АДФ-рибоза или кофеин.
Во многих типах клеток волна кальциевой осцилляции распространяется от клеточного ядра и может приобретать форму сфер или сложных спиралей.
В некоторых тканях (сердце, мозг) кальциевые осцилляции, возникшие в одной клетке, могут стимулировать осцилляцию Са2+ в соседних клетках, причём с той же частотой, что и в клетке, инициировавшей этот процесс.
По-видимому, в этих тканях Са2+-волна может распространяться через межклеточные щелевые контакты (см. п. 16.1), обладающие высокой ионной проводимостью.
В цитоплазме одиночной клетки гормоны и факторы роста практически не влияют на амплитуду повышения концентрации Са2+, ъо увеличивают частоту его флуктуаций (рисунок 143). Обычно уровень Са2+ в цитоплазме изменяется от 10 7 до 5-10 7 М, а частота от одного колебания в минуту до нескольких колебаний в секунду.
Са^-зависимые эффекты гормонов и факторов роста оказываются прямо пропорциональными частоте флуктуации цитоплазматического
Са2+ (рисунок 143). Это можно объяснить тем, что при высокой частоте осцилляций увеличивается вероятность насыщения кальцием кальций- связывающих белков.
Диссоциация Са2+ из высокоаффинных участков кальций-связы- вающих белков происходит за минуты, а Са2+ в цитоплазме осциллирует быстрее, поэтому кальций-связывающие белки воспринимают частотную информацию и, подобно преобразователям переменного тока в постоянный, преобразуют её в медленно развивающееся (за минуты или часы) изменение метаболизма, морфологии или функционального состояния клетки.
В кальций-связывающих белках может быть несколько участков связывания Са2+, между которыми проявляется положительная кооперативность. При связывании Са2+ в структуре белка может увеличиваться количество а-спиралей, и часто на поверхность белковой глобулы экспонируются функциональные группы, участвующие во взаимодействии кальций-связывающего белка с так называемыми эффекторными белками. Таким способом, Са2+ индуцирует взаимодействие двух белков, что приводит к изменению их активности или локализации в клетке.
Са2+ активирует многие катаболические процессы (гликолиз, липо- лиз, протеолиз), а также стимулирует синтез белка, мышечное сокращение и немышечную подвижность, экзоцитоз, ионный транспорт, секрецию нейромедиаторов.
14.4.
Еще по теме 14.3. Осцилляции ВНУТРИКЛЕТОЧНОГО КАЛЬЦИЯ:
- Нарушение гомеостаза внутриклеточного кальция
- Нарушение баланса кальция
- Внутриклеточные рецепторы
- Кальций
- Глава 14 Внутриклеточная сигнализация
- Влияние ионов кальция на биосинтез аргинина
- Нарушение обмена кальция и фосфора.
- Глава 8 Внутриклеточный транспорт белков
- Роль ионов кальция в реализации аллергических реакций при бронхиальной астме
- 1.3.4. Значение сАМР как вторичного внутриклеточного мессенджера
- Уровень кальция крови у больных раком предстательной железы.
- 2. Антагонисты кальция: выделены из группы коронарорасширяющих средств