гюйкчвемхе
юЙЮДЕЛХЙ юлм яяяп б. л. фДЮМНБ ГЮМХЛЮКЯЪ ОПНАКЕЛНИ НПЦЮМХВЕЯЙНИ ЩБНКЧЖХХ БЯЧ ЯБНЧ РБНПВЕЯЙСЧ ФХГМЭ. б ОНД╜УНДЕ Й ЩРХЛ БНОПНЯЮЛ НМ ОПХДЕПФХБЮКЯЪ ЬХПНЙХУ АХНКНЦХВЕ╜ЯЙХУ БГЦКЪДНБ МЮ ОПХПНДС ПЮЯЯЛЮРПХБЮЕЛШУ ЪБКЕМХИ.
б. л. фДЮМНБ ЯВХРЮК, ВРН ПЮГБХРХЕ ЛЮРЕПХХ ОПХБНДХР Й БНГ╜МХЙМНБЕМХЧ АХНКНЦХВЕЯЙНИ ФХГМХ Х ПЕГСКЭРЮРНЛ ЕЕ ЯНБЕПЬЕМ╜ЯРБНБЮМХЪ ЛНФЕР ЯРЮРЭ ОНЪБКЕМХЕ ПЮГСЛЮ. щРН ЕЯРЕЯРБЕММШИ ЩРЮО АХНКНЦХВЕЯЙНИ ЩБНКЧЖХХ. бХД, МЮДЕКЕММШИ ПЮГСЛНЛ, ОН╜КСВЮЕР НЦПНЛМШЕ ЯЕКЕЙРХБМШЕ ОПЕХЛСЫЕЯРБЮ Х БНГЛНФМНЯРХ ДКЪ ЯБНЕЦН БШФХБЮМХЪ Х ОПНЖБЕРЮМХЪ. оПХПНДЮ ЯНБЕПЬХКЮ РПХ ОНОШРЙХ ЯНГДЮРЭ ПЮГСЛ. дБЕ ХГ МХУ, ЩРН ЛНГЦ НАЫЕЯРБЕММШУ МЮЯЕЙНЛШУ Х ЛНГЦ НЯЭЛХМНЦЮ, ГЮЙНМВХКХЯЭ МЕСДЮВМН, Ю ОНЯКЕД╜МЪЪ ОПХМЕЯКЮ СЯОЕУ Х ДЮКЮ БНЯУНДЪЫСЧ БЕРБЭ !ОПНЦПЕЯЯХБМНИ ЩБНКЧЖХХ Б БХДЕ НЯНАНЦН БХДЮ ≈ ВЕКНБЕЙЮ ПЮГСЛМНЦН. б. л. фДЮМНБЮ ХМРЕПЕЯНБЮКХ ЙНЯЛНКНЦХВЕЯЙХЕ ОПНАКЕЛШ. нМ АШК ЯРНПНММХЙНЛ ХДЕХ Н ЛМНФЕЯРБЕММНЯРХ НВЮЦНБ ФХГМХ БН бЯЕКЕММНИ. нДХМ ХГ РЮЙХУ НВЮЦНБ, Р. Е. МЮЬЮ ОКЮМЕРЮ гЕЛКЪ Х ЯСЫЕЯРБСЧЫЮЪ МЮ МЕИ АХНЯТЕПЮ, ЯРЮК ОПЕДЛЕРНЛ ЕЦН МЮСВМНЦН РБНПВЕЯРБЮ. хЯЯКЕДСЪ ЩБНКЧЖХНММСЧ ХЯРНПХЧ АХНЯТЕПШ, б. л. фДЮМНБ ЙЮЙ ЯОЕЖХЮКХЯР-БХПСЯНКНЦ ХГСВЮК ЩБНКЧЖХЧ ЖЮПЯРБЮ бХПЮ. яНЕДХМЪЪ ВЮЯРМНЕ Я НАЫХЛ, НМ ОНДУНДХК Й ЩБН╜КЧЖХХ БХПСЯНБ Я НАЫЕАХНКНЦХВЕЯЙХУ ОНГХЖХИ Х ПЮЯЙПШБЮК ГМЮВЕМХЕ ДКЪ ЩБНКЧЖХНММНИ .РЕНПХХ ДЮММШУ, МЮЙНОКЕММШУ БХ╜ПСЯНКНЦХЕИ. рБНПВЕЯЙНЕ МЮЯКЕДХЕ б. л. фДЮМНБЮ ОН ЩБНКЧЖХХ БЙКЧВЮЕР 5 ЛНМНЦПЮТХИ, БШЬЕДЬХУ Б ЯБЕР Б 1953≈1984 ЦЦ. пЮАНРЮ МЮД ОНЯКЕДМЕИ ЙМХЦНИ ╚щБНКЧЖХЪ БХПСЯНБ╩ НАНПБЮКЮЯЭ Б 1987 Ц. щРХ ЙМХЦХ НЙЮГЮКХ АНКЭЬНЕ БКХЪМХЕ МЮ ЯРЮМНБКЕМХЕ МЮСВМШУ БГЦКЪДНБ МЮ ОПНЖЕЯЯ ЩБНКЧЖХХ БХПСЯНБ Х НПЦЮМХВЕ╜ЯЙНИ ЩБНКЧЖХХ Б ЖЕКНЛ. б ДЮММНИ ЯБЪГХ ХМРЕПЕЯМН ПЮЯЯЛНРПЕРЭ ЯНБПЕЛЕММНЕ ЯНЯРНЪМХЕ Х ОСРХ ДЮКЭМЕИЬЕЦН ПЮГБХРХЪ СВЕМХЪ НА ЩБНКЧЖХХ БХПСЯНБ, Ю РЮЙФЕ МНБЕИЬХЕ ДНЯРХФЕМХЪ Б ЩРНИ НАКЮЯРХ. мЮЬЕ ГЮЙКЧВЕМХЕ ОНЯБЪЫЕМН ПЕЬЕМХЧ ЩРНИ ГЮДЮВХ. пЮЯЯЛЮРПХБЮЕРЯЪ ОЕПХНД 1987≈1988 ЦЦ. хГКНФЕМХЕ МЕ ОПЕРЕМ╜ДСЕР МЮ ОНКМНРС МХ ОН ЙПСЦС ГЮРПНМСРШУ ОПНАКЕЛ, МХ ОН ВХЯКС ХЯОНКЭГНБЮММШУ КХРЕПЮРСПМШУ ХЯРНВМХЙНБ. жЕКЭ ПЮАНРШ ГЮ╜ЙКЧВЮКЮЯЭ 'б БШДЕКЕМХХ МЕЯЙНКЭЙХУ ЙКЧВЕБШУ БНОПНЯНБ, ОПЕД╜ЯРЮБКЪЧЫХУЯЪ МЮХАНКЕЕ ЯСЫЕЯРБЕММШЛХ. нОПЕДЕКЕММНЕ ЛЕЯРН ГЮМХЛЮЧР РЮЙФЕ НАЯСФДЕМХЕ ДХЯЙСЯЯХНММШУ ЩБНКЧЖХНММШУ ОПНАКЕЛ Х НОПЕДЕКЕМХЕ ЛЕЯРЮ Б ЩРНИ ДХЯЙСЯЯХХ ОНГХЖХХ б. л. фДЮМНБЮ.
щБНКЧЖХНММШЕ НЯМНБШ ЯХЯРЕЛЮРХЙХ Б Х П С- Я Н Б. рЮЙЯНМНЛХЪ БХПСЯНБ БМЕЯКЮ ОНГХРХБМШИ БЙКЮД Б НАЫСЧ- ЯХЯРЕЛЮРХЙС. б БХПСЯНКНЦХХ ЯНГДЮМЮ РЮЙ МЮГШБЮЕЛЮЪ СМХБЕП╜ЯЮКЭМЮЪ ФКЮЯЯХ-ТХЙЮЖХЪ, Б ЙНРНПНИ БШДЕКЕМХЕ МЮДБХДНБШУ РЮЙЯНМНБ ОПНБНДХРЯЪ МЕ РНКЭЙН ОН ТЕМНРХОХВЕЯЙХЛ ОПХГМЮЙЮЛ, ■МН Х МЮ НЯМНБЕ ТСМДЮЛЕМРЮКЭМШУ ЯБНИЯРБ. сВХРШБЮЧРЯЪ РХО МСЙКЕХМНБНИ ЙХЯКНРШ, НПЦЮМХГЮЖХЪ ЦЕМНЛЮ, ЯОНЯНА ПЕЮКХГЮЖХХ ЦЕМЕРХВЕЯЙНИ ХМТНПЛЮЖХХ, ЯРПНЕМХЕ БХПХНМЮ Х ДПСЦХЕ ЛНКЕ╜ЙСКЪПМН-АХНКНЦХВЕЯЙХЕ, АХНУХЛХВЕЯЙХЕ Х ТСМЙЖХНМЮКЭМШЕ ОН╜ЙЮГЮРЕКХ. оНДНАМШИ ОНДУНД ЛЮКН ХЯОНКЭГСЕРЯЪ Б ЯХЯРЕЛЮРХЙЕ БШЯЬХУ ЩСЙЮПХНРНБ. оПЮЙРХВЕЯЙЮЪ ЙКЮЯЯХТХЙЮЖХЪ ЩРХУ НПЦЮ╜МХГЛНБ ДН ЯХУ ОНП НЯМНБШБЮЕРЯЪ МЮ ЛНПТНКНЦХВЕЯЙХУ ЙПХРЕ╜ПХЪУ. нОЕПЕДХБ ЯХЯРЕЛЮРХЙС БШЯЬХУ ЩСЙЮПХНРНБ Б СЙЮГЮММНЛ НРМНЬЕМХХ, ЯХЯРЕЛЮРХЙЮ БХПСЯНБ НРЯРЮКЮ НР МЕЕ ОН АХНКНЦХВЕ╜ЯЙХЛ ОПХМЖХОЮЛ ОНЯРПНЕМХЪ. б ЯХЯРЕЛЮРХЙЕ БХПСЯНБ ОПХ БШ╜ДЕКЕМХХ РЮЙЯНМНБ БШЯНЙНЦН ПЮМЦЮ МЕ СВРШБЮЧРЯЪ ЩБНКЧЖХНМ╜МШЕ ЯБЪГХ ЛЕФДС МХЛХ. щРН НАСЯКНБКЕМН РЕЛ, ВРН НМХ ДН ЯЮ╜ЛНЦН ОНЯКЕДМЕЦН БПЕЛЕМХ МЕ АШКХ ПЮЯЙПШРШ.
сЯОЕУХ МЮСЙХ Х Б ОЕПБСЧ НВЕПЕДЭ МЮЙНОКЕМХЕ ДЮММШУ Н ОЕПБХВМНИ ЯРПСЙРСПЕ БХПСЯМШУ ЦЕМНБ Х ХУ АЕКЙНБШУ ОПНДСЙРНБ ЯНГДЮКХ ОПЕДОНЯШКЙХ ДКЪ ОНЯРПНЕМХЪ ЩБНКЧЖХНММШУ МЮВЮК ЯХЯРЕЛЮРХЙХ БХПСЯНБ. б. л. фДЮМНБ ОПХМЪК ЮЙРХБМНЕ СВЮЯРХЕ Б ЩРНИ ПЮАНРЕ. б НЯМНБС ЯХЯРЕЛШ, ОПЕДКНФЕММНИ ХЛ ДКЪ PHK- ЦЕМНЛМШУ БХПСЯНБ, ОНКНФЕМЮ ЯРПЮРЕЦХЪ БХПСЯМНЦН ЦЕМНЛЮ, Ю ЯЮЛХ БХПСЯШ ЯЦПСООХПНБЮМШ ОН ЛЕПЕ БНГПЮЯРЮМХЪ ЯКНФМНЯРХ ЯРПНЕМХЪ.
б МЮЯРНЪЫЕЕ БПЕЛЪ Б КХРЕПЮРСПЕ ОНЪБХКХЯЭ ДЮММШЕ, ОНГБН╜КХБЬХЕ ОПНХГБЕЯРХ ДЮКЭМЕИЬХИ ЮМЮКХГ ЩБНКЧЖХНММШУ ЯБЪГЕИ ЛЕФДС ПЪДНЛ пмй-ЦЕМНЛМШУ БХПСЯНБ, ЛЕФДС БХПСЯМШЛХ Х ЙКЕРНВМШЛХ ЦЕМНЛЮЛХ, Ю РЮЙФЕ ПЮЯЯЛНРПЕРЭ ЩБНКЧЖХНММШЕ БГЮХЛННРМНЬЕМХЪ БХПНХДНБ, БХПХНХДНБ Х ЙКЕРЙХ [Kingsbu╜ry D. W., 1988; Goldbach R., 1988]. мЮ НЯМНБЮМХХ ПЕГСКЭРЮРНБ ЯЕЙБЕМХПНБЮМХЪ БХПСЯМШУ пмй, ЯНОНЯРЮБКЕМХЪ ЯРПЮРЕЦХХ ЦЕ╜МНЛЮ, ЯРПСЙРСПШ Х ТСМЙЖХХ АЕКЙНБ R.
Goldbach (1987), R. Gold╜bach Х J. Wellink (1988) ОПХУНДЪР Й БШБНДС, ВРН пмй-ЦЕМНЛ - МШЕ БХПСЯШ ПЮЯРЕМХИ Я ОНКНФХРЕКЭМНИ ОНКЪПМНЯРЭЧ PHK ХЛЕЧР ЯУНДЯРБН Я БХПСЯЮЛХ ФХБНРМШУ Х ПЮГДЕКЪЧРЯЪ МЮ ДБЕ ЯСОЕПЦПСООШ≈ОХЙНПМЮ-ОНДНАМШУ Х яХМДАХЯ-ОНДНАМШУ БХПС╜ЯНБ. й ЯСОЕПЦПСООЕ ОХЙНПМЮ-ОНДНАМШУ БХПСЯНБ ПЮЯРЕМХИ НРМН╜ЯЪРЯЪ ЙНЛН-, МЕОН- Х ОНРХЦПЕЙБХПСЯШ. нАЫМНЯРЭ ЛЕФДС ЩРХЛХ БХПСЯЮЛХ Х ОХЙНПМЮБ'ХПСЯЮЛХ, ОНПЮФЮЧЫХЛХ ФХБНРМШУ, ГЮ╜ЙКЧВЮЕРЯЪ Б ЯКЕДСЧЫЕЛ. 1. бН БЯЕУ ЯКСВЮЪУ Й 5'-ЙНМЖС ЦЕМНЛ╜МНИ PHK ОПХЯНЕДХМЕМ АЕКНЙ VPgЯ МЕАНКЭЬНИ ЛНКЕЙСКЪПМНИ
пХЯ. 63. яПЮБМЕМХЕ ЦЕМНЛНБ ОХЙНПМЮБХПСЯНБ (ОНКХНБХПСЯ ≈ II) Х ЙНЛНБХПС- ЯНБ (БХПСЯ ЛНГЮХЙХ ЙНПНБЭЕЦН ЦНПНУЮ ≈ блйц) [ОН Goldbach R., Wel╜link W., 1988].
оПЪЛНСЦНКЭМХЙХ ≈ НРЙПШРШЕ ВХРЮЧЫХЕ ПЮЛЙХ; ГЮЬРПХУНБЮМН ≈ НАКЮЯРХ БШЯНЙНИ (АН╜КЕЕ 20%) ЦНЛНКНЦХХ; 1 ≈ АЕКНЙ VPgХЮ 5'-ЙНХЖЕ ЦЕМНЛЮ; 2-МСЙКЕНРМДЯБЪГШБЮЧЫЮЪ НА╜КЮЯРЭ; 3 ≈ НАКЮЯРЭ ЖХЯРЕХМОПНРЕЮГШ; 4 ≈ ОНКХЛЕПЮГМЮЪ НАКЮЯРЭ; ю ≈ ОНКХ(ю)-ОНЯКЕДН- БЮРЕКЭМНЯРЭ ХЮ г'-ЙНМЖЕ ЦЕМНЛЮ; VPg ≈ НАКЮЯРЭ ЙЮОЯХДМШУ АЕКЙНБ VPg∖ P ≈ НАКЮЯРЭ ОПНРЕЮГШ; POL≈ НАКЮЯРЭ пмй-ГЮБХЯХЛНИ пмй-ОНКХЛЕПЮГШ.
ЛЮЯЯНИ, Ю МЮ г'-ЙНМЖЕ ЦЕМНЛЮ ХЛЕЕРЯЪ ОНКХ (ю)-СВЮЯРНЙ. 2. оЕП╜БХВМШЛ ОПНДСЙРНЛ РПЮМЯКЪЖХХ БХПСЯМНЦН ЦЕМНЛЮ ЪБКЪЕРЯЪ ЦХ╜ЦЮМРЯЙХИ ОНКХОЕОРХД-ОПЕДЬЕЯРБЕММХЙ, ЙНРНПШИ ОСРЕЛ ОПНРЕН╜КХРХВЕЯЙНЦН ПЮЯЫЕОКЕМХЪ МЮПЕГЮЕРЯЪ МЮ ТСМЙЖХНМЮКЭМШЕ АЕК╜ЙХ. 3. мЕЯРПСЙРСПМШЕ АЕКЙХ ЙНДХПСЧРЯЪ НДХМЮЙНБШЛ МЮАНПНЛ ЦЕМНБ, ЙНРНПШИ ЛНФЕР ПЮЯЯЛЮРПХБЮРЭЯЪ ЙЮЙ ОНЯРНЪММШИ ЦЕМ╜МШИ ЛНДСКЭ. 4. яРЕОЕМЭ ЦНЛНКНЦХХ МЕЯРПСЙРСПМШУ АЕКЙНБ ОПЕ╜БШЬЮЕР 20%. бН БЯЕУ ЯКСВЮЪУ ЩРХ АЕКЙХ БНБКЕЙЮЧРЯЪ Б ПЕОКХ╜ЙЮЖХЧ PHK-
вЕПРШ ЯУНДЯРБЮ Б НПЦЮМХГЮЖХХ ЦЕМНЛЮ ОНКХНБХПСЯЮ (ЛНМН- ОЮПРХРМШИ БХПСЯ) Х ЙНЛНБХПСЯЮ ЛНГЮХЙХ ЙНПНБЭЕЦН ЦНПНУЮ (ПЮГНАЫЕММШИ ДХОЮПРХРМШИ ЦЕМНЛ) ОПЕДЯРЮБКЕМШ МЮ ПХЯ. 63. нАМЮПСФХБЮЕРЯЪ ЯУНДЯРБН Б ОНПЪДЙЕ ПЮЯОНКНФЕМХЪ Х ТСМЙ╜ЖХХ ЦЕМНБ ЙЮОЯХДМШУ АЕКЙНБ, АЕКЙЮ VPg,ОНКХЛЕПЮГМНЦН ЙНЛО╜КЕЙЯЮ Х ОПНРЕЮГ. б ОНКХЛЕПЮГМНИ Х МСЙКЕНРХДЯБЪГШБЮЧЫЕИ НАКЮЯРЪУ ХЛЕЧРЯЪ ЦНЛНКНЦХВМШЕ ОНЯКЕДНБЮРЕКЭМНЯРХ.
й ЯСОЕПЦПСООЕ яХМДАХЯ-ОНДНАМШУ БХПСЯНБ ПЮЯРЕМХИ НРМН╜ЯЪРЯЪ ХКЪПБХПСЯШ, АПНЛНБХПСЯШ, ЙСЙСЛНБХПСЯШ, РНАЮЛНБХПСЯШ, РНАПЮБХПСЯШ, ТСПНБХПСЯШ, УНПДЕИБХПСЯШ, ЙЮПЛНБХПСЯШ Х РНЛ- АСПБХПСЯШ. нАЫХЛ ОПХГМЮЙНЛ ЩРХУ БХПСЯНБ ЪБКЪЧРЯЪ ЯНУПЮ╜МЕМХЕ ЦНЛНКНЦХВМШУ СВЮЯРЙНБ ЦЕМНЛЮ, МЮКХВХЕ ЙЩО-ЯРПСЙРСПШ МЮ 5'-κoH∣∏e PHK Х НАПЮГНБЮМХЕ ЯСАЦЕМНЛМШУ PHK Б ОПНЖЕЯЯЕ БХПСЯМНЦН ПЕОКХЙЮРХБМНЦН ЖХЙКЮ. мЕЯРПСЙРСПМШЕ АЕКЙХ яХМД╜АХЯ-ОНДНАМШУ БХПСЯНБ ХЛЕЧР ЦНЛНКНЦХВМШЕ СВЮЯРЙХ Я МЕЯРПСЙ╜РСПМШЛХ АЕКЙЮЛХ БХПСЯЮ яХМДАХЯ Х ЙНДХПСЧРЯЪ ЦЕМЮЛХ, НПЦЮ╜МХГНБЮММШЛХ ЯУНДМШЛ НАПЮГНЛ. щРХ АЕКЙХ ХЛЕЧР МСЙКЕНРХД-
ЯБЪГШБЮЧЫСЧ НАКЮЯРЭ, НАКЮДЮЧР пмй-ОНКХЛЕПЮГМНИ ЮЙРХБ╜МНЯРЭЧ Х БНБКЕЙЮЧРЯЪ Б ПЕОКХЙЮЖХЧ пмй. рЮЙХЛ НАПЮГНЛ, ВКЕМШ ЩРНИ ЦПСООШ НАМЮПСФХБЮЧР ЯУНДЯРБН Я БХПСЯНЛ яХМД- АХЯ Б ПЮЯОНКНФЕМХХ Х ТСМЙЖХХ ЦЕМНБ ОНКХЛЕПЮГШ ХКХ, ХМЮВЕ, БЯЕ ЩРХ БХПСЯШ ХЛЕЧР НАЫХИ ПЕОКХЙЮРХБМШИ ЛНДСКЭ. щРН- СЙЮ╜ГШБЮЕР МЮ НАЫХЕ ЩБНКЧЖХНММШЕ ЙНПМХ. юМЮКНЦХВМШИ БШБНД ЛНФМН ЯДЕКЮРЭ Б НРМНЬЕМХХ ОХЙНПМЮ-ОНДНАМШУ БХПСЯНБ ПЮЯРЕ╜МХИ Х ОХЙНПМЮБХПСЯНБ ФХБНРМШУ.
пЮЯРХРЕКЭМНЕ Х ФХБНРМНЕ ЖЮПЯРБЮ ПЮГНЬКХЯЭ Б ЯБНЕИ ЩБН╜КЧЖХХ НЙНКН l,2?109ЛКМ КЕР МЮГЮД. сВХРШБЮЪ БШЯНЙСЧ ЯЙН╜ПНЯРЭ ЩБНКЧЖХХ пмй-ЦЕМНЛНБ, БНГМХЙМНБЕМХЕ ОПЕДЙНБ ЯНБПЕ╜ЛЕММШУ пмй-ЦЕМНЛМШУ БХПСЯНБ ПЮЯРЕМХИ Х ОНГБНМНВМШУ ФХ╜БНРМШУ ЯКЕДСЕР НРМЕЯРХ Й АНКЕЕ ОНГДМЕИ ЩОНУЕ. б ДЮКЭМЕИЬЕЛ ЩРХ ОПНБХПСЯШ ПЮГНЬКХЯЭ Б ЯБНЕИ ЩБНКЧЖХХ, ДЮКХ ЛМНЦНВХЯ╜КЕММШЕ БЕРБХ, ГЮМЪКХ ПЮГКХВМШЕ ЩЙНКНЦХВЕЯЙХЕ МХЬХ Х ГЮЙПЕ╜ОХКХЯЭ Б ПЮЯРЕМХЪУ Х ОНГБНМНВМШУ ╚УНГЪЕБЮУ╩.
хГБЕЯРМШЕ Б МЮЯРНЪЫЕЕ БПЕЛЪ ОХЙНПМЮ-ОНДНАМШЕ БХПСЯШ ПЮЯРЕМХИ ЯСЫЕЯРБЕММН ПЮГКХВЮЧРЯЪ ЛЕФДС ЯНАНИ ОН ЛНПТНКН╜ЦХХ БХПХНМЮ Х ПЪДС ДПСЦХУ ОПХГМЮЙНБ. бХПХНМШ ЙНЛН- Х МЕОН- БХПСЯНБ ЯТЕПХВЕЯЙХЕ, Ю БХПХНМШ ОНРХЦПЕЙБХПСЯНБ ОЮКНВЙННА╜ПЮГМШЕ. цЕМНЛ ЙНЛН- Х МЕОНБХПСЯНБ ПЮГНАЫЕМ Б ДБСУ ВЮЯРХЖЮУ, Ю ОАРХЦПЕЙБХПСЯШ ЪБКЪЧРЯЪ ЛНМНОЮПРХРМШЛХ. еЫЕ АНКЭЬХЛ ПЮГМННАПЮГХЕЛ УЮПЮЙРЕПХГСЧРЯЪ яХМДАХЯ-ОНДНАМШЕ БХПСЯШ. дКЪ АНКЭЬХМЯРБЮ ХГ МХУ УЮПЮЙРЕПМШ ЛСКЭРХОЮПРХРМНЯРЭ Х ПЮГКХВХЪ Б РПЮМЯКЪЖХНММНИ ЯРПЮРЕЦХХ ЦЕМНЛЮ. бЯЕ ЩРХ ОПХГМЮ╜ЙХ НРМНЯЪРЯЪ Й БРНПХВМШЛ, ОПХНАПЕРЕММШЛ Б УНДЕ ЩБНКЧЖХХ.
бНГМХЙМНБЕМХЕ ЛСКЭРХОЮПРХРМШУ ЦЕМНЛНБ пмй-ЯНДЕПФЮЫХУ БХПСЯНБ ЯБЪГШБЮЕРЯЪ, МЮОПХЛЕП, Я ЯНВЕРЮММШЛ ДЕИЯРБХЕЛ ЩЙН╜КНЦХВЕЯЙХУ ТЮЙРНПНБ Х БКХЪМХЕЛ ОПХПНДШ ЦЕМЕРХВЕЯЙНЦН ЛЮ╜РЕПХЮКЮ [Nee S., 1987]. бЕДСЫСЧ ПНКЭ ДКЪ ЩБНКЧЖХХ Б ЩРНЛ МЮОПЮБКЕМХХ ХЦПЮЧР БШЯНЙЮЪ ЛМНФЕЯРБЕММНЯРЭ ОЕПЕМНЯЮ БХ╜ПСЯНБ ПЮЯРЕМХИ, НРЯСРЯРБХЕ ОНКМНЖЕММНИ ХЛЛСММНИ ЯХЯРЕЛШ ╚УНГЪХМЮ╩, ЯОНЯНАМНИ НЦПЮМХВХБЮРЭ ПЮГЛМНФЕМХЕ БХПСЯЮ, Х ГМЮВХРЕКЭМЮЪ НЬХАЙЮ ПЕОКХЙЮЖХХ пмй, НОПЕДЕКЪЧЫЮЪ ЯЕКЕЙ╜РХБМШЕ ОПЕХЛСЫЕЯРБЮ ПЮГНАЫЕММШУ ЦЕМНЛНБ.бЮФМШЛ ЩЙНКНЦХВЕЯЙХЛ ТЮЙРНПНЛ ЩБНКЧЖХНММНЦН ПЮГБХРХЪ ОНКНФХРЕКЭМШУ пмй-ЦЕМНЛМШУ БХПСЯНБ АШКН ХЯОНКЭГНБЮМХЕ ВКЕМХЯРНМНЦХУ ОЕПЕМНЯВХЙНБ, НАЕЯОЕВХБЮЧЫХУ ЙНМРЮЙР ЛЕФДС ПЮЯРЕМХЪЛХ Х НПЦЮМХГЛНЛ ОНГБНМНВМШУ ╚УНГЪЕБ╩. хМРЕПЕЯМН НРЛЕРХРЭ, ВРН НДМХЛ ХГ ЛЕУЮМХГЛНБ ЯНГДЮМХЪ ХГЛЕМВХБНЯРХ ЯКСФХКХ ПЕЙНЛАХМЮЖХХ. пНКЭ ОНЯКЕДМХУ Б ЩБНКЧЖХХ ОНКНФХ╜РЕКЭМШУ пмй-ЦЕМНЛНБ НЯРЮБЮКЮЯЭ МЕДНЯРЮРНВМН ЪЯМНИ. пЕ╜ГСКЭРЮРШ ОПНХЯЬЕДЬХУ ПЕЙНЛАХМЮЖХНММШУ ЯНАШРХИ ЛНФМН БХДЕРЭ ОПХ ЯПЮБМЕМХХ ЦЕМНЛНБ МЕЙНРНПШУ ОХЙНПМЮ- Х яХМДАХЯ- ОНДНАМШУ БХПСЯНБ ПЮЯРЕМХИ [Goldbach R., Wellink J., 1988]. б ЖЕКНЛ ХРНЦНЛ ЩБНКЧЖХХ ЪБКЪЕРЯЪ НАПЮГНБЮМХЕ ДБСУ БЕРБЕИ ОНКНФХРЕКЭМШУ пмй-ЦЕМНЛМШУ БХПСЯНБ, НАЗЕДХМЕММШУ НАЫ╜
МНЯРЭЧ ОПНХЯУНФДЕМХЪ, ЯНУПЮМХБЬХУ ЩКЕЛЕМРШ ЯРПСЙРСПМН╜ТСМЙЖХНМЮКЭМНЦН ЕДХМЯРБЮ Х ХЯОНКЭГСЧЫХУ Б ЙЮВЕЯРБЕ ╚УНГЪ╜ЕБ╩ ОПЕДЯРЮБХРЕКЕИ ФХБНРМШУ Х ПЮЯРХРЕКЭМШУ ЖЮПЯРБ.
б. л. фДЮМНБ ПЮГДЕКЪК ЛМЕМХЕ НА ЩБНКЧЖХНММШУ ЯБЪГЪУ БХПСЯНБ Я ЦЕМЕРХВЕЯЙХЛХ ЩКЕЛЕМРЮЛХ ЙКЕРЙХ. щРЮ ХДЕЪ ОНКС╜ВЮЕР БЯЕ АНКЭЬЕЕ ПЮГБХРХЕ. мЮ НЯМНБЮМХХ ЯРПСЙРСПМН-ТСМЙ╜ЖХНМЮКЭМНЦН ЯУНДЯРБЮ Х ЦНЛНКНЦХХ ЦЕМНЛЮ ОНЯРПНЕМШ ДБЕ ТХ╜КНЦЕМЕРХВЕЯЙХЕ КХМХХ: 1) ХМРПНМШ ЦПСООШ I ≈ БХПНХДШ≈ БХ- ПСЯНХДШ ≈ ДЕКЭРЮ-ЮЦЕМР Х 2) РПЮМЯОНГНМШ ≈ ПЕРПНБХПСЯШ ≈ ЦЕОЮДМЮБХПСЯШ [Kingsbury D. W., 1988]. нАЫМНЯРЭ ОЕПБНИ ТХКНЦЕМЕРХВЕЯЙНИ КХМХХ НОПЕДЕКЪЕРЯЪ РЕЛ, ВРН ЦЕМНЛ ЯНЯРЮБ╜КЪЧЫХУ ЕЕ ЮЦЕМРНБ БН БЯЕУ ЯКСВЮЪУ ОПЕДЯРЮБКЕМ МЕАНКЭЬНИ ОН ПЮГЛЕПС НДМНМХРЕБНИ пмй, АНЦЮРНИ ЙНЛОКЕЛЕМРЮПМШЛХ СВЮЯРЙЮЛХ. хЛЕЧРЯЪ НАКЮЯРХ БШЯНЙНИ ЦНЛНКНЦХХ. оПЮПНДХ╜РЕКЭЯЙНИ ТНПЛНИ, ОН-БХДХЛНЛС, ЪБКЪЧРЯЪ ОНДБХФМШЕ ЦЕМЕРХ╜ВЕЯЙХЕ ЩКЕЛЕМРШ ЙКЕРЙХ, ОНДНАМШЕ ХМРПНМЮЛ ЦПСООШ I, ДЮБЬХЕ МЮВЮКН БХПНХДЮЛ.
хГБЕЯРМН 14 БХПНХДНБ. бЯЕ НМХ ХЯОНКЭГСЧР Б ЙЮВЕЯРБЕ ╚УН╜ГЪЕБ╩ ПЮЯРЕМХЪ. оПЕДЯРЮБКЪЧР ЯНАНИ ╚ЦНКСЧ╩ пмй, МЕ ХЛЕЧ╜ЫСЧ АЕКЙНБНИ НАНКНВЙХ. оПНРЪФЕММНЯРЭ ЦЕМНЛЮ 246≈371 МСЙ╜КЕНРХДНБ. мЕ РПЕАСЧР ДКЪ ЯБНЕИ ПЕОКХЙЮЖХХ БХПСЯЮ-ОНЛНЫМХ╜ЙЮ. пЕОКХЙЮЖХЪ БХПНХДНБ СОПЮБКЪЕРЯЪ дмй-ГЮБХЯХЛНИ PHK- ОНКХЛЕПЮГНИ ЙКЕРЙХ ╚УНГЪХМЮ╩ Я ХЯОНКЭГНБЮМХЕЛ Йпмй-НКХЦН- ЛЕПНБ Б ЙЮВЕЯРБЕ ЛЮРПХЖШ. кЕРЮКЭМНЕ ДЕИЯРБХЕ БХПНХДНБ МЮ ПЮЯРХРЕКЭМСЧ ЙКЕРЙС ЯБЪГШБЮЕРЯЪ Я ОНДЮБКЕМХЕЛ ОПЕ-Лпмй- ОПНЖЕЯЯХМЦЮ [Smarda S., 1987; Albanese J., La Rosa R., 1988].
бХПСЯНХДШ ПЮЯЯЛЮРПХБЮЧРЯЪ ЙЮЙ БХПНХДШ, СРПЮРХБЬХЕ ЯОН╜ЯНАМНЯРЭ Й ЯЮЛНЯРНЪРЕКЭМНИ ПЕОКХЙЮЖХХ. пЕОКХЙЮЖХЪ БХПСЯНХ- ДНБ НЯСЫЕЯРБКЪЕРЯЪ ОПХ ОНЛНЫХ БХПСЯЮ-ОНЛНЫМХЙЮ. хЯОНКЭ╜ГСЧР Б ЙЮВЕЯРБЕ ╚УНГЪЕБ╩ ПЮЯРЕМХЪ. цЕМНЛ ХЛЕЕР ОПНРЪФЕМ╜МНЯРЭ 350≈400 НЯМНБЮМХИ Х ЯНДЕПФХР ОПНРЕХМЙНДХПСЧЫХЕ ОН╜ЯКЕДНБЮРЕКЭМНЯРХ, МЮОПЮБКЪЧЫХЕ ЯХМРЕГ АЕКЙНБ, ЙНРНПШЕ СВЮ╜ЯРБСЧР Б НАПЮГНБЮМХХ НАНКНВЙХ БХПСЯНХДНБ.
нЯНАНЕ ЛЕЯРН Б ПЮЯЯЛЮРПХБЮЕЛНИ ТХКНЦЕМЕРХВЕЯЙНИ КХМХИ ГЮМХЛЮЕР ДЕКЭРЮ-ЮЦЕЖР, ОПЕДЯРЮБКЪЧЫХИ ЯНАНИ ЕДХМЯРБЕММШИ ХГБЕЯРМШИ Б МЮЯРНЪЫЕЕ БПЕЛЪ БХПСЯНХД-ОНДНАМШИ ОЮРНЦЕМ ВЕ╜КНБЕЙЮ. еЦН ПЕОКХЙЮЖХЪ ОПНХЯУНДХР ОПХ ОНЛНЫХ БХПСЯЮ ЦЕОЮ╜РХРЮ б. цЕМНЛ ДЕКЭРЮ-ЮЦЕМРЮ ЯНЯРНХР ХГ 1683 МСЙКЕНРХДНБ. хЛЕ╜ЧРЯЪ 5 НРЙПШРШУ ПЮЛНЙ ЯВХРШБЮМХЪ, ДБЕ ХГ ЙНРНПШУ МЮУНДЪР╜ЯЪ Б ЦЕМНЛМНИ, Ю РПХ ≈ Б ЮМРХЦЕМНЛМНИ НПХЕМРЮЖХХ [MakinoSh. et al., 1987]. йНДХПСЕЛШИ БХПСЯМШЛ ЦЕМНЛНЛ АЕКНЙ Я ЛНКЕЙС╜КЪПМНИ ЛЮЯЯНИ 24 000 ТНЯТНПХКХПНБЮМ ОН ЯЕПХМС. щРНР ТНЯ╜ТНОПНРЕХД НАКЮДЮЕР пмй-ЯБЪГШБЮЧЫЕИ ЮЙРХБМНЯРЭЧ Х СВЮЯР╜БСЕР Б НАПЮГНБЮМХХ ВЮЯРХЖ ДЕКЭРЮ-ЮЦЕМРЮ [Chang Ming-Fu et al., 1988]. дЕКЭРЮ-ЮЦЕМР МЕ ХЛЕЕР ЮМЮКНЦНБ ЯПЕДХ БХПСЯНБ ФХ╜БНРМШУ. мЕ ЪЯМН, ОПНХЯУНДХР КХ ДЕКЭРЮ-ЮЦЕМР МЕОНЯПЕДЯРБЕММН НР ХМРПНМНБ ЦПСООШ I ХКХ ФЕ ЕЦН ЩБНКЧЖХЪ ЬКЮ ОН ЩРЮОЮЛ ВЕ╜
ПЕГ БХПНХДШ Х БХПСЯНХДШ. C ,ОНЯКЕДМХЛХ ДЕКЭРЮ-ЮЦЕМР ХЛЕЕР ГМЮВХРЕКЭМСЧ ЦНЛНКНЦХЧ [Makino Sh. et al., 1987].
бНГЛНФМНЯРЭ ОНЯРПНЕМХЪ ТХКНЦЕМЕРХВЕЯЙНИ КХМХХ РПЮМЯОН╜ГНМШ≈ ПЕРПНБХПСЯШ ≈ ЦЕОЮДМЮБХПСЯШ НАСЯКНБКЕМЮ РЕЛ, ВРН БУНДЪЫХЕ Б МЕЕ ПЕРПН- Х ЦЕОЮДМЮБХПСЯШ ХЯОНКЭГСЧР ДКЪ ЯБНЕИ ПЕОКХЙЮЖХХ ЛЕУЮМХГЛ НАПЮРМНИ РПЮМЯЙПХОЖХХ. нАКЮЯРЭ ЦЕМН╜ЛЮ, ЙНДХПСЧЫЮЪ НАПЮРМСЧ РПЮМЯЙПХОРЮГС, ХЛЕЕР СВЮЯРЙХ БШЯН╜ЙНИ ЦНЛНКНЦХХ. цЕМНЛМЮЪ дмй ЦЕОЮДМЮБХПСЯНБ Х дмй-ЙНОХЪ ПЕРПНБХПСЯМНЦН ЦЕМНЛЮ НАКЮДЮЧР ЯОНЯНАМНЯРЭЧ БЯРПЮХБЮРЭЯЪ Б ЙКЕРНВМШИ ЦЕМНЛ, НАПЮГСЪ ОПХ ЩРНЛ РПЮМЯОНГНМ-ОНДНАМШЕ ЯРПСЙРСПШ. яРПСЙРСПМН-ТСМЙЖХНМЮКЭМНЕ ЯУНДЯРБН РПЮМЯОНГНМНБ Я ХМРЕЦПХПНБЮММШЛХ ЦЕМНЛЮЛХ ПЕРПН- Х ЦЕОЮДМЮБХПСЯНБ СЙЮГШ╜БЮЕР МЮ ХУ ЩБНКЧЖХНММСЧ НАЫМНЯРЭ. нДМЮЙН ЯКНФМНЯРЭ НПЦЮ╜МХГЮЖХХ МЕ ОНГБНКЪЕР ЯЙЮГЮРЭ, ЙЮЙНИ ЩКЕЛЕМР Б ЩРНИ ЯХЯРЕЛЕ ЪБКЪЕРЯЪ ОЕПБХВМШЛ. мЕ ХЯЙКЧВЕМЮ РЮЙФЕ БНГЛНФМНЯРЭ ЩБН╜КЧЖХНММНЦН ДБХФЕМХЪ Б ДБСУ МЮОПЮБКЕМХЪУ, Р. Е. НР ЙКЕРЙХ Й РПЮМЯЛХЯЯХБМШЛ ЦЕМЕРХВЕЯЙХЛ ЩКЕЛЕМРЮЛ Х НР МХУ Й ЙКЕРЙЕ. пЕРПН- Х ЦЕОЮДМЮБХПСЯШ ОПХ ХМРЕЦПЮЖХХ Б ЦЕМНЛ ЙКЕРЙХ ЯОН╜ЯНАМШ БШГШБЮРЭ ХМЯЕПЖХНММШЕ ЛСРЮЖХХ. рЮЙХЕ ОПНЖЕЯЯШ, БЕ╜ПНЪРМН, ЪБКЪЧРЯЪ БЮФМШЛ ЛЕУЮМХГЛНЛ ТНПЛХПНБЮМХЪ ЩСЙЮ╜ПХНРХВЕЯЙНЦН ЦЕМНЛЮ Б ОПНЖЕЯЯЕ ЩБНКЧЖХХ [Jolais F., Sabo ю., 1988].
яЙНПНЯРЭ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ дмй-ЦЕ╜МНЛМШУ БХПСЯНБ. PHK- Х дмй-ЦЕМНЛМШЕ БХПСЯШ ЯСЫЕ╜ЯРБЕММН ПЮГКХВЮЧРЯЪ ОН СПНБМЧ ЛСРЮАЕКЭМНЯРХ. пЕОКХЙЮЖХЪ PHK ОПНХЯУНДХР Я МХГЙНИ РНВМНЯРЭЧ. б ОЕПЕДЮЧЫСЧЯЪ ХМТНП╜ЛЮЖХЧ БМНЯХРЯЪ ГМЮВХРЕКЭМЮЪ НЬХАЙЮ. вЮЯРНРЮ ЛСРЮЖХИ ГЮ ЦЕ╜МЕПЮЖХЧ ДНЯРХЦЮЕР IO-3≈IO-4. щРН Б 100 000 БШЬЕ, ВЕЛ ВЮЯРНРЮ ЛСРЮЖХИ дмй. гЮ 33 ОНЙНКЕМХЪ пмй-ЦЕМНЛМШЕ БХПСЯШ ЛНЦСР МЮЙНОХРЭ РЮЙНИ ГЮОЮЯ ХГЛЕМВХБНЯРХ, МЮ ЯНГДЮМХЕ ЙНРНПНЦН С дмй-ЦЕМНЛМШУ БХПСЯНБ ОНРПЕЁАНБЮКНЯЭ АШ 10 ЛКМ ОНЙНКЕМХИ. мХГЙЮЪ ЛСРЮАЕКЭМНЯРЭ дмй НОПЕДЕКЪЕРЯЪ ОПХПНДНИ ЩРНЦН ЦЕ╜МЕРХВЕЯЙНЦН ЛЮРЕПХЮКЮ, ЮОНЯН©АНЛ ПЕОКХЙЮЖХХ Х ЯОНЯНАМНЯРЭЧ Й ПЕОЮПЮЖХХ. оПХБЕДЕММШЕ ДЮММШЕ ОНГБНКЪКХ ЯВХРЮРЭ, ВРН дмй-ЦЕМНЛМШЕ БХПСЯШ. УЮПЮЙРЕПХГСЧРЯЪ ЦКСАНЙХЛ ЙНМЯЕПБЮ╜РХГЛНЛ МЮЯКЕДЯРБЕММНЯРХ. б МЮЯРНЪЫЕЕ БПЕЛЪ ЩРН РПЮДХЖХНМ╜МНЕ ЛМЕМХЕ РПЕАСЕР ЕЯКХ МЕ ОЕПЕЯЛНРПЮ, РН СРНВМЕМХЪ.
бНОПНЯ Н ЛСРЮАЕКЭМНЯРХ дмй-ЦЕМНЛМШУ БХПСЯНБ ЪБКЪЕРЯЪ ВЮЯРЭЧ АНКЕЕ НАЫЕЦН БНОПНЯЮ Н ЯЙНПНЯРХ ЛНКЕЙСКЪПМНИ ЩБН╜КЧЖХХ дмй МЮ ПЮГКХВМШУ ЯРСОЕМЪУ ЩБНКЧЖХНММНИ КЕЯРМХЖШ. дХЯЙСЯЯХЪ ОН ЩРНЛС ОНБНДС Б МЮСВММНИ КХРЕПЮРСПЕ БЕДЕРЯЪ АН╜КЕЕ 15 КЕР. мЮ НЯМНБЮМХХ МЮЙНОКЕММШУ ДЮММШУ ЛНФМН ЯВХРЮРЭ СЯРЮМНБКЕММШЛ, ВРН ЯЙНПНЯРЭ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ МЕНДХ╜МЮЙНБЮ Б ПЮГКХВМШУ РЮЙЯНМНЛХВЕЯЙХУ ЦПСООЮУ Х ЕДХМШУ ЛНКЕ╜ЙСКЪПМШУ ВЮЯНБ ЩБНКЧЖХХ МЕ ЯСЫЕЯРБСЕР. нЯМНБШБЮЪЯЭ МЮ ПЕ╜ГСКЭРЮРЮУ ЦХАПХДХГЮЖХХ СМХЙЮКЭМШУ ОНЯКЕДНБЮРЕКЭМНЯРЕИ дмй Х ДЮММШУ ЯЕЙБЕМХПНБЮМХЪ ЦНЛНКНЦХВМШУ ЦЕМНБ Х ОЯЕБДН-
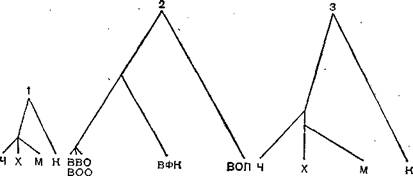
пХЯ. 64. тХКНЦЕМЕРХВЕЯЙХЕ ДПЕБЮ ОНГБНМНВМШУ Х ОНЙЯБХПСЯНБ [ОН Gent╜ry G. A. et al., 1988].
1 ≈ ОНГБНМНВМШЕ; 2 ≈ ОНЙЯБХПСЯШ; 3 ≈ ОНГБНМНВМШЕ. яЙНПНЯРЭ ЩБНКЧЖХХ СПЮБМЕМЮ ЯН ЯЙНПНЯРЭЧ ЩБНКЧЖХХ ОНЙЯБХПСЯНБ (ЙНЩТТХЖХЕМР 2,8). дКХМЮ БЕРБЕИ Б ЯКСВЮЪУ 1 Х 2 ОПНОНПЖХНМЮКЭМЮ ЩБНКЧЖХНММНЛС ПЮЯЯРНЪМХЧ, БШПЮФЕММНЛС Б ВХЯКЕ ЛСРЮЖХИ МЮ 100 ЮЛХМНЙХЯКНРМШУ НЯРЮРЙНБ Б ЛНКЕЙСКЕ РМЛХДММЙХМЮГШ ОНГБНМНВМШУ ХКХ БХПСЯНБ. в ≈ВЕКНБЕЙ; X ≈ УНЛЪЙ; M ≈ ЛШЬЭ; й ≈ ЙСПХЖЮ; BBO ≈ БХПСЯ БЕРПЪМНИ НЯОШ; BOO- БХПСЯ НЯОШ НАЕГЭЪМШ; BOB ≈ БХПСЯ НЯОНБЮЙЖХМШ; бтй ≈ БХПСЯ ТХАПНЛШ ЙПНКХЙЮ; BOn ≈ БХПСЯ НЯОШ ОРХЖ.
ЦЕМНБ С ПЮГМШУ БХДНБ, R. J. Britten (1986) ОПХЬЕК Й БШБНДС, ВРН ЯЙНПНЯРЭ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ ЛНФЕР ПЮГКХВЮРЭЯЪ Б 5 ПЮГ Х ЯНЯРЮБКЪРЭ НР l,3?10~9ДН 6,6?10-9ГЮЛЕМ МЮ СВЮЯРНЙ Б ЦНД. мЮХАНКЕЕ МХГЙХЕ ЯЙНПНЯРХ ЩБНКЧЖХХ дмй НАМЮПСФЕМШ С БШЯЬХУ ОПХЛЮРНБ Х ОРХЖ, Ю МЮХАНКЕЕ БШЯНЙХЕ ≈ С ЦПШГСМНБ, ЛНПЯЙХУ ЕФЕИ Х ДПНГНТХКШ. Li Wen-Hsiung Х. ЯНЮБР. (1987) ЯВХРЮЧР, ВРН ЯЙНПНЯРЭ МСЙКЕНРХДМШУ ГЮЛЕЫЕМХИ С ЦПШГСМНБ Б 4≈8 ПЮГ БШЬЕ, ВЕЛ С БШЯЬХУ ОПХЛЮРНБ, Х Б 2≈4 ПЮГЮ БШЬЕ, ВЕЛ С ЙНОШРМШУ. рЕЛО ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ ЦНЛХМНХДНБ МХФЕ, ВЕЛ БШЯЬХУ ОПХЛЮРНБ. с ВЕКНБЕЙЮ ОН ЯПЮБМЕМХЧ Я ЬХЛ╜ОЮМГЕ Х НПЮМЦСРЮМЦНЛ НРЛЕВЮЕРЯЪ АНКЕЕ МХГЙЮЪ ВЮЯРНРЮ МСЙ╜КЕНРХДМШУ ГЮЛЕЫЕМХИ. б ЙЮВЕЯРБЕ ОПХВХМ ПЮГКХВХИ Б ЯЙНПНЯРХ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ МЮГШБЮЧР ОПНДНКФХРЕКЭМНЯРЭ ОНЙН╜КЕМХИ Х ВХЯКН ПЕОКХЙЮЖХИ дмй Б ЙКЕРЙЮУ ГЮПНДШЬЕБНЦН ОСРХ. сЙЮГШБЮЕРЯЪ РЮЙФЕ МЮ НЯНАЕММНЯРХ ЛЕУЮМХГЛНБ ПЕОКХЙЮЖХХ дмй Х ЩТТЕЙРХБМНЯРЭ ПЕОЮПЮЖХХ.
дмй-РЕМНЛМШЕ БХПСЯШ МЕ ЯНЯРЮБКЪЧР ХЯЙКЧВЕМХЪ Б НАЫЕЛ ОНПЪДЙЕ. яЙНПНЯРЭ ХУ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ ЙЮЙ ЮЦЕМРНБ Я АНКЕЕ ЙНПНРЙХЛ ФХГМЕММШЛ ЖХЙКНЛ ОПЕБШЬЮЕР ЯЙНПНЯРЭ ЛН╜КЕЙСКЪПМНИ ЩБНКЧЖХХ ОНГБНМНВМШУ ╚УНГЪЕБ╩. нАГНП ДЮММШУ ОН ЩРНЛС БНОПНЯС ЯДЕКЮКХ G. A. Gentry Х ЯНЮБР. (1988).
оПЕДЯРЮБКЕМХЕ Н ПЮГКХВХЪУ Б ЯЙНПНЯРХ ЛНКЕЙСКЪПМНИ ЩБН╜КЧЖХХ ОНЙЯБХПСЯНБ Х ХУ ╚УНГЪЕБ╩ ДЮЕР ЯКЕДСЧЫХИ МЮЦКЪДМШИ ОПХЛЕП. мЮ ПХЯ. 64 ОНЙЮГЮМШ ТХКНЦЕМЕРХВЕЯЙХЕ ДПЕБЮ ОНЙЯБХ╜ПСЯНБ Х ОНГБНМНВМШУ, МЮВЕПВЕММШЕ МЮ НЯМНБЮМХХ ПЕГСКЭРЮРНБ ЯПЮБМЕМХЪ ЮЛХМНЙХЯКНРМНИ ОНЯКЕДНБЮРЕКЭМНЯРХ ДЕГНЙЯХРХЛХ-
ДХМЙХМЮГШ. б ЯКСВЮЪУ 1 Х 2 ТХКНЦЕМЕРХВЕЯЙХЕ ДПЕБЮ ОНЯРПНЕ╜МШ ОН ЕДХМНЛС ОПХМЖХОС, МН Б ПЮГКХВМНЛ ЛЮЯЬРЮАЕ. дКХМЮ БЕРБЕИ ОПНОНПЖХНМЮКЭМЮ ЩБНКЧЖХНММНЛС ПЮЯЯРНЪМХЧ, БШПЮ╜ФЕММНЛС Б ВХЯКЕ ЛСРЮЖХИ МЮ 100 ЮЛХМНЙХЯКНРМШУ НЯРЮРЙНБ. лСРЮАЕКЭМНЯРЭ ОНЙЯБХПСЯНБ ОПЕБШЬЮЕР ЯННРБЕРЯРБСЧЫХИ ОН╜ЙЮГЮРЕКЭ ДКЪ ОНГБНМНВМШУ Б 2,8 ПЮГЮ. яПЮБМЕМХЕ ДПЕБЮ 1 Х ДПЕБЮ 2 ОНЙЮГШБЮЕР, ВРН НМХ ПЮГКХВЮЧРЯЪ ОН БШЯНРЕ. щРН ЯБХ╜ДЕРЕКЭЯРБСЕР, ВРН ЯЙНПНЯРЭ ЩБНКЧЖХХ ОНГБНМНВМШУ ЛЕМЭЬЕ ЯЙН╜ПНЯРХ ЩБНКЧЖХХ ОНЙЯБХПСЯНБ. дПЕБН ОНГБНМНВМШУ 3 ОНЯРПНЕМН Б ЛЮЯЬРЮАЕ ТХКНЦЕМЕРХВЕЯЙНЦН ДПЕБЮ ОНЙЯБХПСЯНБ. хЯОНКЭГН╜БЮМ ДНАЮБНВМШИ ЙНЩТТХЖХЕМР 2,8. пЮГКХВХЪ Б ОПНРЪФЕММНЯРХ ДПЕБЮ ОНГБНМНВМШУ Х ДПЕБЮ ОНЙЯБХПСЯНБ, НАСЯКНБКЕММШЕ МЕНДХ╜МЮЙНБНИ ЯЙНПНЯРЭЧ ГЮЛЕЫЕМХЪ ЮЛХМНЙХЯКНРМШУ НЯРЮРЙНБ ДЕГ- НЙЯХРХЛХДХМЙХМЮГШ, ЯРХПЮЧРЯЪ.
бХПСЯШ БЕРПЪМНИ НЯОШ ≈ НОНЪЯШБЮЧЫЕЦН КХЬЮЪ Х БХПСЯШ ЦЕПОЕЯЮ РХОЮ 1Х 2ПЮГНЬКХЯЭ Б ЯБНЕИ ЩБНКЧЖХХ НЙНКН 48,6 ЛКМ КЕР МЮГЮД. яЙНПНЯРЭ ЮЛХМНЙХЯКНРМШУ ГЮЛЕМ Б ЛЮКНИ ЯСАЗЕДХМХЖЕ ПХАНМСЙКЕНРХДПЕДСЙРЮГШ ГЮ ЩРНР ОЕПХНД ОПЕБШ╜ЬЮКЮ ЯННРБЕРЯРБСЧЫХИ ОНЙЮГЮРЕКЭ ДКЪ ТЕПЛЕМРЮ ╚УНГЪЕБ╩ Б 38 ПЮГ. пЮЯЯЛНРПЕММШЕ ДЮММШЕ ОНЙЮГШБЮЧР, ВРН дмд-ЦЕМНЛ- МШЕ БХПСЯШ ЯСЫЕЯРБЕММН НРЯРЮЧР ОН СПНБМЧ ЛСРЮАЕКЭМНЯРХ НР пмй-ЦЕМНЛМШУ, МН МЕЯЙНКЭЙН НОЕПЕФЮЧР ЯБНХУ ОНГБНМНВМШУ ╚УНГЪЕБ╩. щРН ОНГБНКЪЕР ЦНБНПХРЭ НА НРМНЯХРЕКЭМН МХГЙНЛ ЙНМ╜ЯЕПБЮРХГЛЕ МЮЯКЕДЯРБЕММНЯРХ дмй-ЦЕМНЛМШУ БХПСЯНБ х БШДЕ╜КЪРЭ' ХУ ЯПЕДХ НПЦЮМХГЛНБ, ХЯОНКЭГСЧЫХУ Б ЙЮВЕЯРБЕ ЦЕМЕРХ╜ВЕЯЙНЦН ЛЮРЕПХЮКЮ дмй.
лСРЮАЕКЭМНЯРЭ БХПСЯМНЦН ЦЕМНЛЮ ЯНГДЮЕР ОПЕДОНЯШКЙХ ДКЪ НЯСЫЕЯРБКЕМХЪ ЛХЙПНЩБНКЧЖХХ дмй-ЦЕМНЛМШУ БХПСЯНБ, ЙНРН╜ПЮЪ, ЯСДЪ ОН ЯНЯРНЪМХЧ ЕЯРЕЯРБЕММШУ БХПСЯМШУ ОНОСКЪЖХИ, ОПНРЕЙЮЕР ДНЯРЮРНВМН ХМРЕМЯХБМН. нА ЩРНЛ ЯБХДЕРЕКЭЯРБСЧР МЮЯШЫЕММНЯРЭ БХПСЯМШУ ОНОСКЪЖХИ ЛСРЮЖХЪЛХ Х МЮКХВХЕ БМСР- ПХОНОСКЪЖХНММШУ ПЮГКХВХИ. нЯНАЕММН ХМРЕПЕЯМШЕ ДЮММШЕ ОН╜КСВЕМШ ОПХ ХГСВЕМХХ ЮДЕМНБХПСЯНБ. б ЩРХУ ХЯЯКЕДНБЮМХЪУ ОПХЛЕМЪКЮЯЭ ЕДХМЮЪ ЛЕРНДХЙЮ ПЕЯРПХЙЖХНММНЦН ЮМЮКХГЮ БХПХ-, НММНИ дмй. оНЩРНЛС ПЕГСКЭРЮРШ ПЮГКХВМШУ ЮБРНПНБ ЯНОНЯРЮ╜БХЛШ. Li Quan-Gen Х G. Wadell (1988) ХЯОНКЭГНБЮКХ ДКЪ ЮМЮ╜КХГЮ 61 ЬРЮЛЛ ЮДЕМНБХПСЯЮ РХОЮ 3,ХГНКХПНБЮММШИ МЮ 6 ЙНМ╜РХМЕМРЮУ, ЩМДНМСЙКЕЮГШ Bell, Bgll 1, Hpal, SallХ Smal. бШДЕКЕМШ 17 ЦЕМНЛМШУ РХОНБ, ЙНРНПШЕ ПЮГДЕКЕМШ МЮ 3 ЦЕМНЛ╜МШЕ ЦПСООШ. ьРЮЛЛШ, НРМЕЯЕММШЕ Й 1-И ЦПСООЕ, АШКХ БШДЕКЕМШ Б юТПХЙЕ, еБПНОЕ, яЕБЕПМНИ Х чФМНИ юЛЕПХЙЕ, ЙН 2-И ≈ Б юТПХЙЕ, Й 3-И НРМЕЯЕМШ ЬРЮЛЛШ, ЙНРНПШЕ ХЛЕКХ ЦКНАЮКЭМНЕ ПЮЯОПНЯРПЮМЕМХЕ, ГЮ ХЯЙКЧВЕМХЕЛ ЯРПЮМ чФМНИ юЛЕПХЙХ. с ЮДЕМНБХПСЯЮ РХОЮ 16НОХЯЮМН 16 ЦЕМНЛМШУ БЮПХЮМРНБ, ОПХ╜ВЕЛ СВЮЯРЙХ ХГЛЕМВХБНЯРХ ПЮБМНЛЕПМН ПЮЯОПЕДЕКЪКХЯЭ ОН БЯЕ╜ЛС ЦЕМНЛС. яПЕДХ 76 ЬРЮЛЛНБ ЮДЕМНБХПСЯЮ РХОЮ 37,БШДЕКЕМ╜МШУ ГЮ ОНЯКЕДМХЕ 10 КЕР НР АНКЭМШУ МЮ РПЕУ ЙНМРХМЕМРЮУ, НА-
МЮПСФЕМН 20 ЬРЮЛЛНБ, НРКХВЮЧЫХУЯЪ ОН ТХГХВЕЯЙХЛ ЙЮПРЮЛ ЦЕМНЛМНИ дмй НР ОПНРНРХОМНЦН ЬРЮЛЛЮ. хГЛЕМЕМХЕ ТХГХВЕ╜ЯЙХУ ЙЮПР Б МЕЙНРНПШУ ЯКСВЮЪУ ЯНОПНБНФДЮКНЯЭ ЯЕПНКНЦХВЕ╜ЯЙХЛХ ПЮГКХВХЪЛХ, БШЪБКЪЕЛШЛХ Б прцю Х PH. нДМЮЙН ЩРХ ПЮГКХВХЪ АШКХ МЕБЕКХЙХ Х МЕ ЯННРБЕРЯРБНБЮКХ ПЮЯУНФДЕМХЧ, ЙНРНПНЕ НФХДЮКНЯЭ ОН ДЮММШЛ ПЕЯРПХЙЖХНММНЦН ЮМЮКХГЮ [Ad╜rian Fh. et al., 1988].
пНКЭ ОНОСКЪЖХИ Б Щ Б Н К ЧЖХ Х. мЕОНМЪРМШЛ МЮ ОЕП╜БШИ БГЦКЪД ЩБНКЧЖХНММШЛ ОПЕДЯРЮБКЕМХЕЛ б. л. фДЮМНБЮ ЪБКЪЕРЯЪ НРПХЖЮМХЕ ПЕЮКЭМНЯРХ ЯСЫЕЯРБНБЮМХЪ ОНОСКЪЖХИ С БХПСЯНБ. б МЮЯРНЪЫЕИ ЙМХЦЕ ЛНФМН ОПНВЕЯРЭ ЯКЕДСЧЫЕЕ: ╚яРНКЭ ФЕ МЕОПХЕЛКЕЛШЛ Й БХПСЯЮЛ ЪБКЪЕРЯЪ Х ОНМЪРХЕ ОН╜ОСКЪЖХХ, РЮЙ ЙЮЙ БМСРПХЙКЕРНВМЮЪ ЯРЮДХЪ ПЕОПНДСЙЖХХ, Ю РЕЛ АНКЕЕ ХМРЕЦПЮЖХНММШЕ ОПНЖЕЯЯШ МЮЖЕКН КХЬЮЧР ЯЛШЯКЮ РПЮЙ╜РНБЙС ПЕОПНДСЖХПСЧЫЕЦНЯЪ БХПСЯЮ ЙЮЙ ОНОСКЪЖХХ Х Р. Д.╩.
рПСДМНЯРХ ББЕДЕМХЪ Б БХПСЯНКНЦХЧ ОНМЪРХЪ ЙЮРЕЦНПХХ ╚ОН╜ОСКЪЖХЪ╩ КЕЦЙН ОПЕНДНКЕБЮЧРЯЪ, ЕЯКХ СВЕЯРЭ, ВРН ОНОСКЪЖХЪ ЕЯРЭ ЙЮРЕЦНПХЪ ХЯРНПХВЕЯЙЮЪ. яРПСЙРСПЮ Х ОПХМЖХОШ НПЦЮМХ╜ГЮЖХХ ОНОСКЪЖХХ ХГЛЕМЪКХЯЭ Б УНДЕ ЩБНКЧЖХХ. бХПСЯМШЕ ОН╜ОСКЪЖХХ УЮПЮЙРЕПХГСЧРЯЪ ЯБНЕНАПЮГХЕЛ Х ЯСЫЕЯРБЕММН НРКХ╜ВЮЧРЯЪ ОН ЯБНЕЛС ЯРПНЕМХЧ НР ОНОСКЪЖХИ БШЯЬХУ ЩСЙЮПХНРНБ, ЙНРНПШЕ ЯКСФЮР ЩРЮКНМНЛ ОНОСКЪЖХХ ДКЪ АНКЭЬХМЯРБЮ АХНКН╜ЦНБ [жХКХМЯЙХИ ъ- ъ., 1988]. б. л. фДЮМНБ ОПНЬЕК ЛХЛН ЩРНИ НВЕБХДМНИ ХЯРХМШ. щРН ОПЪЛН ЯБЪГЮМН Я ЯНЯРНЪМХЕЛ ЯНБПЕЛЕМ╜МНИ ЩБНКЧЖХНММНИ АХНКНЦХХ Х ЯН БГЦКЪДЮЛХ б. л. фДЮМНБЮ МЮ ДБХФСЫХЕ ЯХКШ ЩБНКЧЖХХ.
яХМРЕРХВЕЯЙЮЪ РЕНПХЪ ЩБНКЧЖХХ ОЕПЕФХБЮЕР ЦКСАНЙХИ ЙПХ╜ГХЯ. еЕ ЖЕМРПЮКЭМЮЪ ДНЦЛЮ, ЯНЦКЮЯМН ЙНРНПНИ МЮЙНОКЕМХЪ Б ОН╜ОСКЪЖХЪУ ЯКСВЮИМШУ ЛСРЮЖХИ Х ХУ ОНЯКЕДСЧЫЮЪ НЖЕМЙЮ ЕЯРЕ╜ЯРБЕММШЛ НРАНПНЛ ЪБКЪЧРЯЪ МЕНАУНДХЛШЛХ Х ДНЯРЮРНВМШЛХ СЯКНБХЪЛХ ДКЪ НЯСЫЕЯРБКЕМХЪ ЛХЙПН- Х ЛЮЙПНЩБНКЧЖХХ, СФЕ МЕ ЛНФЕР ОПЕРЕМДНБЮРЭ МЮ ЛНМНОНКХЧ Б ХГСВЕМХХ ЩБНКЧЖХНМ╜МНЦН ОПНЖЕЯЯЮ. яНБПЕЛЕММЮЪ ЩБНКЧЖХНММЮЪ РЕНПХЪ ДНКФМЮ СВХ╜РШБЮРЭ МЕОНЯРНЪМЯРБН ЦЕМНЛЮ, МЕИРПЮКЭМНЯРЭ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ Х ЛМНЦХЕ ДПСЦХЕ ЪБКЕМХЪ [Antonovics J., 1987; мН Mae-Wan1 1987; Pollard J. W., 1987; David J. R.i 1988]. нРПХЖЮ╜МХЕ б. л. фДЮМНБШЛ НВЕБХДМНЦН ТЮЙРЮ ЯСЫЕЯРБНБЮМХЪ ОНОС╜КЪЖХИ С БХПСЯНБ ЪБКЪЕРЯЪ ЯБНЕНАПЮГМШЛ ОПНРЕЯРНЛ ОПНРХБ АЕЯ╜ЯХКХЪ ЙКЮЯЯХВЕЯЙНИ ЯХМРЕРХВЕЯЙНИ РЕНПХХ ЩБНКЧЖХХ (Б НЯМНБ╜МНЛ НЦПЮМХВХБЮЧЫЕИЯЪ ПЮЯЯЛНРПЕМХЕЛ ОНОСКЪЖХНММШУ .ОПН-' ЖЕЯЯНБ) НАЗЪЯМХРЭ ПЮГБХРХЕ НПЦЮМХВЕЯЙНЦН ЛХПЮ.
нДМНБПЕЛЕММН Я ОЕПЕЯЛНРПНЛ Х ЛНДЕПМХГЮЖХЕИ ЯХМРЕРХВЕ╜ЯЙНИ РЕНПХХ ЩБНКЧЖХХ Б ЯНБПЕЛЕММНИ МЮСЙЕ ОПНХЯУНДХР ПНФ╜ДЕМХЕ МНБНИ ТХКНЯНТХХ ,АХНКНЦХХ. пЮАНРШ б. л. фДЮМНБЮ ОН ЩБНКЧЖХХ ЯОНЯНАЯРБНБЮКХ ЩРНЛС. б. л. фДЮМНБ МЮГБЮК ЯХКС, ЙНРНПЮЪ ЛНФЕР СЯЙНПХРЭ ЩБНКЧЖХНММНЕ ПЮГБХРХЕ. рЮЙНИ ЯХКНИ ЯКСФХР БХПСЯМЮЪ РПЮМЯДСЙЖХЪ. лШЯКЭ Н ПНКХ БХПСЯНБ Б ЦНПХ-
323▓
ГНМРЮКЭМНЛ ОЕПЕМНЯЕ ЦЕМЕРХВЕЯЙНИ ХМТНПЛЮЖХХ БШЯЙЮГШБЮКЮЯЭ Б НАЫЕЛ БХДЕ МЕНДМНЙПЮРМН. гЮЯКСЦЮ б. л. фДЮМНБЮ ГЮЙКЧВЮ╜ЕРЯЪ Б РНЛ, ВРН НМ, ЙЮЙ ОПНТЕЯЯХНМЮК-БХПСЯНКНЦ, БЯЕЯРНПНММЕ НАНЯМНБЮК ЩРС ХДЕЧ.
пЮГБХРХЕ РЕНПХХ ЩБНКЧЖХХ МЕ НГМЮВЮЕР ГЮАБЕМХЪ ДНЯРХФЕ╜МХИ ОПНЬКНЦН [Futuyma D. J., 1988]. мЕ ОПХГМЮБЮЪ ПЕЮКЭМНЯРЭ ЯСЫЕЯРБНБЮМХЪ БХПСЯМШУ ОНОСКЪЖХИ, б. л. фДЮМНБ ТЮЙРХВЕЯЙХ МЕ ОНПШБЮК Я ЯХМРЕРХВЕЯЙНИ РЕНПХЕИ ЩБНКЧЖХХ Х ЬХПНЙН ХЯ╜ОНКЭГНБЮК Б ЯБНХУ ПЮАНРЮУ ОНОСКЪЖХНММШИ Х ЩЙНКНЦХВЕЯЙХИ ОНДУНДШ Й ЩБНКЧЖХНММШЛ ЪБКЕМХЪЛ. пЮЯЯЛНРПХЛ ПЮГБХРХЕ ЩРН╜ЦН МЮОПЮБКЕМХЪ.
йКЮЯЯХВЕЯЙХЛХ Х ОНЯРНЪММН ДЕИЯРБСЧЫХЛХ ХЯРНВМХЙЮЛХ МЮЯКЕДЯРБЕММНИ ХГЛЕМВХБНЯРХ ДКЪ БХПСЯНБ ЪБКЪЧРЯЪ ЛСРЮЖХХ, ХЯРХММШЕ ПЕЙНЛАХМЮЖХХ Х ОЕПЕПЮЯОПЕДЕКЕМХЕ ТПЮЦЛЕМРНБ ЦЕ╜МНЛЮ. мЮЙНОКЕММШЕ ДЮММШЕ ЯБХДЕРЕКЭЯРБСЧР Н РНЛ, ВРН ЩРХ ХЯРНВМХЙХ ЯНГДЮЧР Б ОНОСКЪЖХЪУ НЦПНЛМШИ ГЮОЮЯ ЦЕМЕРХВЕЯЙНИ ХГЛЕМВХБНЯРХ Х НАЕЯОЕВХБЮЧР ЛХЙПНЩБНКЧЖХЧ. щРС ТСМЙЖХЧ ЛНЦСР БШОНКМЪРЭ МЕ РНКЭЙН ГМЮВХРЕКЭМШЕ ОН ОПНРЪФЕММНЯРХ, МН Х ЕДХМХВМШЕ РНВЙНБШЕ ЛСРЮЖХХ. й ХЛЕЧЫХЛЯЪ ДНЙЮГЮРЕКЭ╜ЯРБЮЛ БКХЪМХЪ РНВЕВМШУ ЛСРЮЖХИ МЮ ТЕМНРХО ДНАЮБХКЮЯЭ ПЮАН╜РЮ Jooh Ji-Woon Х ЯНЮБР. (1988), СЯРЮМНБХБЬХУ, ВРН ДХЮАЕРХВЕ╜ЯЙХИ БЮПХЮМР БХПСЯЮ ЩМЖЕТЮКНЛХНЙЮПДХРЮ НРКХВЮЕРЯЪ НР МЕ╜ДХЮАЕРХВЕЯЙНЦН МЮКХВХЕЛ НДМНЦН НКХЦНМСЙКЕНРХДЮ, ЙНРНПШИ МЕ БШЪБКЪЕРЯЪ МЮ Ti-НКХЦНМСЙКЕНРХДМШУ ЙЮПРЮУ МЕДХЮАЕРХВЕЯЙН╜ЦН БЮПХЮМРЮ. хГЛЕМЕМХЕ рЁ-НКХЦНМСЙКЕНРХДМНИ ЙЮПРШ МЕДХЮАЕ╜РХВЕЯЙНЦН БЮПХЮМРЮ АШКН НАСЯКНБКЕМН ЕДХМХВМНИ РНВЙНБНИ ГЮ╜ЛЕМНИ A→-C,КНЙЮКХГНБЮММНИ Б СВЮЯРЙЕ ЦЕМНЛЮ, ЯННРБЕРЯРБСЧ╜ЫЕИ СРПЮВЕММНЛС НКХЦНМСЙКЕНРХДС. дПСЦХУ ГЮЛЕМ МХ Б ЩРНЛ, МХ Б ЯЛЕФМШУ СВЮЯРЙЮУ МЕ НАМЮПСФЕМН. оНКСВЕММШЕ ПЕГСКЭРЮ╜РШ ОНГБНКЪЧР ОПЕДОНКЮЦЮРЭ, ВРН ЯСЫЕЯРБЕММНЕ ХГЛЕМЕМХЕ ОЮРН╜ЦЕММШУ ОНРЕМЖХИ БХПСЯЮ ЩМЖЕТЮКНЛХНЙЮПДХРЮ, БШПЮФЮЧЫЕЕЯЪ Б ОНРЕПЕ ЯОНЯНАМНЯРХ БШГШБЮРЭ ДХЮАЕР, ЛНФЕР АШРЭ НАСЯКНБ╜КЕМН НДМНИ РНВЙНБНИ ЛСРЮЖХЕИ РХОЮ ГЮЛЕМШ. бШГШБЮЧР ХМРЕ╜ПЕЯ РЮЙФЕ ДЮММШЕ, НРМНЯЪЫХЕЯЪ Й ПЕРПНБХПСЯЮЛ. еДХМХВМЮЪ ЛСРЮЖХЪ Б ЦЕМЕ НАНКНВЕВМШУ АЕКЙНБ, ОПХБНДЪЫЮЪ Й ГЮЛЕМЕ КХГХМЮ МЮ ЮПЦХМХМ, МЮПСЬЮЕР ХУ ОПНЖЕЯЯХМЦ Х КХЬЮЕР ЩМДН╜ЦЕММШИ НМЙНРПНОМШИ БХПСЯ КЕИЙНГЮ ЛШЬЕИ КХМХХ ягм/мЕ ЯОНЯНАМНЯРХ Й ПЕОКХЙЮЖХХ Х ПЪДЮ ДПСЦХУ ЯБНИЯРБ. нАПЮРМН МЮ╜ОПЮБКЕММЮЪ ЛСРЮЖХЪ Б ЯЮИРЕ ОПНЖЕЯЯХМЦЮ (ГЮЛЕЫЕМХЕ ЮПЦХМХ╜МЮ МЮ КХГХМ) БЕДЕР Й ПЕБЕПЯХХ Й ДХЙНЛС РХОС [Sithanandam G., Rapp U. R., 1988].
оПНДНКФЮЕРЯЪ МЮЙНОКЕМХЕ ДЮММШУ Н ПНКХ ОЕПЕПЮЯОПЕДЕКЕ╜МХЪ ТПЮЦЛЕМРНБ ЦЕМНЛЮ Б ХГЛЕМВХБНЯРХ НПАХБХПСЯНБ. J. L. Stott Х ЯНЮБР. (1987) ХГСВЮКХ НАПЮГНБЮМХЕ ПЕЮЯЯНПРЮМРНБ Б НПЦЮМХГ╜ЛЕ РЕКЪР, ХМТХЖХПНБЮММШУ ДБСЛЪ ЯЕПНРХОЮЛХ БХПСЯЮ ЯХМЕЦН ЪГШЙЮ. оПЕНАКЮДЮЧЫЕЕ ЙНКХВЕЯРБН (89%) БХПСЯМШУ ЙКНМНБ, ХГНКХПНБЮММШУ ХГ ЙПНБХ ГЮПЮФЕММШУ ФХБНРМШУ, АШКН ПЕЮЯ-
ЯНПРЮМРЮЛХ. б ХУ НАПЮГНБЮМХХ СВЮЯРБНБЮКХ 6 ХГ 10 ЯЕЦЛЕМРНБ, ХЛЕЧЫХУЯЪ Б БХПСЯМНЛ ЦЕМНЛЕ. хДЕМРХТХЖХПНБЮМН 16 БХДНБ ПЕЮЯЯНПРЮМРНБ. P. D. Oberst Х ЯНЮБР. (1987) ОНЙЮГЮКХ, ВРН ЦЕ╜МЕРХВЕЯЙНЕ БГЮХЛНДЕИЯРБХЕ ЛЕФДС ЬРЮЛЛЮЛХ БХПСЯЮ ЯХМЕЦН ЪГШЙЮ Б НПЦЮМХГЛЕ ЕЯРЕЯРБЕММШУ ╚УНГЪЕБ╩ ОПНХЯУНДХР МЕ РНКЭ╜ЙН ОПХ НЯРПНИ, МН Х ДКХРЕКЭМН РЕЙСЫЕИ ХМТЕЙЖХХ. юБРНПШ БШ╜ДЕКЪКХ ПЕЮЯЯНПРЮМРШ ХГ НПЦЮМХГЛЮ АШЙЮ МЮ ОПНРЪФЕМХХ 41 ДМЪ, Б РЕВЕМХЕ ЙНРНПШУ ОПНДНКФЮКЮЯЭ БХПЕЛХЪ. оНКСВЕММШЕ ПЕ╜ГСКЭРЮРШ СЙЮГШБЮЧР, ВРН ОЕПЕЯНПРХПНБЙЮ ЦЕМНЛМШУ ЯЕЦЛЕМРНБ БМНЯХР ГМЮВХРЕКЭМШИ БЙКЮД Б НАПЮГНБЮМХЕ ТЕМН- Х ЦЕМНРХОХ╜ВЕЯЙХУ ПЮГКХВХИ, УЮПЮЙРЕПМШУ ДКЪ ЖХПЙСКХПСЧЫХУ Б ОПХПНДЕ ЬРЮЛЛНБ БХПСЯЮ ЯХМЕЦН ЪГШЙЮ.
аКЮЦНДЮПЪ БШЯНЙНИ ЛСРЮАЕКЭМНЯРХ ЦЕМЕРХВЕЯЙНЦН ЛЮРЕПХЮ╜КЮ ЩБНКЧЖХЪ пмй-ЦЕМНЛМШУ БХПСЯНБ ОПНХЯУНДХР Б НПЦЮМХГЛЕ ГЮПЮФЕММНЦН ╚УНГЪХМЮ╩ БН БПЕЛЪ ХМТЕЙЖХНММНЦН ОПНЖЕЯЯЮ. бХ╜ПСЯ, БМЕДПХБЬХИЯЪ Б НПЦЮМХГЛ ╚УНГЪХМЮ╩, ЦЕМЕРХВЕЯЙХ НРКХВЮ╜ЕРЯЪ НР БХПСЯЮ, ЙНРНПШИ Б ОНЯКЕДСЧЫЕЛ МЮЙЮОКХБЮЕРЯЪ С ■АНКЭМНЦН. F. Gebauer Х ЯНЮБР. (1988) ОПНЯКЕДХКХ ХГЛЕМВХБНЯРЭ БХПСЯЮ ЪЫСПЮ ЯЕПНРХОЮ яГ Б НПЦЮМХГЛЕ ЙПСОМНЦН ПНЦЮРНЦН ЯЙН╜РЮ МЮ ОПНРЪФЕМХХ 539 ДМЕИ ОЕПЯХЯРЕМРМНИ ХМТЕЙЖХХ. яЙНПНЯРЭ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ БХПСЯМНЦН ЦЕМНЛЮ ЯНЯРЮБХКЮ 0,9 ? ?10~2≈7,4?10^2ГЮЛЕМ МЮ СВЮЯРНЙ Б ЦНД; 59% ЛСРЮЖХИ PHK БКЕЙКХ ГЮ ЯНАНИ ЮЛХМНЙХЯКНРМШЕ ГЮЛЕМШ. б ОПНЖЕЯЯЕ ХМТЕЙ╜ЖХХ НР АШЙЮ АШКХ ОНКСВЕМШ ХГНКЪРШ БХПСЯЮ, НРКХВЮЧЫХЕЯЪ Б ЮМРХЦЕММНЛ НРМНЬЕМХХ НР ПНДХРЕКЭЯЙНЦН ЬРЮЛЛЮ. щРХ ДЮММШЕ СЙЮГШБЮЧР, ВРН ЙПСОМШЕ ФБЮВМШЕ ФХБНРМШЕ МЕ РНКЭЙН БШОНК╜МЪЧР ПНКЭ ПЕГЕПБСЮПНБ БХПСЯЮ ЪЫСПЮ, МН И ЯКСФЮР ХЯРНВМХ╜ЙНЛ ХГЛЕМЕММШУ БЮПХЮМРНБ.
мЮЙНОКЕМХЕ ЛСРЮЖХИ Х ХГЛЕМВХБНЯРХ, ╚ОНЯРЮБКЪЕЛНИ╩ ПЕ╜ЙНЛАХМЮЖХЪЛХ, БЕДЕР Й ТНПЛХПНБЮМХЧ БМСРПХ- Х ЛЕФОНОСКЪ╜ЖХНММШУ ПЮГКХВХИ. б ЩРНЛ НРМНЬЕМХХ ВПЕГБШВЮИМН ОНЙЮГЮ╜РЕКЭМШ ДЮММШЕ Н ОНКХНБХПСЯЕ, ПЮЯЯЛНРПЕММШЕ Б НАГНПЕ ч. 3. цЕМДНМЮ (1988). хЯОНКЭГНБЮМХЕ 7ЦНКХЦНМСЙКЕНРХДМНЦН ЙЮПРХПНБЮМХЪ, НОПЕДЕКЕМХЕ ОНЯКЕДНБЮРЕКЭМНЯРЕИ МСЙКЕНРХДНБ Х ЮЛХМНЙХЯКНР, Ю РЮЙФЕ ЮМРХЦЕММШИ ЮМЮКХГ, ОПНБЕДЕММШИ Я ОНЛНЫЭЧ ОЮМЕКХ ЛНМНЙКНМЮКЭМШУ ЮМРХРЕК, ОНГБНКХКХ СЯРЮ╜МНБХРЭ, ВРН ЬРЮЛЛШ ОНКХНБХПСЯЮ, ЖХПЙСКХПСЧЫХЕ Б ПЮГКХВ╜МШУ ЦЕНЦПЮТХВЕЯЙХУ ГНМЮУ, УЮПЮЙРЕПХГСЧРЯЪ ТЕМН- Х ЦЕМНРХ╜ОХВЕЯЙНИ ХМДХБХДСЮКЭМНЯРЭЧ. бШДЕКЕМШ 10 ЦПСОО (яПЕДМЕ╜ЮГХЮРЯЙЮЪ, яЕБЕПМНИ юЛЕПХЙХ, чФМНЮТПХЙЮМЯЙЮЪ, хМДХИЯЙЮЪ Х Р. Д.). R. Rico-Hesse Х ЯНЮБР. (1987) ОПНБЕКХ ЯПЮБМХРЕКЭМНЕ ХГСВЕМХЕ ОНЯКЕДНБЮРЕКЭМНЯРЕИ ЦЕМНЛЮ, ЙНДХПСЧЫХУ СВЮЯРЙХ ЙЮОЯХДМНЦН АЕКЙЮ VPlХ МЕЙЮОЯХДМНЦН ОПНРЕХМЮ ю2 С 62 ЬРЮЛ╜ЛНБ ОНКХНБХПСЯЮ РХОЮ 1. ьРЮЛЛШ АШКХ БШДЕКЕМШ ГЮ ОНЯКЕД╜МХИ 31 ЦНД МЮ 5 ЙНМРХМЕМРЮУ НР АНКЭМШУ ОЮПЮКХРХВЕЯЙХЛ ОН╜КХНЛХЕКХРНЛ. оПНБЕДЕМЮ ЦПСООХПНБЙЮ Х БШЪБКЕМШ ТНЙСЯШ ЖХП╜ЙСКЪЖХХ ЯУНДМШУ ЬРЮЛЛНБ. сЯРЮМНБКЕМН ПЕЙНЛАХМЮМРМНЕ ОПН-
ХЯУНФДЕМХЕ (ДХЙХИ ≈ ДХЙХИ, БЮЙЖХММШИ ≈ ДХЙХИ) МЕЙНРНПШУ ХГНКЪРНБ. б ОПЮЙРХВЕЯЙНЛ НРМНЬЕМХХ БЮФМН НРЛЕРХРЭ, ВРН Б ЕЯРЕЯРБЕММШУ ОНОСКЪЖХЪУ ОНКХНБХПСЯЮ ЛНЦСР ОНЪБКЪРЭЯЪ ЮМРХ╜ЦЕММН ЮРХОХВМШЕ ЬРЮЛЛШ, ЙНРНПШЕ ОПЕНДНКЕБЮЧР ХЛЛСМНКН╜ЦХВЕЯЙХИ АЮПЭЕП, ЯНГДЮБЮЕЛШИ БЮЙЖХМЮЖХЕИ. хГЛЕМВХБНЯРЭ Б ЩРНЛ МЮОПЮБКЕМХХ ЯБЪГЮМЮ Я МЕОНКМШЛ НУБЮРНЛ МЮЯЕКЕМХЪ ХЛ╜ЛСМХГЮЖХЕИ Х МЕБШЯНЙХЛ СПНБМЕЛ ЙНККЕЙРХБМНЦН ХЛЛСМХРЕРЮ [жХКХМЯЙХИ ъ- ъ-, 1988]. оЕПЕД ГДПЮБННУПЮМЕМХЕЛ БНГМХЙЮЕР ОПНАКЕЛЮ ОПХМЪРХЪ ЛЕП ОН ГЮЫХРЕ НР ЮМРХЦЕММН .ХГЛЕМЕММШУ БЮПХЮМРНБ Х ЯНГДЮМХЧ ЯННРБЕРЯРБСЧЫХУ БЮЙЖХМ. мНБШЕ БНГ╜ЛНФМНЯРХ ДКЪ ПЕЬЕМХЪ ЩРНИ ГЮДЮВХ НРЙПШБЮЧР СЯОЕУХ Б ОН╜КСВЕМХХ ФХГМЕЯОНЯНАМШУ ЛЕФРХОНБШУ ЦХАПХДНБ ОНКХНБХПСЯЮ ЦЕММН-ХМФЕМЕПМШЛХ ЛЕРНДЮЛХ [HogleJ. M., 1988].
б ОПНЖЕЯЯЕ ЩБНКЧЖХХ БХПСЯМШЕ ОНОСКЪЖХХ ОПХЯОНЯЮАКХБЮ╜ЧРЯЪ Й ЯПЕДЕ НАХРЮМХЪ, ГЮМХЛЮЧР НОПЕДЕКЕММСЧ ЩЙНКНЦХВЕ╜ЯЙСЧ МХЬС Х ПЮЯУНДЪРЯЪ ЦЕМЕРХВЕЯЙХ. бЮФМШЕ ДЮММШЕ ОНКС╜ВЕМШ ОПХ ХГСВЕМХХ БХПСЯНБ ПНДЮ HantaanvirusЯЕЛЕИЯРБЮ Bu- nyaυiridae [Le Due J. W., 19871. пНД БЙКЧВЮЕР Б ЯЕАЪ БНГАСДХ╜РЕКЪ ЙНПЕИЯЙНИ ЦЕЛНППЮЦХВЕЯЙНИ КХУНПЮДЙХ, БХПСЯ УЮМРЮЮМ, БШДЕКЕММШИ Б 1978 Ц. Б йНПЕЕ, Х УЮМРЮЮМ-ОНДНАМШЕ БХПСЯШ, ЙН╜РНПШЕ ПЮГДЕКЪЧРЯЪ МЮ 3 ЯЕПНКНЦХВЕЯЙХЕ ЦПСООШ. бЯЕ МЮГБЮМ╜МШЕ БХПСЯШ ЯУНДМШ ОН ЯБНЕИ ЩЙНКНЦХХ, ЯОНЯНАС ОЕПЕДЮВХ Х ЛЕ╜УЮМХГЛС БШФХБЮМХЪ Б ОПХПНДЕ. нМХ БШГШБЮЧР С ЦПШГСМНБ УПНМХВЕЯЙСЧ ЮЯХЛОРНЛЮРХВЕЯЙСЧ ХМТЕЙЖХЧ. мЕЯЛНРПЪ МЮ БШ╜ЯНЙХИ ХЛЛСММШИ НРБЕР, БХПСЯ ЛНФЕР ЯНУПЮМЪРЭЯЪ Б НПЦЮМХГЛЕ ╚УНГЪХМЮ╩ ОНФХГМЕММН. бХПСЯ БШДЕКЪЕРЯЪ Х ПЮЯОПНЯРПЮМЪЕРЯЪ БН БМЕЬМЕИ ЯПЕДЕ ЯН ЯКЧМНИ, ЛНВНИ Х ТЕЙЮКХЪЛХ ЦПШГСМНБ; ГЮ ЯВЕР ЩРНЦН ЛЕУЮМХГЛЮ ОЕПЕДЮЕРЯЪ Б ОНОСКЪЖХЪУ ЦПШГСМНБ ЦН╜ПХГНМРЮКЭМН. бХПСЯ УЮМРЮЮМ Х УЮМРЮЮМ-ОНДНАМШЕ БХПСЯШ ЯКС╜ФЮР ОПХВХМНИ ПЮГКХВМШУ ЦЕЛНППЮЦХВЕЯЙХУ КХУНПЮДНЙ С ВЕКН╜БЕЙЮ. гЮПЮФЕМХЕ ВЕКНБЕЙЮ НЯСЫЕЯРБКЪЕРЯЪ ЮЩПНЦЕММШЛ, Ю РЮЙ╜ФЕ ЙНМРЮЙРМШЛ ОСРЕЛ Х ВЕПЕГ СЙСЯ ЦПШГСМНБ.
уЮМРЮЮМБХПСЯШ ХЛЕЧР ЦКНАЮКЭМНЕ ПЮЯОПНЯРПЮМЕМХЕ Х Б ГЮ╜БХЯХЛНЯРХ НР ГЮМХЛЮЕЛНИ ЦЕНЦПЮТХВЕЯЙНИ ГНМШ ПЮГКХВЮЧРЯЪ ЯЕПНКНЦХВЕЯЙХ, ОН ЯРЕОЕМХ БХПСКЕМРМНЯРХ ДКЪ ВЕКНБЕЙЮ Х ДП. бХПСЯ УЮМРЮЮМ Х АКХГЙХЕ Й МЕЛС ЬРЮЛЛШ ЖХПЙСКХПСЧР Б йХ╜РЮЕ, йНПЕЕ, МЮ дЮКЭМЕЛ бНЯРНЙЕ, Ю РЮЙФЕ МЮ ЧЦЕ яХАХПХ Х Б еБПНОЕИЯЙНИ ВЮЯРХ яяяп. с ВЕКНБЕЙЮ НМ БШГШБЮЕР РЪФЕКСЧ ЦЕЛНППЮЦХВЕЯЙСЧ КХУНПЮДЙС Я ОНВЕВМШЛ ЯХМДПНЛНЛ, ЯНОПНБНФ╜ДЮЧЫСЧЯЪ БШЯНЙНИ ЯЛЕПРМНЯРЭЧ. б ОПНРХБНОНКНФМНЯРЭ ЩРНЛС УЮМРЮЮМ-ОНДНАМШЕ БХПСЯШ, ЖХПЙСКХПСЧЫХЕ Б яЙЮМДХМЮБХХ, гЮОЮДМНИ Х жЕМРПЮКЭМНИ еБПНОЕ, Ю РЮЙФЕ Б МЕЙНРНПШУ ПЮИН╜МЮУ еБПНОЕИЯЙНИ ВЮЯРХ яяяп (ОПНРНРХОМШИ ЬРЮЛЛ Puumala), ЯКСФЮР ОПХВХМНИ НРМНЯХРЕКЭМН КЕЦЙН ОПНРЕЙЮЧЫЕЦН Х ДНАПН╜ЙЮВЕЯРБЕММНЦН ГЮАНКЕБЮМХЪ ≈ ЦЕЛНППЮЦХВЕЯЙНИ МЕТПНОЮРХХ. уЮМРЮЮМ-ОНДНАМШЕ БХПСЯШ, ПЮЯОПНЯРПЮМЕММШЕ Б яью Х йЮМЮ╜ДЕ (ОПНРНРХОМШИ ЬРЮЛЛ Prospect Hill),МЕ НАКЮДЮЧР БХПС╜
КЕМРМНЯРЭЧ ДКЪ ВЕКНБЕЙЮ. оПХВХМНИ СЙЮГЮММШУ ПЮГКХВХИ, ОН╜ЛХЛН ЦЕНЦПЮТХВЕЯЙНИ ХГНКЪЖХХ, ЪБКЪЕРЯЪ ДЕИЯРБХЕ ЩЙНКНЦХВЕ╜ЯЙНЦН ТЮЙРНПЮ ≈ ХЯОНКЭГНБЮМХЕ ПЮГКХВМШУ ╚УНГЪЕБ╩. ╚уНГЪХ╜МНЛ╩ БХПСЯЮ УЮМРЮЮМ ЯКСФЮР ЛШЬХ Apodemus agrarius,БХПСЯЮ Puumala ≈ НАШЙМНБЕММЮЪ ОНКЕБЙЮ Clethrionomys glareolus,Ю БХПСЯ Prospect HillЩЙНКНЦХВЕЯЙХ ЯБЪГЮМ Я ЦПШГСМЮЛХ Microtus Californicus.мЮ ГМЮВЕМХЕ ЩЙНКНЦХВЕЯЙНЦН ТЮЙРНПЮ Б ДХТТЕПЕМ╜ЖХПНБЙЕ УЮМРЮЮМБХПСЯНБ СЙЮГШБЮЕР РНР ТЮЙР, ВРН ЮПЕЮКШ Apo- demus agrariusХ Clethrionomys glareolusБ еБПНОЕ ОЕПЕЙПШ╜БЮЧРЯЪ. б ПЕГСКЭРЮРЕ МЮ НДМНИ Х РНИ ФЕ РЕППХРНПХХ ЖХПЙСКХ╜ПСЧР ЬРЮЛЛШ БХПСЯНБ, ЙНРНПШЕ ПЮГКХВЮЧРЯЪ ЙЮЙ ЯЕПНКНЦХ╜ВЕЯЙХ, РЮЙ Х ОН ОЮРНЦЕММШЛ ОНРЕМЖХЪЛ ДКЪ ВЕКНБЕЙЮ.
уЮМРЮЮМБХПСЯШ МЕ ЪБКЪЧРЯЪ ЕДХМЯРБЕММНИ БХПСЯМНИ ЦПСО╜ОНИ, ОПЕДЯРЮБХРЕКХ ЙНРНПНИ ПЮГНЬКХЯЭ ЛЕФДС ЯНАНИ Б ПЕГСКЭ╜РЮРЕ МЕ РНКЭЙН ЦЕНЦПЮТХВЕЯЙНИ, МН Х ЩЙНКНЦХВЕЯЙНИ ХГНКЪЖХХ. бЮФМШЕ ДЮММШЕ ОНКСВЕМШ Б НРМНЬЕМХХ БХПСЯНБ ПНДЮ Bunya- virusesЯЕЛЕИЯРБЮ Bunyaviridae [Calisher Ch. H., 1988]. нРМН╜ЯЪЫХЕЯЪ Й ЩРНЛС ПНДС БХПСЯШ ЮМРХЦЕММНИ ЦПСООШ CВЕРЙН ДХТ╜ТЕПЕМЖХПСЧРЯЪ ДПСЦ НР ДПСЦЮ ЯЕПНКНЦХВЕЯЙХ Б прцю, PCK Х PH Х ПЮГКХВЮЧРЯЪ ОН ОЕПБХВМНИ ЯРПСЙРСПЕ ОНБЕПУМНЯРМНЦН ЦКХ╜ЙНОПНРЕХДЮ G.хУ ЮПЕЮК БЙКЧВЮЕР аПЮГХКХЧ Х ПЪД ДПСЦХУ ЯРПЮМ чФМНИ юЛЕПХЙХ. бХПСЯШ ЦПСООШ CЯОНЯНАМШ Й ОЕПЕ╜ПЮЯОПЕДЕКЕМХЧ ТПЮЦЛЕМРНБ ЦЕМНЛЮ Х ОНРЕМЖХЮКЭМН НАПЮГСЧР ЕДХМШИ ЦЕММШИ ОСК. рЕЛ МЕ ЛЕМЕЕ ЦЕМЕРХВЕЯЙЮЪ ХМДХБХДСЮКЭ╜МНЯРЭ ВКЕМНБ ЦПСООШ ЯНУПЮМЪЕРЯЪ. щРН ДНЯРХЦЮЕРЯЪ РЕЛ, ВРН ЙЮФДШИ БХПСЯ ГЮМХЛЮЕР ЯБНЧ ЩЙНКНЦХВЕЯЙСЧ МХЬС, Б ЯХКС ВЕЦН ОСРХ ХУ ЖХПЙСКЪЖХХ Б ОНГБНМНВМШУ ╚УНГЪЕБЮУ╩ Х ОЕПЕМНЯВХ╜ЙЮУ МЕ ЯЙПЕЫХБЮЧРЯЪ. щЙНКНЦХВЕЯЙЮЪ ХГНКЪЖХЪ УЮПЮЙРЕПМЮ РЮЙФЕ ДКЪ ЛМНЦХУ АСМЭЪБХПСЯНБ, БЯРПЕВЮЧЫХУЯЪ МЮ НДМНИ РЕП╜ПХРНПХХ Х НРМНЯЪЫХУЯЪ Й ЮМРХЦЕММШЛ ЦПСООЮЛ цСЮЛЮ, оЮРНХЯ, яХЛАС Х йЮКХТНПМХЪ.
мЕНДМНПНДМНЯРЭ ОНОСКЪЖХИ БХПСЯНБ Х ХУ АШЯРПЮЪ ЛХЙПН╜ЩБНКЧЖХЪ НРМНЯЪРЯЪ Й ВХЯКС НАЫХУ ЪБКЕМХИ, УЮПЮЙРЕПМШУ ДКЪ БХПСЯНБ ПЮГКХВМШУ ЯХЯРЕЛЮРХВЕЯЙХУ ЦПСОО. бЛЕЯРЕ Я РЕЛ ДЮФЕ С АКХГЙХУ БХПСЯНБ ЩРНР ОПНЖЕЯЯ ХЛЕЕР ЯБНЧ ЯОЕЖХТХЙС. л. Ya- mashita Х ЯНЮБР. (1988) ХГСВХКХ НЯНАЕММНЯРХ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ БХПСЯЮ ЦПХООЮ б Х ГЮЙНМНЛЕПМНЯРХ ЯЛЕМШ ЬРЮЛЛНБ ГЮ ОНЯКЕДМХЕ 40 КЕР.
C ЩРНИ ЖЕКЭЧ АШКЮ НОПЕДЕКЕМЮ ОЕПБХВМЮЪ ЯРПСЙРСПЮ ЦЕМЮ ЦЕЛЮЦЦКЧРХ╜МХМЮ (мю) Х ЦЕМЮ МЕЯРПСЙРСПМШУ АЕКЙНБ (NS)С БХПСЯМШУ ХГНКЪРНБ, ОНКС╜ВЕММШУ Б 1940≈1987 ЦЦ. дКЪ ЯЕЙБЕМХПНБЮМХЪ ХЯОНКЭГНБЮКХ дмй-ЙНОХХ ЯННРБЕРЯРБСЧЫХУ ЯЕЦЛЕМРНБ ЦЕМНЛЮ. щБНКЧЖХНММНЕ ПЮЯЯРНЪМХЕ ЛЕФДС ХГН╜КХРЮЛХ НОПЕДЕКЪКХ ХЯУНДЪ ХГ ВХЯКЮ МСЙКЕНРХДМШУ ГЮЛЕМ Б СЙЮГЮММШУ ЦЕ╜МЮУ. пЕГСКЭРЮРШ ЯПЮБМХБЮКХ Я ЮМЮКНЦХВМШЛХ ДЮММШЛХ, ХЛЕЧЫХЛХЯЪ Б КХ╜РЕПЮРСПЕ Б НРМНЬЕМХХ БХПСЯНБ ЦПХООЮ ю Х я.
йЮЙ ЯКЕДСЕР ХГ ПХЯ. 65, МЮ ЙНРНПНЛ ОПЕДЯРЮБКЕМШ ДЮММШЕ, ОНКСВЕММШЕ ГЮ 7 ОНЯКЕДНБЮРЕКЭМШУ ЩОХДЯЕГНМНБ, ГЮЙНМНЛЕПМН-
пХЯ. 65. пЮГАПНЯ ОПХГМЮЙНБ Х ЯЛЕМЮ ЬРЮЛЛНБ Б ОНОСКЪЖХЪУ БХПСЯНБ ЦПХООЮ ю, б Х C[,ОН Yamashita л. et al., 1988]. рНВЙХ, КЕФЮЫХЕ МЮ ЦНПХГНМРЮКЭМНИ ОКНЯЙНЯРХ, ЯННРБЕРЯРБСЧР ЬРЮЛЛЮЛ, БШДЕКЕММШЛ Б. НДХМ Х РНР ФЕ ЩОХДЯЕ- ГНМ. оПНРЪФЕММНЯРЭ КХМХИ, ЯНЕДХМЪЧЫХУ ЩРХ РНВЙХ, ЯННРБЕРЯРБСЕР ЩБНКЧ╜ЖХНММНЛС ПЮЯЯРНЪМХЧ ЛЕФДС ЬРЮЛЛЮЛХ.
ЯРХ ДБХФЕМХЪ ЦЕМЕРХВЕЯЙНЦН ЛЮРЕПХЮКЮ Б ОНОСКЪЖХЪУ БХПСЯНБ ЦПХООЮ ю, б Х CПЮГКХВЮКХЯЭ. пНДНМЮВЮКЭМХЙНЛ ЬРЮЛЛНБ, ОПЕ╜НАКЮДЮЧЫХУ Б РНР ХКХ ХМНИ ЩОХДЯЕГНМ, ДКЪ БХПСЯЮ ЦПХООЮ ю ЯКСФХР КХЬЭ НДХМ ПНДХРЕКЭЯЙХИ ЬРЮЛЛ, Ю ДКЪ БХПСЯНБ ЦПХО╜ОЮ б Х C ≈ МЕЯЙНКЭЙН. щРН ЪБКЕМХЕ НЯНАЕММН БШПЮФЕМН Б ЯКС╜ВЮЕ БХПСЯЮ ЦПХООЮ я. б ПЕГСКЭРЮРЕ Б ОНОСКЪЖХЪУ БХПСЯНБ ЯН╜ГДЮЕРЯЪ ПЮГКХВМШИ ПЮГАПНЯ ОПХГМЮЙНБ. б ЙЮФДШИ ЩОХДЯЕГНМ ОНОСКЪЖХЪ БХПСЯЮ ЦПХООЮ ю ОПЕДЯРЮБКЕМЮ НДМНИ, Ю ОНОСКЪЖХХ БХПСЯНБ б Х C ≈ МЕЯЙНКЭЙХЛХ ПНДНЯКНБМШЛХ. б ОНЯКЕДМЕЛ ЯКС╜ВЮЕ БШФХБЮЕР ДБЕ КХМХХ ХКХ АНКЕЕ. хГ ПЕГСКЭРЮРНБ ЯПЮБМЕМХЪ ОПНРЪФЕММНЯРХ БЕРБЕИ, ЯНЕДХМЪЧЫХУ ЬРЮЛЛШ, ЯКЕДСЕР, врн ЩБНКЧЖХЪ БХПСЯЮ ЦПХООЮ б ХДЕР ЛЕДКЕММЕИ, ВЕЛ БХПСЯЮ ЦПХО╜ОЮ ю, МН АШЯРПЕИ, ВЕЛ БХПСЯЮ ЦПХООЮ я,
оНПЪДНЙ ЯЛЕМШ ЬРЮЛЛНБ С БХПСЯЮ ЦПХООЮ ю, ОН-БХДХЛНЛС, НАСЯКНБКЕМ АНКЕЕ БШЯНЙНИ ЛСРЮАЕКЭМНЯРЭЧ БХПСЯМНЦН ЦЕМНЛЮ, Ю РЮЙФЕ ЛЕМЭЬЕИ ОПНДНКФХРЕКЭМНЯРЭЧ ФХГМЕММНЦН ЖХЙКЮ Х АНКЕЕ АШЯРПНИ ЯЛЕМНИ ОНЙНКЕМХИ ГЮ ЕДХМХЖС БПЕЛЕМХ Б ЛЮЯ╜ЬРЮАЕ ОНОСКЪЖХХ. щРН ЯОНЯНАЯРБСЕР МЮЙНОКЕМХЧ ЛСРЮМРНБ, ЯПЕДХ ЙНРНПШУ ХЛЕЧРЯЪ ЕДХМХВМШЕ ЛСРЮМРШ, ЯОНЯНАМШЕ ОПЕ╜НДНКЕРЭ ЙНККЕЙРХБМШИ ХЛЛСМХРЕР. нМХ НАЕЯОЕВХБЮЧР ОПНДНК╜ФЕМХЕ ПНДЮ Х ЯСЫЕЯРБНБЮМХЕ БХПСЯЮ БН БПЕЛЕМХ. нЯРЮКЭМЮЪ ВЮЯРЭ ПНДХРЕКЭЯЙНИ ОНОСКЪЖХХ СРПЮВХБЮЕРЯЪ. бХПСЯШ ЦПХООЮ б Х CХЯОШРШБЮЧР ЛЕМЭЬЕЕ ХЛЛСММНЕ ДЮБКЕМХЕ. щРН, МЕЯЛНРПЪ
МЮ ЛЕМЭЬХИ ЯОЕЙРП ЮМРХЦЕММНИ ХГЛЕМВХБНЯРХ, НАСЯКНБКЕММНИ НРМНЯХРЕКЭМН МХГЙНИ ЛСРЮАЕКЭМНЯРЭЧ, НАЕЯОЕВХБЮЕР ОПНДНКФХ╜РЕКЭМСЧ ЖХПЙСКЪЖХЧ ЬРЮЛЛНБ МЮ ОПНДНКФЕМХХ МЕЯЙНКЭЙХУ ЩОХДЯЕГНМНБ. бХПСЯ ЦПХООЮ б ХГЛЕМЪЕРЯЪ Б 2≈6 ПЮГ ЛЕДКЕММЕИ, ВЕЛ БХПСЯ ЦПХООЮ ю. оНЙЮГЮРЕКЕЛ МХГЙНИ ЛСРЮАЕКЭМНЯРХ БХПС╜ЯЮ ЦПХООЮ CЪБКЪЧРЯЪ НРЯСРЯРБХЕ ЛСРЮЖХИ Б ЦЕМЕ NSС БХПСЯ╜МШУ ХГНКЪРНБ, ПЮГДЕКЕММШУ ХМРЕПБЮКНЛ Б 19 КЕР, Х ХГЛЕМЕМХЪ РНКЭЙН ОН ДБСЛ МСЙКЕНРХДЮЛ Б ЦЕМЕ мю С ХГНКЪРНБ, ОНКСВЕМ╜МШУ Я ХМРЕПБЮКНЛ Б 31 ЦНД. лНКЕЙСКЪПМЮЪ ЩБНКЧЖХЪ БХПСЯЮ ЦПХООЮ б ОПНРЕЙЮЕР ЯН ЯЙНПНЯРЭЧ 1,1 ? IO-3МСЙКЕНРХДМШУ ГЮ╜ЛЕМ МЮ СВЮЯРНЙ Б ЦНД.
щЙНКНЦХВЕЯЙЮЪ МХЬЮ, ЙНРНПСЧ ГЮМХЛЮЧР БХПСЯШ, БЙКЧВЮ╜ЕР Б ЯЕАЪ МЕ РНКЭЙН НПЦЮМХГЛ Х ОНОСКЪЖХЧ ╚УНГЪХМЮ╩, МН Х БНЯОПХХЛВХБШЕ ЙКЕРЙХ, Б ЙНРНПШУ ОПНХЯУНДХР ПЕОПНДСЙЖХЪ БХ╜ПСЯНБ. дНКЦНЕ БПЕЛЪ ЯВХРЮКНЯЭ, ВРН БХДНБЮЪ ЯОЕЖХТХВМНЯРЭ БХПСЯЮ ОН НРМНЬЕМХЧ Й ╚УНГЪХМС╩ Х БШАНП НОПЕДЕКЕММШУ ЙКЕ╜РНЙ-ЛХЬЕМЕИ Б НПЦЮМХГЛЕ ОНЯКЕДМЕЦН НАЕЯОЕВХБЮЧРЯЪ ЯОЕЖХ╜ТХВМНЯРЭЧ БГЮХЛНДЕИЯРБХЪ БХПСЯЮ Я ЙКЕРНВМШЛХ ПЕЖЕОРНПЮЛХ. оНГДМЕЕ АШКХ ОНКСВЕМШ ДНЙЮГЮРЕКЭЯРБЮ РНЦН, ВРН НЦПЮМХВЕ╜МХЕ ЙПСЦЮ ╚УНГЪЕБ╩ НАСЯКНБКЕМН РЮЙФЕ СЯКНБХЪЛХ ПЕОПНДСЙ╜ЖХХ БХПСЯЮ Б ВСБЯРБХРЕКЭМШУ ЙКЕРЙЮУ. хГ МНБШУ ПЮАНР, ОНЯБЪ╜ЫЕММШУ ЩРНИ ОПНАКЕЛЕ, ЛНФМН БШДЕКХРЭ ЯННАЫЕМХЕ J. Bene- vente Х A. Shatkin (1988), ХГСВЮБЬХУ ОПХВХМС МЕЯОНЯНАМНЯРХ ПЕНБХПСЯЮ ОРХЖ Й ПЕОПНДСЙЖХХ Б ЙКЕРЙЮУ ЛШЬХ. бХПСЯ ЯОЕЖХ╜ТХВЕЯЙХ БГЮХЛНДЕИЯРБНБЮК Я ЙСКЭРСПНИ ЛШЬХМШУ L-ЙКЕРНЙ Х 'ОПНМХЙЮК Б ЖХРНОКЮГЛС, Б ЙНРНПНИ ОПНХЯУНДХКН НАПЮГНБЮМХЕ БХПСЯМШУ РПЮМЯЙПХОРНБ. нДМЮЙН БХПСЯМШЕ^ Лпмй МЕ ЯБЪГШБЮ╜КХЯЭ Я ОНКХЯНЛЮЛХ, Х ЯХМРЕГ БХПСЯМШУ АЕКЙНБ МЕ НЯСЫЕЯРБКЪК╜ЯЪ. хМТЕЙЖХНММШИ ЖХЙК ОПЕПШБЮКЯЪ. бХПСЯЯОЕЖХТХВЕЯЙХЕ пмй, ЯХМРЕГХПСЧЫХЕЯЪ Б ю-ЙКЕРЙЮУ, АШКХ ТСМЙЖХНМЮКЭМН ЮЙ╜РХБМШ Х РПЮМЯКХПНБЮКХЯЭ МЮ АЕКЙХ Б СЯКНБХЪУ in vitroБ ПЕРХ╜ЙСКНЖХРЮУ ЙПНКХЙЮ. рЮЙХЛ НАПЮГНЛ, ПЕОПНДСЙЖХЪ ПЕНБХПСЯЮ ОРХЖ Б ЙКЕРЙЮУ ЛШЬХ АКНЙХПНБЮКЮЯЭ МЮ ЯРЮДХХ ХМХЖХЮЖХХ ЯХМРЕГЮ АЕКЙНБ.
нДМХЛ ХГ ОСРЕИ ЩБНКЧЖХХ БХПСЯНБ ЪБКЪЕРЯЪ ПЮЯУНФДЕМХЕ Б НПЦЮМХГЛЕ ╚УНГЪХМЮ╩. рЮЙНИ ОСРЭ ОПНДЕКЮКХ ЦЕПОЕРХВЕЯЙХЕ БХ╜ПСЯШ ВЕКНБЕЙЮ, ЙНРНПШЕ ХЯОНКЭГСЧР ПЮГКХВМШЕ ЙКЕРЙХ-ЛХЬЕ╜МХ, ХЛЕЧР НОПЕДЕКЕММШИ РЙЮМЕБШИ РПНОХГЛ Х ПЮГКХВЮЧРЯЪ ОН ОЮРНЦЕМЕГС БШГШБЮЕЛШУ ХЛ ГЮАНКЕБЮМХИ. рЙЮМЕБШИ РПНОХГЛ ЩРХУ БХПСЯНБ Б ГМЮВХРЕКЭМНИ ЯРЕОЕМХ НАЕЯОЕВХБЮЕРЯЪ АХНУХ╜ЛХВЕЯЙХЛХ ЛЕУЮМХГЛЮЛХ: ЛНДХТХЙЮЖХЕИ ПЕЦСКЪЖХХ РПЮМЯЙПХО╜ЖХХ, ОНЯРРПЮМЯКЪЖХНММШЛ ХГЛЕМЕМХЕЛ ЯРПСЙРСПМШУ АЕКЙНБ Х ЮЙРХБМНЯРЭЧ ПЕЦСКЪРНПМШУ ЦЕМНБ. щРХ ЯБНИЯРБЮ ОПХНАПЕРЕМШ Б ОПНЖЕЯЯЕ ЩБНКЧЖХХ Х БМНЯЪР ЯБНИ БЙКЮД Б ЯОЕЖХТХВМНЯРЭ БГЮХ╜ЛНДЕИЯРБХЪ БХПСЯЮ Я ЙКЕРЙЮЛХ [Mach л. et al., 1988].
оПХБЕДЕММШЕ ДЮММШЕ ОНЙЮГШБЮЧР, ВРН Б ЛХЙПНЩБНКЧЖХХ БХПСЯМШУ ОНОСКЪЖХИ ОПНЯКЕФХБЮЧРЯЪ ЩКЕЛЕМРШ ЦПЮДСЮКХЯРХ╜
ВЕЯЙНЦН ТНПЛННАПЮГНБЮМХЪ Х БЮФМСЧ ПНКЭ ДКЪ ЕЦН НЯСЫЕЯРБКЕ╜МХЪ ХЦПЮЕР ЩЙНКНЦХВЕЯЙХИ ТЮЙРНП. б ЖЕКНЛ ЩРН СЙКЮДШБЮЕРЯЪ Б ЙНМЖЕОЖХЧ ЯХМРЕРХВЕЯЙНИ РЕНПХХ ЩБНКЧЖХХ Н ЛЕДКЕММНЛ МЮ╜ЙНОКЕМХХ МЕГМЮВХРЕКЭМШУ ХГЛЕМЕМХИ Х ОНЯРЕОЕММНЛ ПЮЯУНФ╜ДЕМХХ ОНОСКЪЖХИ, ГЮМХЛЮЧЫХУ ЯБНХ ЩЙНКНЦХВЕЯЙХЕ МХЬХ. нЯН╜АЕММНЯРХ ЦПЮДСЮКХЯРХВЕЯЙНИ ЩБНКЧЖХХ БХПСЯНБ, НАСЯКНБКЕММШЕ ЩБНКЧЖХНММШЛХ БНГЛНФМНЯРЪЛХ ЦЕМНЛЮ Х ЯРПСЙРСПНИ ОНОСКЪ╜ЖХИ БХПСЯНБ, НАЯСФДЮКХЯЭ ПЮМЕЕ [жХКХМЯЙХИ ъ- ъ-, 1988]. цПЮ╜ДСЮКХЯРХВЕЯЙНЕ ТНПЛННАПЮГНБЮМХЕ МЕ ЪБКЪЕРЯЪ ЕДХМЯРБЕММШЛ ОСРЕЛ, ОН ЙНРНПНЛС ЯКЕДСЧР БХПСЯШ. щБНКЧЖХЪ БХПСЯНБ ОПН╜ХЯУНДХР РЮЙФЕ Б ПЕГСКЭРЮРЕ БГПШБНБ, ЯЙЮВЙНБ, ЯЮКЭРЮЖХНММН. нДМЮ ХГ БНГЛНФМНЯРЕИ ДКЪ ЩРНЦН НРЙПШБЮЕРЯЪ АКЮЦНДЮПЪ МЕИ╜РПЮКЭМНЯРХ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ.
гМЮВЕМХЕ РЕНПХХ МЕИРПЮКЭМНЯРХ ЛНКЕЙСКЪП╜МНИ ЩБНКЧЖХХ ДКЪ БХПСЯНКНЦХХ. б. л. фДЮМНБ АШЯР╜ПН БНЯОПХМХЛЮК МНБШЕ ХДЕХ ЩБНКЧЖХНММНЦН СВЕМХЪ. нМ МЕ ОПНЬЕК ЛХЛН РЕНПХХ МЕИРПЮКЭМНЯРХ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ л. Kimura (1985), ОНЯРСКЮРШ ЙНРНПНИ ХГКНФЕМШ Б МЮЯРНЪЫЕИ ЙМХЦЕ.
нЦПНЛМШИ ТЮЙРХВЕЯЙХИ ЛЮРЕПХЮК, МЮЙНОКЕММШИ Б БХПСЯН╜КНЦХХ, ОНДРБЕПФДЮЕР ЙНМЖЕОЖХЧ л. йХЛСПШ Х ЯБХДЕРЕКЭЯРБС╜ЕР, ВРН ЛНКЕЙСКЪПМЮЪ ЩБНКЧЖХЪ БХПСЯНБ ХДЕР ОН ОСРХ МЕИРПЮ╜КХГЛЮ. щРН НРМНЯХРЯЪ МЕ РНКЭЙН Й дмй-ЦЕМНЛМШЛ, МН Х PHK- ЦЕМНЛМШЛ БХПСЯЮЛ. дКЪ СРБЕПФДЕМХЪ МЕИРПЮКХЯРЯЙНИ РЕНПХХ ОНЯКЕДМЕЕ НЯНАЕММН БЮФМН. щРЮ РЕНПХЪ АШКЮ ПЮГПЮАНРЮМЮ МЮ НЯМНБЕ ТЮЙРХВЕЯЙХУ ДЮММШУ, ОНКСВЕММШУ ОПХ ХГСВЕМХХ НПЦЮ╜МХГЛНБ, ЙНРНПШЕ ХЯОНКЭГСЧР Б ЙЮВЕЯРБЕ ЦЕМЕРХВЕЯЙНЦН ЛЮРЕ╜ПХЮКЮ дмй- уЮПЮЙРЕП ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ пмй-ЦЕМНЛМШУ БХПСЯНБ ОНЙЮГШБЮЕР, ВРН ОПХМЖХОШ МЕИРПЮКХГЛЮ СМХБЕПЯЮКЭМШ Х ПЮЯОПНЯРПЮМЪЧРЯЪ МЮ БЯЧ АХНЯТЕПС.
хЯОНКЭГНБЮМХЕ пмй-ЦЕМНЛМШУ БХПСЯНБ ЙЮЙ НАЗЕЙРЮ ХЯЯКЕ╜ДНБЮМХИ НРЙПШКН МНБШЕ БНГЛНФМНЯРХ ДКЪ ОНГМЮМХЪ ГЮЙНМН╜ЛЕПМНЯРЕИ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ. нАЫХИ УНД ПЮГБХРХЪ ФХГ╜МХ МЮ гЕЛКЕ ОПНЯКЕФЕМ МЮ ОПНРЪФЕМХХ 3,5 ЛКПД КЕР. лСРЮ╜ЖХНММШЕ ХГЛЕМЕМХЪ МЮЙЮОКХБЮЧРЯЪ Б .ЦЕМНЛЕ Х Б АЕКЙЮУ БЯЕУ НПЦЮМХГЛНБ ХГ ОНЙНКЕМХЪ Б ОНЙНКЕМХЕ. аКЮЦНДЮПЪ МХГЙНИ ЛС- РЮАЕКЭМНЯРХ дмй ЯЙНПНЯРЭ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ НПЦЮМХГ╜ЛНБ, ХЯОНКЭГСЧЫХУ ЩРНР ЦЕМЕРХВЕЯЙХИ ЛЮРЕПХЮК, НРМНЯХРЕКЭМН МЕБЕКХЙЮ. дКЪ АЕКЙНБ ЛКЕЙНОХРЮЧЫХУ НОПЕДЕКЕМШ ЯКЕДСЧЫХЕ БЕКХВХМШ: ХМЯСКХМ ≈ 0,44, ЛХНЦКНАХМ ≈ 0,89, a-ЖЕОЭ ЦЕЛНЦКН╜АХМЮ≈ 1,2, КХГНЖХЛ ≈ 2,0, ОЮМЙПЕЮРХВЕЯЙЮЪ пмйЮГЮ ≈ 2,1, ТХАПХМНОЕОРХДШ ≈ 8,3?10~9МЮ СВЮЯРНЙ Б ЦНД Х Р. Д. яЙНПНЯРЭ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ ЦЕМНБ ОЮОНБЮБХПСЯНБ, ЙНДХПСЧЫХУ ЛЮКШИ цЮМРХРЕМ, АНКЭЬНИ р-ЮМРХЦЕМ Х АЕКЙХ VPlХ VP2,ЯН╜ЯРЮБКЪЕР 4,1 X IO-9≈6,5?10-9МЮ СВЮЯРНЙ Б ЦНД. йПЮРЙНБПЕЛЕМ╜МНЯРЭ ВЕКНБЕВЕЯЙНЦН ЯСЫЕЯРБНБЮМХЪ МЕ ОНГБНКЪЕР МЕОНЯПЕДЯР╜БЕММН МЮАКЧДЮРЭ ЩРХ ОПНЖЕЯЯШ. дКЪ БНЯЯРЮМНБКЕМХЪ УНДЮ ЛН╜
КЕЙСКЪПМНИ ЩБНКЧЖХХ ОПХАЕЦЮЧР Й ЙНЯБЕММНЛС ЛЕРНДС ≈ Й ЯПЮБМЕМХЧ НПЦЮМХГЛНБ, ОПНХЯУНДЪЫХУ ХГ НАЫЕЦН ЙНПМЪ Х ПЮГНЬЕДЬХУЯЪ ЩБНКЧЖХНММН. оН ЛЮЯЬРЮАС ПЮГКХВХИ ЯСДЪР Н БПЕЛЕМХ ОНЪБКЕМХЪ НПЦЮМХГЛНБ Х Н ОПНИДЕММНЛ ЩБНКЧЖХНММНЛ ОСРХ. бШЯНЙЮЪ ЛСРЮАЕКЭМНЯРЭ пмй, ОПЕБШЬЮЧЫЮЪ ЛСРЮАЕКЭ- МНЯРЭ дмй Б IOOOOO ПЮГ, Х АШЯРПНЕ МЮЙНОКЕМХЕ ЛНКЕЙСКЪПМШУ ХГЛЕМЕМХИ ОНГБНКЪЧР БНЯЯРЮМЮБКХБЮРЭ УНД ЛНКЕЙСКЪПМНИ ЩБН╜КЧЖХХ пмй-ЦЕМНЛМШУ БХПСЯНБ Я ОНЛНЫЭЧ АНКЕЕ ЯНБЕПЬЕММНЦН ЛЕРНДЮ, Р. Е. ОПЪЛНЦН ЯПЮБМЕМХЪ ЯНБПЕЛЕММШУ БХПСЯНБ Х ХУ ОПЕДЙНБ. лЮРЕПХЮКНЛ ДКЪ ХЯЯКЕДНБЮМХИ ЯКСФЮР ЬРЮЛЛШ БХ╜ПСЯНБ, БШДЕКЕММШЕ Я ХМРЕПБЮКНЛ Б 5; 10; 20 КЕР Х АНКЕЕ Х ПЮГ╜ДЕКЕММШЕ ЩБНКЧЖХНММНИ ДХЯРЮМЖХЕИ Б 5000; 10 000 Х 20 000 ОН╜ЙНКЕМХИ Х T-. Д. (ОПХ ЯПЕДМЕЛ ФХГМЕММНЛ ЖХЙКЕ БХПСЯЮ, ПЮБМНЛ 8 >В). яПЮБМЕМХЕ ЩРХУ ЬРЮЛЛНБ ДЮЕР БНГЛНФМНЯРЭ ОПНЯКЕДХРЭ УНД ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ Х НОПЕДЕКХРЭ ЕЕ УЮПЮЙРЕП. рЮЙХЛ НАПЮГНЛ, Н ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ пмй-ЦЕМНЛМШУ БХПСЯНБ ЯС╜ДЪР МЕ ОН ЦНРНБШЛ ПЕГСКЭРЮРЮЛ ХЯРНПХХ ОПНЬКНЦН, Ю МЕОНЯПЕД╜ЯРБЕММН ОН ХГЛЕМЕМХЧ ЮПУЮХВМШУ ТНПЛ.
жЕМРПЮКЭМШЛ ОНКНФЕМХЕЛ МЕИРПЮКХЯРЯЙНИ РЕНПХХ ЪБКЪЕРЯЪ ЕЕ РПЕРХИ ОНЯРСКЮР, ЯНЦКЮЯМН ЙНРНПНЛС ЛСРЮЖХНММШЕ ГЮЛЕМШ, ОПХБНДЪЫХЕ Й ЛЕМЭЬХЛ МЮПСЬЕМХЪЛ ЯРПСЙРСПШ Х ТСМЙЖХХ ЛНКЕЙСКШ (ЙНМЯЕПБЮРХБМШЕ ГЮЛЕМШ), Б УНДЕ ЩБНКЧЖХХ ОПНХЯ╜УНДЪР ВЮЫЕ РЕУ, ЙНРНПШЕ БШГШБЮЧР АНКЕЕ ЯСЫЕЯРБЕММНЕ МЮПС╜ЬЕМХЕ ЯРПСЙРСПШ Х ТСМЙЖХХ ДЮММНИ ЛНКЕЙСКШ. пЕВЭ ХДЕР Н ОПЕХЛСЫЕЯРБЕММНЛ МЮЙНОКЕМХХ МЕИРПЮКЭМШУ ХКХ ОНВРХ МЕИ╜РПЮКЭМШУ ЛСРЮЖХИ, Р. Е. ЛСРЮЖХИ, ЯСЫЕЯРБЕММН МЕ БКХЪЧЫХУ МЮ ТЕМНРХО ХКХ ФЕ ДЮЧЫХУ ЯКЮАШИ НРПХЖЮРЕКЭМШИ ЩТТЕЙР.
пЮЯЯЛНРПХЛ ОПХЛЕП, ОНЙЮГШБЮЧЫХИ ТХЙЯЮЖХЧ МЕИРПЮКЭ╜МШУ ЛСРЮЖХИ Б БХПСЯМНЛ ЦЕМНЛЕ. б. J. Jonson Х ЯНЮБР. (1986) ОПНБЕКХ ЯЕЙБЕМХПНБЮМХЕ дмй-ЙНОХИ 26S PHK БХПСКЕМРМНЦН ЬРЮЛЛЮ БХПСЯЮ БЕМЕЯСЩКЭЯЙНЦН ЩМЖЕТЮКНЛХЕКХРЮ КНЬЮДЕИ рПХМХДЮД Х ОНКСВЕММНЦН ХГ МЕЦН ЮРРЕМСХПНБЮММНЦН ЬРЮЛЛЮ ря-83. й ЮРРЕМСЮЖХХ ОПХБЕКХ ДКХРЕКЭМНЕ ОЮЯЯХПНБЮМХЕ Х ЯЕ╜КЕЙЖХЪ БХПСЯЮ Б ЙСКЭРСПЮУ ЙКЕРНЙ. лНКЕЙСКЮ 26S PHK ЯНДЕП╜ФХР ЦЕМШ 3 ЯРПСЙРСПМШУ АЕКЙНБ БХПХНМЮ: ОНБЕПУМНЯРМШУ ЦКХ╜ЙНОПНРЕХДНБ е2 Х ElХ БМСРПЕММЕЦН АЕКЙЮ я. оНЙЮГЮМН, ВРН МХ ОНЯКЕДНБЮРЕКЭМНЯРЭ ЦЕМНБ, МХ ОПНРЪФЕММНЯРЭ ЛНКЕЙСКШ, МХ ОН╜КНФЕМХЕ НРЙПШРНИ ПЮЛЙХ ЯВХРШБЮМХЪ ОПХ ЮРРЕМСЮЖХХ МЕ ХГ╜ЛЕМЪКХЯЭ. юРРЕМСХПНБЮММШИ ЬРЮЛЛ ря-83 НРКХВЮКЯЪ НР БХПС╜КЕМРМНЦН ОПЕДЬЕЯРБЕММХЙЮ ЬРЮЛЛЮ рПХМХДЮД ОН 13 МСЙКЕН╜РХДМШЛ ГЮЛЕМЮЛ, ХГ. ЙНРНПШУ РНКЭЙН 5 БЕКХ Й ГЮЛЕМЮЛ ЮЛХМН╜ЙХЯКНР. бЯЕ 5 ЛСРЮЖХИ, БКХЪЧЫХУ МЮ ЮЛХМНЙХЯКНРМСЧ ОНЯКЕ╜ДНБЮРЕКЭМНЯРЭ, КНЙЮКХГНБЮКХЯЭ Б ЦЕМЕ ДКЪ АЕКЙЮ е2. щРНР ЦЕМ, Ю РЮЙФЕ МЕЙНДХПСЧЫЮЪ НАКЮЯРЭ С 5'-ЙНМЖЮ ЛНКЕЙСКШ 26S PHK ПЮЯЯЛЮРПХБЮЧРЯЪ ЙЮЙ БНГЛНФМШЕ ЦЕМЕРХВЕЯЙХЕ ДЕРЕПЛХМЮМРШ БХПСКЕМРМНЯРХ. б ТСМЙЖХНМЮКЭМНЛ ОКЮМЕ ЦКХЙНОПНРЕХД е2 БШ╜ГШБЮЕР НАПЮГНБЮМХЕ ОПНРЕЙРХБМШУ ЮМРХРЕК Х, ОН-БХДХЛНЛС,
НОПЕДЕКЪЕР ЯОЕЖХТХВМНЯРЭ ОПХ БГЮХЛНДЕИЯРБХХ БХПСЯЮ Я ЙКЕР╜ЙНИ. с БХПСЯНБ ЙНЛОКЕЙЯЮ БЕМЕЯСЩКЭЯЙНЦН ЩМЖЕТЮКНЛХЕКХРЮ КНЬЮДЕИ АЕКНЙ е2 УЮПЮЙРЕПХГСЕРЯЪ ГМЮВХРЕКЭМНИ ЮМРХЦЕММНИ ХГЛЕМВХБНЯРЭЧ. бЯЕ 7 ЛСРЮЖХИ, БШЪБКЕММШЕ Б ЦЕМЮУ АЕКЙНБ ДКЪ ElХ CЮРРЕМСХПНБЮММНЦН ЬРЮЛЛЮ ря-83, МНЯХКХ МЕИРПЮКЭ╜МШИ УЮПЮЙРЕП Х МЕ БКЕЙКХ ГЮ ЯНАНИ ЮЛХМНЙХЯКНРМШУ ГЮЛЕМ.
бНГЛНФМНЯРЭ БНГМХЙМНБЕМХЪ Х ТХЙЯЮЖХХ Б ЦЕМНЛЕ МЕИРПЮКЭ╜МШУ ЛСРЮЖХИ НОПЕДЕКЪЕРЯЪ ЯБНИЯРБЮЛХ ЦЕМЕРХВЕЯЙНЦН ЙНДЮ. мСЙКЕНРХДШ, ЯНЯРЮБКЪЧЫХЕ ЙНДНМШ, ХЛЕЧР МЕНДХМЮЙНБСЧ ЯЛШЯКНБСЧ МЮЦПСГЙС. оПНХЯУНФДЕМХЕ ЩРХУ ПЮГКХВХИ ЯБЪГЮМН Я ЩБНКЧЖХЕИ ЦЕМЕРХВЕЯЙНЦН ЙНДЮ. хГ 3 МСЙКЕНРХДНБ, ЯНЯРЮБКЪ╜ЧЫХУ ЙНДНМШ, МЮХАНКЭЬЮЪ ЯЛШЯКНБЮЪ МЮЦПСГЙЮ КНФХРЯЪ МЮ ОЕПБШИ Х БРНПНИ. рПЕРХИ МСЙКЕНРХД УЮПЮЙРЕПХГСЕРЯЪ МХГЙНИ ЯОЕЖХТХВМНЯРЭЧ. б ПЕГСКЭРЮРЕ ЩРНЦН ЯПНДЯРБН ЙНДНМНБ Б НРМН╜ЬЕМХХ ЮЛХМНЙХЯКНР ВЮЯРН НОПЕДЕКЪЕРЯЪ ДБСЛЪ ОЕПБШЛХ МСЙ╜КЕНРХДЮЛХ. рПЕРЭЪ ОНГХЖХЪ ЛНФЕР АШРЭ ОПЕДЯРЮБКЕМЮ НДМХЛ ХГ МЕЯЙНКЭЙХУ МСЙКЕНРХДНБ. яЛШЯК РПХОКЕРЮ Х ЕЦН ЯОНЯНАМНЯРЭ ЙНДХПНБЮРЭ ЯХМРЕГ НОПЕДЕКЕММНИ ЮЛХМНЙХЯКНРШ ОПХ ЩРНЛ МЕ ХГЛЕМЪЧРЯЪ. рЮЙХЛ НАПЮГНЛ, МСЙКЕНРХДМШЕ ГЮЛЕМШ Б РПЕРЭЕЛ ОНКНФЕМХХ ЙНДНМЮ ВЮЫЕ БЯЕЦН МЕИРПЮКЭМШ ХКХ, ХМЮВЕ, МЕИ╜РПЮКЭМШЕ ЛСРЮЖХХ НАШВМН ТХЙЯХПСЧРЯЪ ХЛЕММН Б ЩРНИ ОНГХЖХХ.
оПЕХЛСЫЕЯРБЕММНЕ МЮЙНОКЕМХЕ ЛСРЮЖХИ Б РПЕРЭЕЛ ОНКНФЕ╜МХХ ЙНДНМЮ УЮПЮЙРЕПМН ДКЪ БХПСЯМНЦН ЦЕМНЛЮ. N. Saitou (1987) НАНАЫХК ДЮММШЕ Н УЮПЮЙРЕПЕ Х КНЙЮКХГЮЖХХ МСЙКЕНРХДМШУ ГЮЛЕМ, НАМЮПСФЕММШУ Б ЦЕМЮУ ДКЪ ЦЕЛЮЦЦКЧРХМХМНБ HlХ м2, ЦЕМЮУ МЕИПЮЛХМХДЮГШ NlХ N2,Ю РЮЙФЕ ЦЕМЮУ МЕЯРПСЙРСПМНЦН АЕКЙЮ NSХ ЛЮРПХЙЯМНЦН АЕКЙЮ MБХПСЯЮ ЦПХООЮ ю ВЕКНБЕЙЮ. бЯЕЦН Б РПЕРЭЕЛ ОНКНФЕМХХ ЙНДНМЮ НРЛЕВЮКНЯЭ 268 ЛСРЮЖХИ, Ю Б ОЕПБНЛ Х БРНПНЛ ≈ РНКЭЙН 189.
лСРЮЖХХ Б РПЕРЭЕЛ ОНКНФЕМХХ ЙНДНМЮ ВЮЫЕ БЯЕЦН ЯХМНМХ╜ЛХВМШ, Р. Е. НМХ МЕ ОПХБНДЪР Й ГЮЛЕМЮЛ ЮЛХМНЙХЯКНРМШУ НЯРЮР╜ЙНБ Б ЛНКЕЙСКЕ АЕКЙЮ. J. R. Wiener Х W. й. Joklik (1988) ОПН╜БЕКХ ЯПЮБМЕМХЕ МСЙКЕНРХДМШУ ОНЯКЕДНБЮРЕКЭМНЯРЕИ ЦЕМНЛМНЦН ЯЕЦЛЕМРЮ л2ПЕНБХПСЯНБ ВЕКНБЕЙЮ РХОЮ 1; 2 Х 3,ПЮГНЬЕДЬХУ╜ЯЪ Б ЯБНЕИ ЩБНКЧЖХХ Х ПЮГКХВЮЧЫХУЯЪ ЯЕПНКНЦХВЕЯЙХ Х ОН МЕ╜ЙНРНПШЛ ДПСЦХЛ АХНКНЦХВЕЯЙХЛ ЯБНИЯРБЮЛ. яЕЦЛЕМР л2ЙНДХ╜ПСЕР ЯХМРЕГ ЙХЯКНЦН АЕКЙЮ μ/, УЮПЮЙРЕПХГСЧЫЕЦНЯЪ МХГЙХЛ ЯН╜ДЕПФЮМХЕЛ ЖХЯРХМЮ, ЦХЯРХДХМЮ Х ЛЕРХНМХМЮ Х АНЦЮРНЦН ОПН╜КХМНЛ. нЙНКН 27% ЮЛХМНЙХЯКНРМШУ НЯРЮРЙНБ МЮУНДЪРЯЪ Б Ю-ЯОХПЮКЭМНИ ЙНМТХЦСПЮЖХХ. б ПЕГСКЭРЮРЕ ОПНРЕНКХРХВЕЯЙНЦН ПЮЯЫЕОКЕМХЪ АЕКЙЮ μl,ЙНРНПНЕ ОПНХЯУНДХР ЛЕФДС ЮЛХМНЙХЯ╜КНРМШЛХ НЯРЮРЙЮЛХ, МЮУНДЪЫХЛХЯЪ Б ОНГХЖХЪУ 42 Х 43, НАПЮ╜ГСЕРЯЪ АЕКНЙ μ7C, ЙНРНПШИ ЪБКЪЕРЯЪ НЯМНБМШЛ ЯРПСЙРСПМШЛ ЙНЛОНМЕМРНЛ БХПХНМЮ. яЕЦЛЕМРШ л2ЯЕПНРХОНБ 1Х 2, 1Х 3Х 2Х 3ХЛЕКХ ЯННРБЕРЯРБЕММН 15%, 23% Х 23% НАЫХУ НЯМНБЮ╜МХИ. б ОПНРХБНОНКНФМНЯРЭ ЩРНЛС ОПНДСЙР ЦЕМЮ л2,АЕКНЙ μ/, УЮПЮЙРЕПХГНБЮКЯЪ БШЯНЙНИ ЯРЕОЕМЭЧ ЦНЛНКНЦХХ ≈ 97%. йНМ╜
ЯЕПБЮРХГЛ АЕКЙЮ μ/ ДНЯРХЦЮКЯЪ РЕЛ, ВРН АНКЭЬХМЯРБН ЛСРЮЖХИ PHK ТХЙЯХПНБЮКНЯЭ Б РПЕРЭЕЛ ОНКНФЕМХХ ЙНДНМЮ Х МЕ ОПХБН╜ДХКН Й ЮЛХМНЙХЯКНРМШЛ ГЮЛЕМЮЛ. оПХ ЯПЮБМЕМХХ ЦЕМНЛМНЦН ЯЕЦЛЕМРЮ л2ЯЕПНРХОЮ 1Х 3Б РПЕРЭЕЛ ОНКНФЕМХХ ЙНДНМЮ АШКН НАМЮПСФЕМН 87,6%, ЯЕПНРХОНБ 1Х 2≈86,4% Х ЯЕПНРХОНБ 2Х 3≈86,6% БЯЕУ БШЪБКЕММШУ ЛСРЮЖХИ. нРМНЯХРЕКЭМЮЪ ВЮЯРНРЮ ЛСРЮЖХИ Б ОЕПБНЛ ОНКНФЕМХХ ЙНДНМЮ АШКЮ 9,6%; 11,8% Х 11,2%, Ю БН БРНПНЛ≈2,8%; 1,8% Х 2,2% ЯННРБЕРЯРБЕММН ДКЪ ЯЕПНРХОНБ 1Х г, 1Х 2 Х 2 Х 3.рНКЭЙН 18 ХГ 324 ЛСРЮЖХИ, БШЪБКЕММШУ ОПХ ЯПЮБМЕМХХ ЯЕПНРХОНБ 1Х 3,ОПХБНДХКХ Й ЮЛХ╜МНЙХЯКНРМШЛ ГЮЛЕМЮЛ. дКЪ ЯЕПНРХОНБ 1Х 2, 2Х 3ЩРНР ОНЙЮ╜ГЮРЕКЭ ЯНЯРЮБКЪК ЯННРБЕРЯРБЕММН 20 ХГ 501 Х 23 ХГ 507.
оНЛХЛН ЯБНИЯРБ ЦЕМЕРХВЕЯЙНЦН ЙНДЮ, МЕИРПЮКЭМШИ УЮПЮЙРЕП ЛСРЮЖХИ НОПЕДЕКЪЕРЯЪ НЯНАЕММНЯРЪЛХ ЯРПСЙРСПШ АЕКЙНБНИ ЛН╜КЕЙСКШ. лНКЕЙСКЮ АЕКЙЮ ЯНЯРНХР ХГ МЕАНКЭЬНЦН ТСМЙЖХНМЮКЭ╜МН ЮЙРХБМНЦН ЖЕМРПЮ Х ГМЮВХРЕКЭМНИ ОН ОПНРЪФЕММНЯРХ ╚ОЮЯ╜ЯХБМНИ╩ ВЮЯРХ. тХЙЯЮЖХЪ ЛСРЮЖХИ Б ╚ОЮЯЯХБМНИ╩ ВЮЯРХ МЕ ХГ╜ЛЕМЪЕР Х ТСМЙЖХЧ АЕКЙЮ Х МЕ БКХЪЕР МЮ ТЕМНРХО. йПНЛЕ РНЦН, ЙНППЕКЪЖХЪ ЛЕФДС ОЕПБХВМНИ ЯРПСЙРСПНИ ЛНКЕЙСКШ АЕКЙЮ Х ЯРПСЙРСПЮЛХ БШЯЬЕЦН ОНПЪДЙЮ, ЙНРНПШЕ НОПЕДЕКЪЧР ТСМЙЖХН╜МЮКЭМСЧ ЮЙРХБМНЯРЭ ЛНКЕЙСКШ, МНЯХР БШПНФДЕММШИ УЮПЮЙРЕП. оНЩРНЛС ХГЛЕМЕМХЕ ЮЛХМНЙХЯКНРМНИ ОНЯКЕДНБЮРЕКЭМНЯРХ, МЮ╜ЯРСОЮЧЫЕЕ Б ПЕГСКЭРЮРЕ ЛСРЮЖХИ, ЛНФЕР МЕ БКХЪРЭ МЮ ЯРПСЙ╜РСПШ БШЯЬЕЦН ОНПЪДЙЮ Х МЮ ТСМЙЖХЧ АЕКЙЮ. лСРЮЖХХ МНЯЪР МЕИРПЮКЭМШИ УЮПЮЙРЕП.
йКЮЯЯХВЕЯЙХЛ ОПХЛЕПНЛ ЯНУПЮМЕМХЪ ТСМЙЖХНМЮКЭМНИ ЮЙ╜РХБМНЯРХ БХПСЯМНЦН АЕКЙЮ Б СЯКНБХЪУ МЮЙНОКЕМХЪ МЕИРПЮКЭМШУ ЛСРЮЖХИ ЯКСФХР ЯКЕДСЧЫЕЕ. J. Blok Х G. л. Air (1982) ЯПЮБ╜МХКХ ОЕПБХВМСЧ ЯРПСЙРСПС СВЮЯРЙНБ МЕИПЮЛХМХДЮГШ 8 ОНДРХ╜ОНБ БХПСЯЮ ЦПХООЮ ю. лНКЕЙСКЮ МЕИПЮЛХМХДЮГШ ХЛЕЕР РПЕУ╜ЛЕПМСЧ ЯРПСЙРСПС: МНФЙЮ, ЦНКНБЙЮ Х РПЮМЯЛЕЛАПЮММШИ ЯЕЦЛЕМР Я ЦХДПНТНАМШЛ СВЮЯРЙНЛ МЮ NH2-κoH∏e. C ЦНКНБЙНИ ЯБЪГЮМЮ ЮМРХЦЕММЮЪ Х ТЕПЛЕМРЮРХБМЮЪ ЮЙРХБМНЯРЭ АЕКЙЮ. цХДПНТНАМШИ СВЮЯРНЙ НАЕЯОЕВХБЮЕР ОПХЙПЕОКЕМХЕ ЛНКЕЙСКШ Й КХОХДМНЛС АХЯКНЧ БХПХНМЮ. мНФЙЮ ЯНЕДХМЪЕР ЩРХ СВЮЯРЙХ ЛНКЕЙСКШ. яРЮ╜АХКХГЮЖХЪ МНФЙХ НАЕЯОЕВХБЮЕРЯЪ СЦКЕБНДЮЛХ Х ДХЯСКЭТХДМШ╜ЛХ ЯБЪГЪЛХ. яПЮБМЕМХЕ ОЕПБХВМНИ ЯРПСЙРСПШ РПЮМЯЛЕЛАПЮММН╜ЦН ЯЕЦЛЕМРЮ Х МНФЙХ МЕИПЮЛХМХДЮГШ ОНЙЮГЮКН, ВРН ЩРХ СВЮЯР╜ЙХ НРКХВЮЧРЯЪ ОН ЮЛХМНЙХЯКНРМНИ ОНЯКЕДНБЮРЕКЭМНЯРХ. мЕ╜ЯЛНРПЪ МЮ МХГЙСЧ ЦНЛНКНЦХЧ, НАЫХИ УЮПЮЙРЕП ОНКХОЕОРХДМШУ ЖЕОЕИ Х ХУ ЯБНИЯРБЮ ЯНУПЮМЪКХЯЭ. рПЮМЯЛЕЛАПЮММШИ ЯЕЦЛЕМР С БЯЕУ ОНДРХОНБ МЕИПЮЛХМХДЮГШ ЯНДЕПФЮК БШЯНЙХИ ОПНЖЕМР ЦХДПНТНАМШУ ЮЛХМНЙХЯКНР. б ЯРПСЙРСПЕ МНФЙХ НАМЮПСФЕМШ НЯРЮРЙХ ЖХЯРЕХМЮ, ЯОНЯНАМШЕ ТНПЛХПНБЮРЭ ДХЯСКЭТХДМШЕ ЯБЪ╜ГХ, Х ЯЮИРШ ЦКХЙНГХКХПНБЮМХЪ. дХЯСКЭТХДМШЕ ЯБЪГХ Х ОПХЯН╜ЕДХМЕМХЕ СЦКЕБНДНБ НАЕЯОЕВХБЮЧР ЯРЮАХКХГЮЖХЧ РПЕУЛЕПМНИ ^ЯРПСЙРСПШ МНФЙХ. оНЙЮГЮМН, ВРН БШПЮФЕММЮЪ ЛСРЮЖХНММЮЪ ОЕ╜
ПЕЯРПНИЙЮ ОЕПБХВМНИ ЯРПСЙРСПШ МЕ БКХЪКЮ МЮ РПЕРХВМСЧ ЯРПСЙ╜РСПС ХГСВЕММШУ СВЮЯРЙНБ ЛНКЕЙСКШ МЕИПЮЛХМХДЮГШ Х МЕ ХГ╜ЛЕМХКЮ ХУ ТСМЙЖХЧ. мЮЙНОХБЬХЕЯЪ ЛСРЮЖХХ МНЯХКХ МЕИРПЮКЭ╜МШИ УЮПЮЙРЕП'.
рЮЙХЛ НАПЮГНЛ, МЕИРПЮКЭМШЕ ЛСРЮЖХХ ПЮГДЕКЪЧРЯЪ МЮ ДБЮ РХОЮ. лСРЮЖХХ ОЕПБНЦН РХОЮ ТХЙЯХПСЧРЯЪ Б ЦЕМНЛЕ, МН МЕ БКХЪ╜ЧР МЮ ЮЛХМНЙХЯКНРМСЧ ОНЯКЕДНБЮРЕКЭМНЯРЭ ЛНКЕЙСКШ АЕКЙЮ, Ю БРНПНЦН ≈ХГЛЕМЪЧР МСЙКЕНРХДМСЧ ОНЯКЕДНБЮРЕКЭМНЯРЭ Х ОНЯКЕДНБЮРЕКЭМНЯРЭ ЮЛХМНЙХЯКНРМШУ НЯРЮРЙНБ Б ЛНКЕЙСКЕ АЕК╜ЙЮ, МН МЕ МЮПСЬЮЧР ЕЦН ТСМЙЖХЧ. йЮЙ ЯКЕДСЕР ХГ ОПХБЕДЕММШУ ОПХЛЕПНБ, НАЮ РХОЮ МЕИРПЮКЭМШУ ЛСРЮЖХИ БЯРПЕВЮЧРЯЪ С БХ╜ПСЯНБ. лНКЕЙСКЪПМЮЪ ЩБНКЧЖХЪ БХПСЯНБ ХДЕР ОН ОСРХ ОПЕХЛС╜ЫЕЯРБЕММНЦН МЮЙНОКЕМХЪ ЩРХУ ЛСРЮЖХИ. б ПЕГСКЭРЮРЕ БХПСЯ╜МШИ ЦЕМНЛ ЯРЮМНБХРЯЪ ЦНРНБШЛ Й БГПШБС, ЯЙЮВЙС, ПЕГЙНЛС ОПЕ╜НАПЮГНБЮМХЧ. щРН ЯНГДЮЕР БНГЛНФМНЯРЭ ДКЪ ОНЪБКЕМХЪ МНБШУ ТНПЛ ХКХ ОПХБНДХР БХПСЯ Й ЦХАЕКХ.
аХНКНЦХВЕЯЙХИ ЯЛШЯК МЕИРПЮКХЯРЯЙНИ РЕНПХХ ГЮЙКЧВЮЕР╜ЯЪ Б РНЛ, ВРН Б НЯМНБЕ ЩБНКЧЖХНММШУ ХГЛЕМЕМХИ МЮ ЛНКЕЙС╜КЪПМНЛ СПНБМЕ, Р. Е. ОЕПЕЯРПНИЙХ ЯЮЛНЦН ЦЕМЕРХВЕЯЙНЦН ЛЮРЕПХЮКЮ, КЕФЮР МЕ БНГДЕИЯРБХЪ БМЕЬМЕИ ЯПЕДШ, Ю МЕГЮ╜БХЯХЛШЕ НР МХУ БМСРПЕММХЕ ЦЕМЕРХВЕЯЙХЕ ОПНЖЕЯЯШ. хГЛЕМЕ╜МХЕ дмй Х PHK Б НЯМНБМНЛ ОПНХЯУНДХР ОСРЕЛ ТХЙЯЮЖХХ МЕИРПЮКЭМШУ Х ОНВРХ МЕИРПЮКЭМШУ ЛСРЮЖХИ, МЮЙНОКЕМХЕ ЙНРНПШУ МЕ ЙНМРПНКХПСЕРЯЪ НРАНПНЛ Х НЯСЫЕЯРБКЪЕРЯЪ АЕГ ЕЦН БЛЕЬЮРЕКЭЯРБЮ. яЙНПНЯРЭ ГЮЙПЕОКЕМХЪ ЩРХУ ЛСРЮЖХИ НОПЕДЕКЪЕРЯЪ ЦКЮБМШЛ НАПЮГНЛ ВЮЯРНРНИ ЛСРХПНБЮМХЪ. яХМ╜РЕРХВЕЯЙЮЪ РЕНПХЪ ЩБНКЧЖХХ ОНДДЕПФХБЮЕР ОПНРХБНОНКНФ╜МНЕ ЛМЕМХЕ Н РНЛ, ВРН ТХЙЯХПНБЮММШЕ ЛСРЮЖХНММШЕ ХГЛЕМЕ╜МХЪ ЮДЮОРХБМШ Х НАСЯКНБКЕМШ ОНКНФХРЕКЭМШЛ ДЮПБХМНБ╜ЯЙХЛ НРАНПНЛ. оПЕХЛСЫЕЯРБЕММН ГЮЙПЕОКЪЧРЯЪ ЛСРЮЖХХ, ХЛЕЧЫХЕ ТЕМНРХОХВЕЯЙНЕ ОПНЪБКЕМХЕ Х ДЮЧЫХЕ ОНКНФХ╜РЕКЭМШИ ЩТТЕЙР. тЮЙР МЮЙНОКЕМХЪ МЕИРПЮКЭМШУ Х ОНВРХ МЕИРПЮКЭМШУ ЛСРЮЖХИ ОНДРБЕПФДЕМ НАЬХПМШЛ ТЮЙРХВЕЯЙХЛ ЛЮРЕПХЮКНЛ, ОНКСВЕММШЛ ОПХ ЯПЮБМХРЕКЭМНЛ ХГСВЕМХХ ОЕП╜БХВМНИ ЯРПСЙРСПШ ЦЕМНЛЮ Х АЕКЙНБ ПЮГКХВМШУ НПЦЮМХГЛНБ. дЮММШЕ Н БХПСЯЮУ РЮЙФЕ ЯБХДЕРЕКЭЯРБСЧР Б ОНКЭГС МЕИРПЮ╜КХЯРЯЙНИ РЕНПХХ.
мЕИРПЮКЭМШЕ Х ОНВРХ МЕИРПЮКЭМШЕ ЛСРЮЖХХ ЮЙЙСЛСКХ╜ПСЧРЯЪ Б ЦЕМНЛЕ Х АЕКЙЮУ, МН ДН ОНПШ ЯСЫЕЯРБЕММН МЕ БКХЪ-. ЧР МЮ БШФХБЮЕЛНЯРЭ Х БНЯОПНХГБЕДЕМХЕ БХДЮ. оПХ ХГЛЕМЕМХХ СЯКНБХИ ЯПЕДШ ОНКНФЕМХЕ ЛНФЕР ХГЛЕМХРЭЯЪ, ЛСРЮЖХХ, ЙНРН╜ПШЕ АШКХ МЕИРПЮКЭМШЛХ, ЯРЮМНБЪРЯЪ ЯЕКЕЙРХБМН ЖЕММШЛХ Х ОПЕБПЮЫЮЧРЯЪ Б ХЯУНДМШИ ЛЮРЕПХЮК ДКЪ ЮДЮОРХБМНИ ЩБН╜КЧЖХХ. щРНР ЛЮРЕПХЮК ОПЕДЯСЫЕЯРБСЕР Б ЦЕМНЛЕ Х СБЕКХВХ╜БЮЕР РЕЛ ЯЮЛШЛ ЩБНКЧЖХНММШИ ОНРЕМЖХЮК НПЦЮМХГЛНБ. оН- БХДХЛНЛС, РЮЙНИ ПЕГЕПБ ЮЙРХБМН ХЯОНКЭГСЕРЯЪ Б ОЕПХНДШ ОНБШЬЕМХЪ ЩБНКЧЖХНММНИ ЮЙРХБМНЯРХ, БН БПЕЛЪ ЙНРНПШУ,
ЯНЦКЮЯМН ЙНМЖЕОЖХХ ОПЕПШБХЯРНЦН ПЮБМНБЕЯХЪ, ОПНХЯУНДХР БХДННАПЮГНБЮМХЕ.
щБНКЧЖХНММНЕ ГМЮВЕМХЕ МЕИРПЮКЭМШУ Х ОНВРХ МЕИРПЮКЭ╜МШУ ЛСРЮЖХИ ГЮЙКЧВЮЕРЯЪ РЮЙФЕ Б РНЛ, ВРН НМХ ЯНГДЮЧР БНГЛНФМНЯРЭ ДКЪ НЯСЫЕЯРБКЕМХЪ ПЕГЙХУ ЩБНКЧЖХНММШУ ЯДБХ╜ЦНБ. Й РЮЙНЛС ЯДБХЦС ЛНФЕР ОПХБЕЯРХ ДЮФЕ НДМЮ ╚СДЮВМЮЪ╩ ЛСРЮЖХЪ, ЙНРНПЮЪ ОПНХЯУНДХР Б ЦЕМНЛЕ, МЮЯШЫЕММНЛ ЛНКВЮ╜ЫХЛХ Х МЕ ОПНЪБКЪЧЫХЛХ ЯЕАЪ ЛСРЮЖХЪЛХ, Х ЯНГДЮЧЫЮЪ ГЮ ЯВЕР ЩРНЦН МНБСЧ ЙНЛАХМЮЖХЧ ЦЕМЕРХВЕЯЙХУ ТЮЙРНПНБ. нАПЮГСЕРЯЪ ЦЕМ Я МНБШЛХ ЯБНИЯРБЮЛХ. пЕГЙН ХГЛЕМЪЕРЯЪ ТЕ╜МНРХО. оПНЖЕЯЯ МНЯХР УЮПЮЙРЕП ТЮГНБНЦН ОЕПЕУНДЮ Х ОПНХЯ╜УНДХР ЯЙЮВЙНЛ, ЯЮКЭРЮЖХНММН. бНГЛНФЕМ ЙЮЙ ОНКНФХРЕКЭМШИ, РЮЙ Х НРПХЖЮРЕКЭМШИ ЩТТЕЙР. мЮ ОНЯКЕДМЕЛ ЛШ НЯРЮМНБХЛ╜ЯЪ МХФЕ ОПХ НАЯСФДЕМХХ ОПНАКЕЛШ ЛЮЯЯНБШУ БШЛХПЮМХИ. йЮЙ МЮЙНОКЕМХЕ МЕИРПЮКЭМШУ Х ОНВРХ МЕИРПЮКЭМШУ ЛСРЮЖХИ, РЮЙ Х ОЕПЕУНД ЩРНЦН ЛЮРЕПХЮКЮ Б МНБНЕ ЙЮВЕЯРБН НЯСЫЕЯР╜БКЪЧРЯЪ АЕГ СВЮЯРХЪ НРАНПЮ. оНЩРНЛС ОНЪБХБЬХЕЯЪ ОПХГМЮ╜ЙХ ЛНЦСР МЕ ХЛЕРЭ ЮДЮОРХБМНЦН ГМЮВЕМХЪ. оПХЛЕПНЛ РЮЙХУ, ОПХНАПЕРЕМХИ ЪБКЪЕРЯЪ АНКЭЬХМЯРБН ЯКСВЮЕБ ОНКХЛНПТХГЛЮ АЕКЙНБ, ЬХПНЙН ПЮЯОПНЯРПЮМЕММНЦН С БШЯЬХУ ЩСЙЮПХНРНБ. оПЕ╜НАКЮДЮЧЫЮЪ ВЮЯРЭ ОНКХЛНПТМШУ ЮККЕКЕИ, ХЛЕЧЫХУЯЪ С ЙЮ╜ЙНЦН-КХАН БХДЮ, ОНДДЕПФХБЮЕРЯЪ ГЮ ЯВЕР ЛСРЮЖХНММНЦН ОПНЖЕЯ╜ЯЮ Х ЯКСВЮИМНИ ЩКХЛХМЮЖХХ.
нЯНАЕММНЯРЭ ПЮЯЯЛНРПЕММНЦН ЩБНКЧЖХНММНЦН ОСРХ ГЮЙКЧВЮ╜ЕРЯЪ Б РНЛ, ВРН НРАНП МЕ ЯНГДЮЕР МНБШУ ТНПЛ. нМ ОПНХГБНДХР КХЬЭ ХУ ЮОПНАЮЖХЧ Х АПЮЙНБЙС. мНБШЕ ТНПЛШ, ОПНЬЕДЬХЕ ЩРС ОПНБЕПЙС, ДЮКЕЕ ╚ДНЬКХТНБЭЦБЮЧРЯЪ╩ НРАНПНЛ. б ОПНРХБНОН╜КНФМНЯРЭ ЩРНЛС ЯХМРЕРХВЕЯЙЮЪ РЕНПХЪ ЩБНКЧЖХХ НРБНДХР НР╜АНПС БЕДСЫСЧ ПНКЭ Б ТНПЛННАПЮГНБЮРЕКЭМШУ ОПНЖЕЯЯЮУ. яКЕ╜ДСЕР НРЛЕРХРЭ, ВРН СЙЮГЮММШЕ ЩБНКЧЖХНММШЕ ОСРХ МЕ ХЯЙКЧ╜ВЮЧР, Ю ДНОНКМЪЧР ДПСЦ ДПСЦЮ, НАЕЯОЕВХБЮЪ ЩБНКЧЖХНММНЕ ПЮГБХРХЕ АХНЯТЕПШ.
юМЮКХГ ХГЛЕМВХБНЯРХ ОПХПНДМШУ ОНОСКЪЖХИ БХПСЯНБ Х ПЮЯ╜ЯЛНРПЕМХЕ ДБХФЕМХЪ ГЮАНКЕБЮЕЛНЯРХ ОНЙЮГШБЮЧР, ВРН НДМХЛ ХГ ЯОНЯНАНБ ЩБНКЧЖХХ БХПСЯНБ ЪБКЪЕРЯЪ БМЕГЮОМНЕ ЯЙЮВЙННА╜ПЮГМНЕ ТНПЛННАПЮГНБЮМХЕ. гЮ ОНЯКЕДМХЕ 2QКЕР НАМЮПСФЕМШ БХПСЯШ, БШГШБЮЧЫХЕ МНБШЕ АНКЕГМХ ВЕКНБЕЙЮ Х ФХБНРМШУ. щРН ЩРХНКНЦХВЕЯЙХЕ ЮЦЕМРШ яохд бхв-1 Х бхв-2, БХПСЯШ ХЛЛС╜МНДЕТХЖХРЮ НАЕГЭЪМ, ЙНЬЕЙ Х ЙПСОМНЦН ПНЦЮРНЦН ЯЙНРЮ, БШГШ╜БЮЧЫХЕ ХЛЛСМНДЕТХЖХРМШЕ ЯНЯРНЪМХЪ С ЯБНХУ ╚УНГЪЕБ╩; ЩМРЕ╜ПНБХПСЯ 70,ЯКСФЮЫХИ ОПХВХМНИ ЦЕЛНППЮЦХВЕЯЙНЦН ЙЕПЮРНЙНМЗ╜ЧМЙРХБХРЮ ВЕКНБЕЙЮ; ОЮПБНБХПСЯ, БШГШБЮЧЫХИ ЛХНЙЮПДХР Х ДХЮПЕЧ С ЫЕМЙНБ; ЩРХНКНЦХВЕЯЙХИ ЮЦЕМР с-ЙКЕРНВМНЦН КЕИЙНГЮ ВЕКНБЕЙЮ Х РПНОХВЕЯЙНЦН ЯОЮЯРХВЕЯЙНЦН ОЮПЮОЮПЕГЮ; БХПСЯ T- ЙКЕРНВМНЦН КЕИЙНГЮ ВЕКНБЕЙЮ РХОЮ 1; ЩМРЕПХРМШЕ БХПСЯШ ВЕКН╜БЕЙЮ (ПНРЮ-, ЙЮКЭЖХ-, ЮЯРПНБХПСЯШ, мНПТНКЙ-ОНДНАМШЕ ЮЦЕМРШ Х Р. Д.). б АНКЭЬХМЯРБЕ ЯКСВЮЕБ ПЕВЭ ХДЕР МЕ Н ОНЪБКЕМХХ МН╜
БШУ БХПСЯНБ, Ю Н ПЮЯОНГМЮБЮМХХ НЯРЮБЬХУЯЪ МЕХГБЕЯРМШЛХ ЮЦЕМРНБ Х БШГШБЮЕЛШУ ХЛХ ГЮАНКЕБЮМХИ. мЮПЪДС Я ЩРХЛ ХГ╜БЕЯРМШ ОПХЛЕПШ НАПЮГНБЮМХЪ МНБШУ ТНПЛ. мЮХАНКЕЕ ДНЯРН╜БЕПМШЛХ ХГ МХУ ЪБКЪЧРЯЪ БНГМХЙМНБЕМХЕ бхв-1, ОЮПБНБХПСЯЮ ЛХНЙЮПДХРЮ Х ДХЮПЕХ ЫЕМЙНБ Х ЩМРЕПНБХПСЯЮ 70.щРХ БХПСЯШ ОНЪБХКХЯЭ БМЕГЮОМН Х АШЯРПН ПЮЯОПНЯРПЮМХКХЯЭ ЯПЕДХ МЕХЛ╜ЛСММШУ ╚УНГЪЕБ╩. хУ МЮЬЕЯРБХЕ ОПХБЕКН Й ЩОХДЕЛХЪЛ МЕХГ╜БЕЯРМШУ ПЮМЕЕ АНКЕГМЕИ. нЖЕМХБЮЪ ПЮГЛЕП ОПНХГНЬЕДЬХУ ЩБН╜КЧЖХНММШУ ЯДБХЦНБ, ЯКЕДСЕР НРЛЕРХРЭ, ВРН Б ЙЮФДНЛ ЯКСВЮЕ ДЕКН НЦПЮМХВХКНЯЭ ОНОНКМЕМХЕЛ СФЕ ХГБЕЯРМШУ ПНДНБ Х ЯЕ╜ЛЕИЯРБ (ПНДНБ ParvovirusХ Enterovirus,ЯЕЛЕИЯРБЮ Retroviri- dae)МНБШЛХ ОПЕДЯРЮБХРЕКЪЛХ. нАПЮГНБЮКХЯЭ МНБШЕ БХДШ.
бМЕГЮОМНЕ ЯЙЮВЙННАПЮГМНЕ ОНЪБКЕМХЕ ХГЛЕМЕММШУ ЬРЮЛЛНБ СФЕ ХГБЕЯРМШУ БХПСЯНБ ОПНХЯУНДХР АНКЕЕ ВЮЯРН Х ЪБКЪЕРЯЪ УЮ╜ПЮЙРЕПМНИ НЯНАЕММНЯРЭЧ БМСРПХБХДНБНИ ЛХЙПНЩБНКЧЖХХ PHK- ЦЕМНЛОШУ БХПСЯНБ. рЮЙХЕ ЬРЮЛЛШ ХЦПЮЧР АНКЭЬСЧ ПНКЭ Б ЩОХДЕЛХНКНЦХХ Х ЩОХГННРНКНЦХХ ЦПХООЮ, ЪЫСПЮ Х ЛМНЦХУ ДПС╜ЦХУ ХМТЕЙЖХИ. мНБШИ ЬРЮЛЛ БХПСЯЮ ЦПХООЮ ю ОРХЖ, МЮОПХЛЕП, ОПЕНДНКЕК ЛЕФБХДНБНИ АЮПЭЕП Х БШГБЮК Б 1979≈1980 ЦЦ. МЮ юРКЮМРХВЕЯЙНЛ ОНАЕПЕФЭЕ яью НОСЯРНЬХРЕКЭМСЧ ЩОХГННРХЧ ЦПХООЮ ЯПЕДХ РЧКЕМЕИ. щРН ОЕПБШИ Х ОНЙЮ ЕДХМЯРБЕММШИ ХГ╜БЕЯРМШИ ЯКСВЮИ ОПЪЛНЦН ОЕПЕУНДЮ ДНЯРЮРНВМН ЯОЕЖХЮКХГХПН╜БЮММНЦН БХПСЯЮ ЦПХООЮ ю ОРХЖ Й ЛКЕЙНОХРЮЧЫХЛ. бНГЛНФМН, ВРН ПЮЯЯЛНРПЕММШЕ ЩБНКЧЖХНММШЕ ЪБКЕМХЪ Б ПЪДЕ ЯКСВЮЕБ НАСЯКНБКЕМШ ЛСРЮЖХЪЛХ, ЙНРНПШЕ ╚ОЮДЮЧР МЮ ОНДЦНРНБКЕММСЧ ОНВБС╩, Р. Е. БНГМХЙЮЧР Б ЦЕМНЛЕ, МЮЯШЫЕММНЛ МЕИРПЮКЭМШЛХ ЛСРЮЖХЪЛХ Х ЦНРНБНЛ Й ОЕПЕЯРПНИЙЕ.
оНЛХЛН ЩБНКЧЖХНММНИ ПНКХ, МЕИРПЮКЭМШЕ ЛСРЮЖХХ БШОНК╜МЪЧР С пмй-ЦЕМНЛМШУ БХПСЯНБ ЕЫЕ НДМС ВПЕГБШВЮИМН ЯБНЕ╜НАПЮГМСЧ ТСМЙЖХЧ. щРН ГЮЫХРЮ НР ТНПЯХПНБЮММНИ ЩБНКЧЖХХ. аКЮЦНДЮПЪ БШЯНЙНИ ЛСРЮАЕКЭМНЯРХ ЦЕМЕРХВЕЯЙНЦН ЛЮРЕПХЮКЮ ЛНКЕЙСКЪПМЮЪ ЩБНКЧЖХЪ пмй-ЦЕМНЛМШУ БХПСЯНБ ХДЕР ВПЕГБШ╜ВЮИМН АШЯРПН. нАНАЫЮЪ ДЮММШЕ Н БХПСЯЮУ ЦПХООЮ ю, б Х я, ЪЫСПЮ, ПЕНБХПСЯЮУ, бхв Х ДП., ЛНФМН ЯЙЮГЮРЭ, ВРН ЯЙНПНЯРЭ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ БХПСЯМНЦН ЦЕМНЛЮ ЯНЯРЮБКЪЕР 2 X XlO-2≈5?ιlн-5 ГЮЛЕМ МЮ СВЮЯРНЙ Б ЦНД. оНЙЮГЮРЕКХ ЙНКЕА╜КЧРЯЪ НР БХПСЯЮ Й БХПСЯС Х НР ЦЕМЮ Й ЦЕМС. бЮФМШЕ Б ТСМЙ╜ЖХНМЮКЭМНЛ НРМНЬЕМХХ АЕКЙХ Х ЙНДХПСЧЫХЕ ХУ СВЮЯРЙХ ЦЕМН╜ЛЮ ХГЛЕМЪЧРЯЪ ЛЕДКЕММЕЕ, ВЕЛ ЛЕМЕЕ БЮФМШЕ. мЮОПХЛЕП, ОПХ ЯПЮБМЕМХХ ЛЮРПХВМНЦН АЕКЙЮ ХГНКЪРНБ БХПСЯЮ ЦПХООЮ A (H3N2) 1972 Х 1979 ЦЦ. НАМЮПСФЕМШ БЯЕЦН ДБЕ ЮЛХМНЙХЯКНРМШЕ ГЮЛЕ╜МШ. лЮРПХВМШИ АЕКНЙ МЕЯЕР ГМЮВХРЕКЭМСЧ ТСМЙЖХНМЮКЭМСЧ МЮЦПСГЙС: НАПЮГСЕР ДНЛЕМ МЮ БМСРПЕММЕИ ОНБЕПУМНЯРХ ЖХРН╜ОКЮГЛЮРХВЕЯЙНИ ЛЕЛАПЮМШ, ЦДЕ ОПНХЯУНДХР НАЗЕДХМЕМХЕ ЯЕЦ╜ЛЕМРНБ ЦЕМНЛЮ ОПХ ЯАНПЙЕ БХПХНМНБ, ЯОЕЖХТХВЕЯЙХ ПЮЯОНГМЮЕР БХПСЯМШЕ ЦКХЙНОПНРЕХДШ Х НАЕЯОЕВХБЮЕР ТНПЛХПНБЮМХЕ Х ЯРЮ╜АХКЭМНЯРЭ НАНКНВЙХ ОНД КХОХДМШЛ АХЯКНЕЛ БХПХНМЮ. оН-БХДХ╜
ЛНЛС, Б ЮЛХМНЙХЯКНРМНИ ОНЯКЕДНБЮРЕКЭМНЯРХ ЛНКЕЙСКШ ЛЮР╜ПХВМНЦН АЕКЙЮ ТХЙЯХПНБЮКХЯЭ КХЬЭ РЕ ГЮЛЕМШ, ЙНРНПШЕ МЕ МЮПСЬЮКХ ЕЦН ТСМЙЖХЧ. б ОПНРХБНОНКНФМНЯРЭ ЩРНЛС Б ЛНКЕ╜ЙСКЕ КЕЦЙНИ ЖЕОХ ЦЕЛЮЦЦКЧРХМХМЮ БХПСЯЮ ЦПХООЮ A (H3N2) ЕФЕЦНДМН ГЮЛЕМЪКНЯЭ 0,24≈0,57%, Ю Б РЪФЕКНИ ≈ 0,2≈0,91% ЮЛХМНЙХЯКНРМШУ НЯРЮРЙНБ. нЯРЮЕРЯЪ МЕЪЯМШЛ, ОПНРЕЙЮЕР КХ ЛНКЕЙСКЪПМЮЪ ЩБНКЧЖХЪ пмй-ЦЕМНЛМШУ БХПСЯНБ ПЮБМНЛЕПМН БН БПЕЛЕМХ. дЮММШЕ НА ЩРНЛ ОПНРХБНПЕВХБШ. б ОНКЭГС ОНЯРН╜ЪММНИ ЯЙНПНЯРХ ЩБНКЧЖХХ ЯБХДЕРЕКЭЯРБСЕР РНР ТЮЙР, ВРН МЕЙН╜РНПШЕ ПЮАНРШ, Б ЙНРНПШУ СРБЕПФДЮКНЯЭ ОПНРХБМНЕ, АШКХ БШ╜ОНКМЕМШ МЮ МЕАНКЭЬНЛ ЛЮРЕПХЮКЕ Х ХУ ПЕГСКЭРЮРШ Б ДЮКЭ╜МЕИЬЕЛ МЕ ОНДРБЕПДХКХЯЭ. б ВЮЯРМНЯРХ, ХГЛЕМХКНЯЭ ОПЕДЯРЮБ╜КЕМХЕ Н РЕВЕМХХ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ БХПСЯЮ ЦПХООЮ б. хЯЯКЕДСЪ ОПЕДЯРЮБХРЕКЭМСЧ ЦПСООС ЬРЮЛЛНБ, л. Yamashita Х ЯНЮБР. (1988) ОНЙЮГЮКХ, ВРН НМЮ ХДЕР ОНЯРНЪММН БН БПЕЛЕМХ. б ЖЕКНЛ ЛНФМН ЯЙЮГЮРЭ, ВРН С пмй-ЦЕМНЛМШУ БХПСЯНБ ЯСЫЕ╜ЯРБСЧР ЛНКЕЙСКЪПМШЕ ВЮЯШ ЩБНКЧЖХХ Х ЩРХ ВЮЯШ ДБХФСРЯЪ ДЮ МЕЯЙНКЭЙН ОНПЪДЙНБ АШЯРПЕЕ, ВЕЛ С дмй-ЦЕМНЛМШУ БХПСЯНБ.
мЮХАНКЕЕ ХГБЕЯРМНИ ОПХВХМНИ БШЛХПЮМХЪ НПЦЮМХГЛНБ ЪБ╜КЪЕРЯЪ МЕЯОНЯНАМНЯРЭ'ОПХЯОНЯНАКЪРЭЯЪ Й ХГЛЕМЪЧЫХЛЯЪ ЮАХН╜РХВЕЯЙХЛ Х АХНРХВЕЯЙХЛ СЯКНБХЪЛ ЯПЕДШ ЯХКЮЛХ ЯБНЕЦН ЦЕМН╜ТНМДЮ. аШЯРПЮЪ ЛНКЕЙСКЪПМЮЪ ЩБНКЧЖХЪ пмй-ЦЕМНЛМШУ БХ╜ПСЯНБ ОНГБНКЪЕР МЮГБЮРЭ ЕЫЕ НДМС БНГЛНФМСЧ, РЕОЕПЭ СФЕ БМСРПЕММЧЧ ОПХВХМС БШЛХПЮМХЪ ≈ ПЮГПСЬЕМХЕ ЯКНФХБЬЕИЯЪ ЦЕМЕРХВЕЯЙНИ ЯРПСЙРСПШ ОНД БКХЪМХЕЛ ХГАШРЙЮ ЛСРЮЖХИ. мЕИ╜РПЮКЭМШИ УЮПЮЙРЕП ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ ГЮЛЕДКЪЕР ЩРНР ОПНЖЕЯЯ. йЮЙ ЦЕМЕРХВЕЯЙХИ ЛЮРЕПХЮК, РЮЙ ОПНДСЙРШ ЦЕМНБ АЕК╜ЙХ НАКЮДЮЧР ГМЮВХРЕКЭМНИ АСТЕПМНЯРЭЧ Х ОПХМХЛЮЧР МЮ ЯЕАЪ ОНРНЙ ЛСРЮЖХИ. мЕИРПЮКЭМШЕ ЛСРЮЖХХ МЮЙЮОКХБЮЧРЯЪ Б ЛНКЕ╜ЙСКЮУ PHK Х АЕКЙНБ, МН МЕ БШГШБЮЧР НРПХЖЮРЕКЭМНЦН ЩТТЕЙ╜РЮ. бШЯНЙЮЪ ЯЙНПНЯРЭ ЛСРЮЖХНММШУ ГЮЛЕЫЕМХИ Б ЦЕМЮУ ЯНБЛЕ╜ЫЮЕРЯЪ Я ЯНУПЮМЕМХЕЛ ФХГМЕММН БЮФМШУ ТСМЙЖХИ. яНГДЮЕРЯЪ ГЮЫХРЮ НР ТНПЯХПНБЮММНИ ЩБНКЧЖХХ. бНГМХЙЮЕР БНОПНЯ, ЙЮЙ ДНКЦН СЯРЮМНБХБЬХИЯЪ АЮПЭЕП ЛНФЕР БШДЕПФХБЮРЭ ДЮБКЕМХЕ ЛСРЮЖХИ. оПХ НАЯСФДЕМХХ ЩРНЦН .БНОПНЯЮ ЯКЕДСЕР НАПЮРХРЭЯЪ Й ОПНАКЕЛЕ ЛЮЯЯНБШУ БШЛХПЮМХИ ЙЮЙ Й НАЫЕАХНКНЦХВЕЯЙНЛС ЪБКЕМХЧ.
оНЯРНЪММН ОНБРНПЪЧЫХЛЯЪ ЩОХГНДНЛ ФХГМХ МЮ гЕЛКЕ ЪБ╜КЪЧРЯЪ ЛЮЯЯНБШЕ БШЛХПЮМХЪ БХДНБ Х АНКЕЕ БШЯНЙХУ РЮЙЯНМН╜ЛХВЕЯЙХУ ЦПСОО НПЦЮМХГЛНБ. б ЛЕГНГНЕ ЩРХ ЯНАШРХЪ ОПНХЯУН╜ДХКХ Я ОЕПХНДХВМНЯРЭЧ 26 ЛДМ КЕР, Ю Б ЙЮИМНГНЕ ≈ 35≈40 ЛКМ КЕР. лЮЯЯНБШЕ БШЛХПЮМХЪ МЕ МНЯЪР БЯЕНАЫЕЦН УЮПЮЙРЕПЮ. хУ ХГАХПЮРЕКЭМНИ ФЕПРБНИ . ЯРЮМНБЪРЯЪ НОПЕДЕКЕММШЕ РЮЙЯНМШ. оПХВХМШ ЛЮЯЯНБШУ БШЛХПЮМХИ МЕДНЯРЮРНВМН ЪЯМШ. лМНЦНВХЯ╜КЕММШЕ ЦХОНРЕГШ, ЯБЪГШБЮЧЫХЕ ЛЮЯЯНБШЕ БШЛХПЮМХЪ Я БМЕЬ╜МХЛХ ОПХВХМЮЛХ (ЛХПНБШЕ ЙЮРЮЯРПНТШ, БШГБЮММШЕ ОЮДЕМХЕЛ ЮЯРЕПНХДНБ, БЯОШЬЙЮЛХ ЯБЕПУМНБШУ ГБЕГД, ОПНУНФДЕМХЕЛ He-
ЛЕГХДШ Х Р. Д.)> ОПХЦНДМШ ДКЪ ВЮЯРМШУ ЯКСВЮЕБ, МН МЕ НАЗЪЯР МЪЧР БЯЕЦН ЪБКЕМХЪ Б ЖЕКНЛ. оН ЛМЕМХЧ л. б. бНКЭЙЕМЬРЕИМЮ Х р. я. пЮЯЯЮ (1987), ЛЮЯЯНБШЕ БШЛХПЮМХЪ ГЮБХЯЪР МЕ РНКЭЙН НР ХГЛЕМЕМХЪ БМЕЬМХУ АХНРХВЕЯЙХУ Х ЮАХНРХВЕЯЙХУ ТЮЙРНПНБ, МН Х НР БМСРПЕММХУ ОПХВХМ. бХДШ ЛНЦСР БШЛХПЮРЭ Б ПЕГСКЭРЮ╜РЕ МЮЙНОКЕМХЪ МЕИРПЮКЭМШУ ЛСРЮЖХИ, ЙНРНПШЕ ЯРЮМНБЪРЯЪ БПЕД╜МШЛХ ОПХ ЯБНЕЛ ЯНВЕРЮМХХ. лЕУЮМХГЛ БШЛХПЮМХЪ ЯБЪГЮМ РЮЙ╜ФЕ Я ОЯЕБДНМЕИРПЮКЭМШЛХ ЛСРЮЖХЪЛХ, НЙЮГШБЮЧЫХЛХ КХЬЭ МЕГМЮВХРЕКЭМНЕ НРПХЖЮРЕКЭМНЕ БНГДЕИЯРБХЕ. мЮЙНОКЕМХЕ МЕИ╜РПЮКЭМШУ ЛСРЮЖХИ ЛНФЕР МЮПСЬХРЭ ЯНЯРНЪМХЕ ЙНЛОЕМЯЮЖХХ ОЯЕБДНМЕИРПЮКЭМШУ ЛСРЮЖХИ Х ОПХБЕЯРХ Й СУСДЬЕМХЧ ОПХЯОН╜ЯНАКЕММНЯРХ Х ЦХАЕКХ БХДЮ. бШЛХПЮМХЕ Б ЖЕКНЛ ЕЯРЭ ЮБРНЙЮ╜РЮКХРХВЕЯЙХИ ОПНЖЕЯЯ. ╚бХДХНХЯВЕГМНБЕМХЕ╩ МЮЯРСОЮЕР БМЕ╜ГЮОМН Х МНЯХР УЮПЮЙРЕП ТЮГНБНЦН ОЕПЕУНДЮ. яКЕДСЕР ДНАЮБХРЭ, ВРН ЯНАШРХЪ ЛНЦСР ОНКСВХРЭ Х ДПСЦНЕ МЮОПЮБКЕМХЕ. б ЩРНЛ ЯКСВЮЕ ОЕПЕЯРПНИЙЮ ЦЕМНЛЮ ДЮЕР ОНКНФХРЕКЭМШИ ЩТТЕЙР. яН╜ГДЮЧРЯЪ ОПЕДОНЯШКЙХ ДКЪ ДЮКЭМЕИЬЕЦН ЩБНКЧЖХНММНЦН ПЮГБХ╜РХЪ. б ЩБНКЧЖХНММНИ ХЯРНПХХ АХНЯТЕПШ НДМХ БХДШ ОНЦХАЮКХ, МЕ НЯРЮБХБ ОНРНЛЯРБЮ, Ю ДПСЦХЕ ДЮБЮКХ БЕРБХ ОНЯКЕДСЧЫЕИ ЩБНКЧЖХХ.
щРЮ ЦХОНРЕГЮ ОПХЛЕМХЛЮ Й пмй-ЦЕМНЛМШЛ БХПСЯЮЛ Я ХУ БШЯНЙНИ ЯЙНПНЯРЭЧ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ. лНФМН ОПЕДОН╜КНФХРЭ, ВРН БМСРПЕММХЕ ОПХВХМШ, ЙНРНПШЕ ЯМХФЮЧР ФХГМЕЯОН╜ЯНАМНЯРЭ БХПСЯЮ Х ЯНГДЮЧР ОПЕДОНЯШКЙХ ДКЪ ЕЦН ХЯВЕГМНБЕМХЪ, НОПЕДЕКЪЧРЯЪ ОПНЦПЕЯЯХПСЧЫХЛ МЮЙНОКЕМХЕЛ МЕИРПЮКЭМШУ Х ОЯЕБДНМЕИРПЮКЭМШУ ЛСРЮЖХИ. оЕПБНМЮВЮКЭМН ОНЯКЕДМХЕ ОПХМН╜ЯЪР КХЬЭ МЕГМЮВХРЕКЭМШИ БПЕД. нДМЮЙН Б ДЮКЭМЕИЬЕЛ ХГЛЕ╜МЕМХЪ СЛМНФЮЧРЯЪ. бНГЛНФМНЯРЭ ЙНЛОЕМЯЮЖХХ ЛСРЮЖХИ СЛЕМЭ╜ЬЮЕРЯЪ. щРНР ОПНЖЕЯЯ МЕНРБПЮРХЛ, ЙЮЙ Х ЯРЮПЕМХЕ. б ПЕГСКЭРЮ╜РЕ БШПНФДЕММНЯРХ ЦЕМЕРХВЕЯЙНЦН ЙНДЮ Х БШПНФДЕММНЯРХ ЙНП╜ПЕКЪЖХХ ЛЕФДС ОЕПБХВМНИ Х ОПНЯРПЮМЯРБЕММНИ ЯРПСЙРСПНИ АЕК╜ЙЮ ХГЛЕМЕМХЪ МЮЙЮОКХБЮЧРЯЪ ОНДЯОСДМН. гЮРЕЛ ,ЯКЕДСЕР ТЮГН╜БШИ ОЕПЕУНД, Б ЯБНЕИ НЯМНБЕ ХЛЕЧЫХИ ПЕГЙСЧ ОЕПЕЯРПНИЙС ЦЕМНЛЮ Х БМЕГЮОМНЕ ХГЛЕМЕМХЕ ОПНЯРПЮМЯРБЕММНИ ЯРПСЙРСПШ ЛНКЕЙСК АЕКЙЮ. мЮЯРСОЮЧР ТСМЙЖХНМЮКЭМШЕ МЮПСЬЕМХЪ. фХГ╜МЕЯОНЯНАМНЯРЭ БХПСЯЮ ЯМХФЮЕРЯЪ. яНГДЮЧРЯЪ ОПЕДОНЯШКЙХ ДКЪ ЕЦН ХЯВЕГМНБЕМХЪ.
нДПЪУКЕМХЕ, ЯРЮПНЯРЭ Х ЯЛЕПРЭ ≈ ЩРН СДЕК БЯЕЦН ФХБНЦН. й РЮЙНЛС ХРНЦС ОПХУНДЪР МЕ РНКЭЙН ХМДХБХДСЮКЭМШИ НПЦЮМХГЛ, МН Х ЯННАЫЕЯРБЮ. с БШЯЬХУ ЩСЙЮПХНРНБ, ПЮГЛМНФЮЧЫХУЯЪ ОН╜КНБШЛ ОСРЕЛ, ЕДХМХЖЕИ БШЛХПЮМХЪ ЪБКЪЕРЯЪ БХД. бХДНБНЕ ЯН╜НАЫЕЯРБН НАКЮДЮЕР НАЗЕДХМЕММШЛ ЦЕМНТНМДНЛ, ЙНРНПШИ ЖЕКХ╜ЙНЛ БНБКЕЙЮЕРЯЪ Б ОПНЖЕЯЯ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ, ЯНГДЮЧ╜ЫЕИ ОПЕДОНЯШКЙХ ДКЪ БШЛХПЮМХЪ. бЯЕ НЯНАХ, ЯНЯРЮБКЪЧЫХЕ БХД, ПЮГДЕКЪЧР НАЫСЧ ЯСДЭАС Х РЕПЪЧР ФХГМЕММШЕ ОНРЕМЖХХ. оНОСКЪЖХЪ Х БХД С пмй-ЦЕМНЛМШУ БХПСЯНБ ХЛЕЧР ХМНЕ ЦЕМЕ╜РХВЕЯЙНЕ ЯРПНЕМХЕ. нМХ ОПЕДЯРЮБКЪЧР ЯНАНИ ЯНБНЙСОМНЯРЭ нр╜
ДЕКЭМШУ ЙКНМНБ, ЙНРНПШЕ Б ОНЯКЕДНБЮРЕКЭМНЛ ПЪДС ОНЙНКЕМХИ НАПЮГСЧР НАНЯНАКЕММШЕ ВХЯРШЕ КХМХХ. цЕМЕРХВЕЯЙХИ НАЛЕМ ЛЕФДС МХЛХ МЕ ОПНХЯУНДХР ЯНБЯЕЛ ХКХ ФЕ НМ НВЕМЭ ЯКЮА. гЮ ЯВЕР ЯКСВЮИМШУ ОПНЖЕЯЯНБ ЛНКЕЙСКЪПМЮЪ ЩБНКЧЖХЪ ЯНЯРЮБКЪЧ╜ЫХУ ОНОСКЪЖХЧ КХМХИ МНЯХР ХМДХБХДСЮКЭМШИ УЮПЮЙРЕП. б НД╜МХУ ЯКСВЮЪУ ЩБНКЧЖХЪ ОПНРЕЙЮЕР АКЮЦНОПХЪРМН Х ОЕПЕЯРПНИЙЮ ЦЕМНЛЮ МЕ ОПЕОЪРЯРБСЕР ОПНДНКФЕМХЧ ПНДЮ Х ДЮКЭМЕИЬЕЛС ЯСЫЕЯРБНБЮМХЧ КХМХХ, Ю Б ДПСЦХУ≈ЯМХФЮЕР ФХГМЕЯОНЯНА╜МНЯРЭ БХПСЯЮ. рЮЙХЕ КХМХХ ЯРЮМНБЪРЯЪ ФЕПРБЮЛХ БШЛХПЮМХЪ.
бШПЮФЕМХЕЛ ЩРХУ ОПНЖЕЯЯНБ ОПХ ЕЯРЕЯРБЕММНЛ ЙПСЦНБНПН╜РЕ БХПСЯНБ ЯКСФЮР БМЕГЮОМНЕ ОПЕЙПЮЫЕМХЕ ЩОХДЕЛХИ Х ХЯВЕГ╜МНБЕМХЕ ЖХПЙСКХПСЧЫХУ ЬРЮЛЛНБ. пЕГЙХИ ЯОЮД ЩОХДЕЛХВЕЯЙХУ БЯОШЬЕЙ Х АШЯРПНЕ ЯНЙПЮЫЕМХЕ ГЮАНКЕБЮЕЛНЯРХ, ЙНРНПШЕ ОПН╜ХЯУНДЪР, МЕЯЛНРПЪ МЮ МЮКХВХЕ БНЯОПХХЛВХБНЦН МЮЯЕКЕМХЪ, НР╜ЛЕВЮЧРЯЪ ОПХ ЦПХООЕ МЮ ОПНРЪФЕМХХ ЩПШ ЦНЯОНДЯРБЮ РНЦН ХКХ ХМНЦН ЬХТРНБНЦН БЮПХЮМРЮ БХПСЯЮ ЦПХООЮ ю. яНГДЮЕРЯЪ БОЕ╜ВЮРКЕМХЕ, ВРН ХМТЕЙЖХЪ ОНЯРЕОЕММН ГЮРСУЮЕР ОПХ ОЕПЕДЮВЕ НР ╚УНГЪХМЮ╩ Й ╚УНГЪХМС╩ [Kendal A. P., 1987]. оЕПХНДШ ОПЕНА╜КЮДЮМХЪ ЙЮФДНЦН ЬХТРНБНЦН БЮПХЮМРЮ БХПСЯЮ ЦПХООЮ ю РЮЙФЕ ОПЕЙПЮЫЮКХЯЭ БМЕГЮОМН. еЯКХ ХГЛЕПЪРЭ ЯНАШРХЪ Б ЦКНАЮКЭМНЛ ЛЮЯЬРЮАЕ, РН ЛНФМН ЯЙЮГЮРЭ, ВРН БХПСЯ СРПЮВХБЮКЯЪ ХГ ЖХП╜ЙСКЪЖХХ ЛЕМЭЬЕ ВЕЛ ГЮ ЦНД. бМЕГЮОМНЕ ОПЕЙПЮЫЕМХЕ ЖХПЙС╜КЪЖХХ ОПЕФДЕ ЦНЯОНДЯРБНБЮБЬХУ ЬРЮЛЛНБ МЕКЭГЪ НАЗЪЯМХРЭ РНКЭЙН ДЮБКЕМХЕЛ ЙНККЕЙРХБМНЦН ХЛЛСМХРЕРЮ. оН-БХДХЛНЛС, ЬХТРНБШЕ ЬРЮЛЛШ БХПСЯЮ ЦПХООЮ к Х ХУ ДПЕИТНБШЕ БЮПХЮМРШ Б ПЪДЕ ЯКСВЮЕБ ЯОНЯНАМШ ОПНИРХ КХЬЭ НЦПЮМХВЕММНЕ ВХЯКН ЦЕМЕПЮЖХИ, МЕ ЛНЦСР АЕЯЙНМЕВМН ОЕПЕДЮБЮРЭЯЪ НР ╚УНГЪХМЮ╩ Й ╚УНГЪХМС╩ Х ГЮУНДЪР Б ЙНМЖЕ ЙНМЖНБ Б ЩБНКЧЖХНММШИ РСОХЙ. бНГЛНФМНЯРЭ ЯМХФЕМХЪ ФХГМЕЯОНЯНАМНЯРХ Х ЦХАЕКХ БХПСЯЮ НОПЕДЕКЪЕРЯЪ РЕЛ, ВРН ЬРЮЛЛШ, БШГШБЮЧЫХЕ ЩОХДЕЛХХ Х ОЮМ╜ДЕЛХХ, ОПНХЯУНДЪР НР ЯЕКЕЙРХБМН ЖЕММШУ ЛСРЮМРНБ ХКХ ПЕЮЯ- ЯНПРЮМРНБ Х ЦЕМЕЮКНЦХВЕЯЙХ ОПЕДЯРЮБКЪЧР ЯНАНИ ВХЯРШЕ КХМХХ. лНКЕЙСКЪПМЮЪ ЩБНКЧЖХЪ ЛМНЦХУ ХГ МХУ ОПНУНДХР МЕАКЮЦНОПХ╜ЪРМН Х МЮЙКЮДШБЮЕР НЦПЮМХВЕМХЪ МЮ ОПНДНКФХРЕКЭМНЯРЭ ЯСЫЕ╜ЯРБНБЮМХЪ БХПСЯЮ. ...
нЯРЮЕРЯЪ МЕЪЯМШЛ, ПЮЯОПНЯРПЮМЪЕРЯЪ КХ ОПНЖЕЯЯ БШЛХПЮ╜МХЪ, ЙНРНПНЛС, БЕПНЪРМН, ОНДБЕПФЕМШ НРДЕКЭМШЕ ЬРЮЛЛШ, МЮ ОНОСКЪЖХХ ,Б ЖЕКНЛ Х БХД. мЕКЭГЪ ХЯЙКЧВХРЭ, ВРН БПЕЛЪ ЯСЫЕ╜ЯРБНБЮМХЪ ХГБЕЯРМШУ МЮ ЯЕЦНДМЪ пмй-ЦЕМНЛМШУ БХПСЯНБ Б ХУ ЯНБПЕЛЕММНИ ТНПЛЕ НЦПЮМХВЕМН. оН-БХДХЛНЛС, ОЕПЕД МХЛХ НР╜ЙПШБЮЧРЯЪ ДБЮ ОСРХ ≈ ОНЦХАМСРЭ ХКХ ХГЛЕМХРЭЯЪ. мЕЙНРНПШЕ пмй-ЦЕМНЛМШЕ БХПСЯШ Я БШЯНЙНИ ЯЙНПНЯРЭЧ ЩБНКЧЖХХ ЛНЦСР ЯРЮРЭ ФЕПРБЮЛХ БШЛХПЮМХЪ. бЛЕЯРЕ Я РЕЛ МЕКЭГЪ МЕ СВХРШБЮРЭ, ВРН БШЯНЙЮЪ ЯЙНПНЯРЭ ЩБНКЧЖХХ PHK ЯНВЕРЮЕРЯЪ Я ЯНУПЮМЕМХ╜ЕЛ ФХГМЕММН БЮФМШУ ТСМЙЖХИ, НАЕЯОЕВХБЮЧЫХУ ЦЕМЕРХВЕЯЙХИ ЦНЛЕНЯРЮГ. оНЛХЛН ЯОНЯНАМНЯРХ Й АШЯРПНИ ЩБНКЧЖХХ, ДКЪ пмй- ЦЕМНЛМШУ БХПСЯНБ УЮПЮЙРЕПЕМ ЦКСАНЙХИ ЩБНКЧЖХНММШИ ГЮЯРНИ,
ЙНРНПШИ БШПЮФЮЕРЯЪ Б ГМЮВХРЕКЭМНЛ ЙНМЯЕПБЮРХГЛЕ МЮЯКЕД╜ЯРБЕММНЯРХ Х ДКХРЕКЭМНЛ ЯНУПЮМЕМХХ ТСМДЮЛЕМРЮКЭМШУ ЯБНИЯРБ Х НЯМНБМШУ АХНКНЦХВЕЯЙХУ УЮПЮЙРЕПХЯРХЙ БНГАСДХРЕКЪ [жХ- КХМЯЙХИ Ъ. ъ., 1988]. яСДЪ ОН ОХЯЭЛЕММШЛ ОЮЛЪРМХЙЮЛ ХЯРН╜ПХХ, ФЕКРЮЪ КХУНПДЙЮ ОНПЮФЮКЮ ВЕКНБЕЙЮ МЕ ЛЕМЕЕ ВЕЛ 190 КЕР, ЙПШЛЯЙЮЪ ЦЕЛНППЮЦХВЕЯЙЮЪ КХУНПДЙЮ МЕ ЛЕМЕЕ ВЕЛ 600 КЕР Х АЕЬЕМЯРБН 2000 КЕР МЮГЮД. щРХНКНЦХВЕЯЙХЕ ЮЦЕМРШ ЩРХУ ХМ╜ТЕЙЖХИ ОПНМЕЯКХ ЯБНХ НОПЕДЕКЪЧЫХЕ БХДНБШЕ ЯБНИЯРБЮ, БШ╜ПЮФЮЧЫХЕЯЪ Б ЯОНЯНАМНЯРХ БШГШБЮРЭ УЮПЮЙРЕПМНЕ ГЮАНКЕБЮ╜МХЕ, ВЕПЕГ БЕЙЮ. нПХЕМРХПНБНВМН ЩРЮ ЩБНКЧЖХНММЮЪ ДХЯРЮМЖХЪ ДКЪ БХПСЯЮ ФЕКРНИ КХУНПЮДЙХ ЯННРБЕРЯРБСЕР БЕКХВХМЕ ОНПЪДЙЮ 130 000, Ю ДКЪ БХПСЯНБ ЙПШЛЯЙНИ ЦЕЛНППЮЦХВЕЯЙНИ КХУНПЮДЙХ Х АЕЬЕМЯРБЮ ≈ ЯННРБЕРЯРБЕММН 350 000 Х 145 000 ОНЙНКЕМХИ. щРЮ ДХЯРЮМЖХЪ ОН ВХЯКС ОНЙНКЕМХИ ОПЕБШЬЮЕР ПЮЯЯРНЪМХЕ, ЙН╜РНПНЕ НРДЕКЪЕР ЯНБПЕЛЕММНЦН ВЕКНБЕЙЮ НР ЕЦН НАЕГЭЪМНОНДНА╜МНЦН ОПЕДЙЮ. с пмй-ЦЕМНЛМШУ БХПСЯНБ НРЛЕВЮЧРЯЪ ДКХРЕКЭ╜МЮЪ ЙНМЯЕПБЮЖХЪ Х ЯНУПЮМЕМХЕ НОПЕДЕКЕММШУ ЦЕММШУ ЛНДСКЕИ,, ХЛЕЧЫХУ ДПЕБМЕЕ ОПНХЯУНФДЕМХЕ. мЮКХВХЕ РЮЙХУ ЛНДСКЕИ, ОПЕДЯРЮБКЪЧЫХУ ЯНАНИ ЯХЯРЕЛШ ПЕОКХЙЮЖХХ НДМНМХРЕБНИ PHK Я ОНКНФХРЕКЭМНИ ОНКЪПМНЯРЭЧ, НАЗЕДХМЪЕР Б ЕДХМШЕ ЦПСООШ ОХЙНПМЮ- Х яХМДАХЯ-ОНДНАМШУ БХПСЯНБ ДЮКЕЙН ПЮГНЬЕДЬХЕЯЪ Б ЯБНЕИ ЩБНКЧЖХХ БХПСЯШ ФХБНРМШУ Х ПЮЯРЕМХИ. щРХ БХПСЯШ ОПНЬКХ ДКХРЕКЭМШИ ЩБНКЧЖХНММШИ ОСРЭ НР НАЫХУ ОПЕДЙНБ, МН ЯНУПЮМХКХ ТСМЙЖХНМЮКЭМН БЮФМШИ СВЮЯРНЙ ЦЕМНЛЮ. бЯЕ ЩРН СЙЮГШБЮЕР МЮ РН, ВРН ЛЮЯЯНБШЕ БШЛХПЮМХЪ пмй-ЦЕМНЛМШУ БХ╜ПСЯНБ, ЕЯКХ НМХ Х ХЛЕЧР ЛЕЯРН, ХГАХПЮРЕКЭМШ. юМЮКНЦХВМШИ ОНПЪДНЙ МЮАКЧДЮЕРЯЪ ОПХ БШЛХПЮМХХ БХДНБ С БШЯЬХУ НПЦЮ╜МХГЛНБ.
оПНХЯУНФДЕМХЕ ОЮМДЕЛХВЕЯЙХУ .БЮПХЮМРНБ БХПСЯЮ ЦПХООЮ ю. жЕМРПЮКЭМШЛ Х ДН ЯХУ ОНП МЕПЕЬЕМ╜МШЛ БНОПНЯНЛ СВЕМХЪ Н ЦПХООЕ ЪБКЪЕРЯЪ БНОПНЯ НА ХЯРНВМХЙЮУ ОПНХЯУНФДЕМХЪ ОЮМДЕЛХВЕЯЙХУ БЮПХЮМРНБ БХПСЯЮ ЦПХООЮ ю, ЖХПЙСКХПСЧЫХУ Б ВЕКНБЕВЕЯЙНИ ОНОСКЪЖХХ, ХКХ, ХМЮВЕ, БНО╜ПНЯ Н ЛЕУЮМХГЛЕ' ЮМРХЦЕММНЦН ЯДБХЦЮ. б. л. фДЮМНБ ОПХДЕП╜ФХБЮКЯЪ ЩЙНКНЦХВЕЯЙНИ ЙНМЖЕОЖХХ Х ВПЕГБШВЮИМН ЛМНЦН ЯДЕ╜КЮК ДКЪ ЕЕ ПЮГБХРХЪ. яНЦКЮЯМН ДЮММНИ ЙНМЖЕОЖХХ, БНГМХЙМН╜БЕМХЕ ОЮМДЕЛХВЕЯЙХУ БЮПХЮМРНБ НАСЯКНБКЕМН БЙКЧВЕМХЕЛ Б ЦЕМНЛ БХПСЯЮ ЦПХООЮ ю ВЕКНБЕЙЮ ЦЕМНБ БХПСЯЮ ЦПХООЮ ФХ╜БНРМШУ.
щРЮ ЦХОНРЕГЮ АШКЮ БШЯЙЮГЮМЮ НЙНКН 20 КЕР МЮГЮД Х АШЯРПН ГЮБНЕБЮКЮ АНКЭЬСЧ ОНОСКЪПМНЯРЭ ЯПЕДХ ПЮГКХВМШУ ЯОЕЖХЮКХ╜ЯРНБ, ГЮМХЛЮЧЫХУЯЪ ХГСВЕМХЕЛ БХПСЯЮ ЦПХООЮ Х ЦПХООНГМНИ ХМТЕЙЖХХ ≈ БХПСЯНКНЦНБ, ЛНКЕЙСКЪПМШУ АХНКНЦНБ, АХНКНЦНБ, ЛЕДХЙНБ Х БЕРЕПХМЮПНБ. йЮГЮКНЯЭ, ВРН БНОПНЯ Н ОПНХЯУНФДЕМХХ ОЮМДЕЛХВЕЯЙХУ БЮПХЮМРНБ БХПСЯЮ ЦПХООЮ ю ОПХМЖХОХЮКЭМН ПЕ╜ЬХКЯЪ. ьРЮЛЛШ, БШГШБЮЧЫХЕ ОЮМДЕЛХХ, БНГМХЙЮЧР Б ПЕГСКЭ╜РЮРЕ ОЕПЕПЮЯОПЕДЕКЕМХЪ ТПЮЦЛЕМРНБ ЦЕМНЛЮ ЛЕФДС БХПСЯЮЛХ
ЦПХООЮ ВЕКНБЕЙЮ Х ФХБНРМШУ, ЩРХ БХПСЯШ НРМНЯЪРЯЪ Й НДМНЛС БХДС Х НАПЮГСЧР ЕДХМШИ ЦЕММШИ ОСК. нДМЮЙН ЬКН БПЕЛЪ, Ю ПЕЬЮЧЫХЕ ДНЙЮГЮРЕКЭЯРБЮ Б ОНКЭГС ЩЙНКНЦХВЕЯЙНИ ЙНМЖЕОЖХХ ОНКСВЕМШ МЕ АШКХ. лЮКН РНЦН, ЩРЮ ЙНМЖЕОЖХЪ ЯРЮКЮ ХЯОШРШ╜БЮРЭ РПСДМНЯРХ Б ЯБЪГХ Я МЮЙНОКЕМХЕЛ ТЮЙРНБ Н яОЕЖХЮКХГЮ╜ЖХХ БХПСЯЮ ЦПХООЮ ю Й ЯБНХЛ ╚УНГЪЕБЮЛ╩, Н ОПНАКЕЛЮРХВМНЯРХ ОПЕНДНКЕМХЪ БХДНБНЦН АЮПЭЕПЮ, НА НАПЮГНБЮМХХ ДХЯЦЮПЛНМХВ╜МШУ ЯНВЕРЮМХИ ЦЕМНБ ОПХ ХУ ОЕПЕПЮЯОПЕДЕКЕМХХ, Н МХГЙНИ ФХГ╜МЕЯОНЯНАМНЯРХ ПЕЮЯЯНПРЮМРНБ БХПСЯНБ ЦПХООЮ ВЕКНБЕЙЮ Х ФХ╜БНРМШУ Х Р. Д. б ПЕГСКЭРЮРЕ ОЕПЕЯНПРХПНБЙХ ТПЮЦЛЕМРНБ ЦЕ╜МНЛЮ ЛЕФДС БХПСЯЮЛХ ЦПХООЮ ВЕКНБЕЙЮ Х ФХБНРМШУ Б СЯКНБХ╜ЪУ КЮАНПЮРНПХХ АШКХ ОНКСВЕМШ ПЕЮЯЯНПРЮМРШ, ЙНРНПШЕ ХЛЕЧР ЮМРХЦЕММШЕ ТНПЛСКШ, ЯННРБЕРЯРБСЧЫХЕ РЮЙНБШЛ ХГБЕЯРМШУ ОЮМДЕЛХВЕЯЙХУ БЮПХЮМРНБ. щРН, НДМЮЙН, МЕ ОНДЙПЕОХКН ЩЙНКН╜ЦХВЕЯЙСЧ ЙНМЖЕОЖХЧ. пЕЮЯЯНПРЮМРШ ОПЕДЯРЮБКЪКХ ЯНАНИ КХЬЭ ЮМРХЦЕММШЕ, Ю МЕ АХНКНЦХВЕЯЙХЕ ЮМЮКНЦХ ОЮМДЕЛХВЕЯЙХУ БЮПХ╜ЮМРНБ. оЕПЕЯНПРХПНБЙЮ ТПЮЦЛЕМРНБ ЦЕМНЛЮ МЮПСЬЮКЮ БХДНЯОЕ╜ЖХТХВЕЯЙСЧ БХПСКЕМРМНЯРЭ БХПСЯЮ Х КХЬЮКЮ ЕЦН ОНРЕМЖХХ Й ОЕПЕДЮВЕ Х ЩОХДЕЛХВЕЯЙНЛС ПЮЯОПНЯРПЮМЕМХЧ Б ОНОСКЪЖХХ. щЙНКНЦХВЕЯЙЮЪ ЙНМЖЕОЖХЪ МЕ ЯЛНЦКЮ НАЗЪЯМХРЭ ОПХВХМШ ОН╜ЯКЕДМЕЦН ЮМРХЦЕММНЦН ЯДБХЦЮ, ХЛЕБЬЕЦН ЛЕЯРН Б 1977 Ц. Х БШ╜ПЮГХБЬЕЦНЯЪ БН БМЕГЮОМНЛ БНГБПЮЫЕМХХ Б ЖХПЙСКЪЖХЧ БХПСЯЮ ЦПХООЮ A (HlNl).оПНХЯУНФДЕМХЕ ЩРНЦН БХПСЯЮ МЕ АШКН ЯБЪ╜ГЮМН Я ОЕПЕПЮЯОПЕДЕКЕМХЕЛ- ЦЕМЕРХВЕЯЙНЦН ЛЮРЕПХЮКЮ ЛЕФДС БХПСЯЮЛХ ЦПХООЮ ЙХРНБ, ЯБХМЕИ Х ОРХЖ. ьРЮЛЛШ 1977 Ц. НЙЮ╜ГЮКХЯЭ ЮМЮКНЦХВМШ ЬРЮЛЛЮЛ БХПСЯЮ ЦПХООЮ A (HlNl),ЖХПЙС╜КХПНБЮБЬХЛ Б ВЕКНБЕВЕЯЙНИ ОНОСКЪЖХХ Б 1950≈1952 ЦЦ. оПХ╜БЕДЕММШЕ ТЮЙРШ СЙЮГШБЮЧР ,МЮ ОПЪЛСЧ ОПЕЕЛЯРБЕММНЯРЭ ЬРЮЛ╜ЛНБ Х ДКХРЕКЭМНЕ ЯНУПЮМЕМХЕ БХПСЯЮ, НАЕЯОЕВХБЬЕЕ БНГЛНФ╜МНЯРЭ ЕЦН БНГБПЮЫЕМХЪ Б ЖХПЙСКЪЖХЧ.
бЯЕ ЩРН ОНБШЯХКН ХМРЕПЕЯ Й ЮКЭРЕПМЮРХБМНЛС ЛМЕМХЧ, ЯН╜ЦКЮЯМН ЙНРНПНЛС ЦПХОО ю ВЕКНБЕЙЮ ЪБКЪЕРЯЪ ЮМРПНОНМНГМНИ ХМТЕЙЖХЕИ, ЙНРНПЮЪ БШГШБЮЕРЯЪ ЬРЮЛЛЮЛХ, ЯБЪГЮММШЛХ ХЯЙ╜КЧВХРЕКЭМН Я ВЕКНБЕВЕЯЙНИ ОНОСКЪЖХЕИ. оЕПЕУНД ЦЕМНБ ХГ ╚ФХБНРМШУ ПЕГЕПБСЮПНБ╩, ДЮФЕ ЕЯКХ ЩРН БНГЛНФМН, МЕ ХЛЕЕР НРМНЬЕМХЪ Й ЮМРХЦЕММНЛС ЯДБХЦС. оНЯКЕДМХИ БНГМХЙЮЕР Б ПЕ╜ГСКЭРЮРЕ БНГБПЮЫЕМХЪ Б ЖХПЙСКЪЖХЧ ЬРЮЛЛНБ, ДКХРЕКЭМН ЯН╜УПЮМЪЧЫХУЯЪ Б ВЕКНБЕВЕЯЙНИ ОНОСКЪЖХХ. йЮЙ СВЕМШИ ЬХПНЙН╜ЦН ОПНТХКЪ Х ЩМЖХЙКНОЕДХВЕЯЙХ НАПЮГНБЮММШИ ВЕКНБЕЙ б. л. фДЮМНБ МЕ ГЮЛШЙЮКЯЪ Б ПЮЛЙЮУ РЕУ БГЦКЪДНБ, ЙНРНПШЛ НРДЮБЮК ОПЕДОНВРЕМХЕ, Ю ЯРПЕЛХКЯЪ НУБЮРХРЭ БЯЧ ОПНАКЕЛС Б ЖЕКНЛ. щРН НРМНЯХКНЯЭ Х Й СВЕМХЧ Н ЦПХООЕ. б. л. фДЮМНБ АШК ЯРНПНММХЙНЛ ОКЧПЮКХГЛЮ ЛМЕМХИ Х ОНКЮЦЮК, ВРН РЮЙЮЪ ХДЕНКНЦХЪ НАЕЯОЕВХБЮЕР ОПНЦПЕЯЯ МЮСЙХ. б ЩРНИ ЯБЪГХ ХМРЕПЕЯ╜МН НЯРЮМНБХРЭЯЪ МЮ ОНКНФЕМХЪУ ЮМРПНОНМНГМНИ ЦХОНРЕГШ ОПН╜ХЯУНФДЕМХЪ ОЮМДЕЛХВЕЯЙХУ БЮПХЮМРНБ ЦПХООЮ ю ВЕКНБЕЙЮ Х МЮ ЕЕ ПЮГБХРХХ.
яНБПЕЛЕММШЕ БГЦКЪДШ МЮ ЦПХООНГМСЧ ХМТЕЙЖХЧ ЙЮЙ МЮ ЮМРПНОНМНГ НАНЯМНБЮМШ Б ПЮАНРЕ R. е. Hope-Simpson Х D. б. Go╜lubev (1987), БШДБХМСБЬХУ МНБСЧ ЙНМЖЕОЖХЧ ЩОХДЕЛХВЕЯЙНЦН ОПНЖЕЯЯЮ ОПХ ЦПХООЕ ю. юБРНПШ НАПЮЫЮЧР БМХЛЮМХЕ МЮ РН, ВРН ВХЯКН ЬХТРНБШУ БЮПХЮМРНБ БХПСЯЮ, ЙНРНПШЕ БШГШБЮЧР ЦПХООЮ ю С ВЕКНБЕЙЮ, МЕ РПХ (HlNl 1946≈1957 ЦЦ. Х ОНЯКЕ 1977 r.; H2N2 1957≈1968 ЦЦ. Х H3N2ОНЯКЕ 1968 Ц.), Ю ВЕРШПЕ. вЕРБЕПРШЛ БЮПХЮМРНЛ ЪБКЪЕРЯЪ БХПСЯ ЦПХООЮ A (HONl),ОН╜ЪБХБЬХИЯЪ Б 1929 Ц. Х ЖХПЙСКХПНБЮБЬХИ ДН 1946 Ц. нОХПЮЪЯЭ МЮ ЯЕПНКНЦХВЕЯЙХЕ ДЮММШЕ, ЛНФМН ЦНБНПХРЭ РЮЙФЕ Н ОЪРНЛ ЬХТРНБНЛ БЮПХЮМРЕ ЦПХООЮ A (HswINl),ЖХПЙСКХПНБЮБЬЕЛ Б 1918≈1929 ЦЦ. Х ОЕПЕЬЕДЬЕЛ Б ,ОНОСКЪЖХЧ ЯБХМЕИ, ЦДЕ НМ ЯН╜УПЮМХКЯЪ ДН МЮЯРНЪЫЕЦН БПЕЛЕМХ. нАЗЕДХМЕМХЕ БХПСЯНБ ЦПХО╜ОЮ A (HswINl), (HONl)Х (HlNl)Б НДХМ ОНДРХО ОПХМЖХОХ╜ЮКЭМН МЕБЕПМН. щРХ БХПСЯШ БШГШБЮЧР МЕОНКМШИ ОЕПЕЙПЕЯРМШИ ХЛЛСМХРЕР.
оНЪБКЕМХЕ ЙЮФДНЦН ХГ МХУ МНЯХКН УЮПЮЙРЕП ЮМРХЦЕМ╜МНЦН ЯДБХЦЮ. б ЯКСВЮЕ ЯЛЕМШ БХПСЯЮ ЦПХООЮ A (HONl)МЮ (HlNl)ЩРНР ЯДБХЦ ОПНХГНЬЕК МЕ Б ПЕГСКЭРЮРЕ ОЕПЕЯНПРХПНБЙХ ТПЮЦЛЕМРНБ ЦЕМНЛЮ. дНЙСЛЕМРХПНБЮМН, ВРН ОПХВХМНИ БНГМХЙ╜МНБЕМХЪ БХПСЯЮ ЦПХООЮ A (HlNl)ОНЯКСФХКЮ ГМЮВХРЕКЭМЮЪ ЛС╜РЮЖХЪ Б ЦЕМЕ ДКЪ ЦЕЛЮЦЦКЧРХМХМЮ БХПСЯЮ ЦПХООЮ A (HONl). нЦПЮМХВЕММНЯРЭ ВХЯКЮ ОЮМДЕЛХВЕЯЙХУ БЮПХЮМРНБ БХПСЯЮ ЦПХО╜ОЮ ю ВЕКНБЕЙЮ МЕ МЮУНДХР НАЗЪЯМЕМХЪ Я ОНГХЖХИ ЩЙНКНЦХВЕЯЙНИ ЙНМЖЕОЖХХ. оН ЯЛШЯКС ЙНМЖЕОЖХХ, ЯБНАНДМЮЪ ОЕПЕЯНПРХПНБЙЮ ТПЮЦЛЕМРНБ ЦЕМНЛЮ ЛЕФДС БХПСЯЮЛХ ЦПХООЮ ВЕКНБЕЙЮ, ФХБНР╜МШУ Х ОРХЖ ЛНЦКЮ АШ ОПХБЕЯРХ Й НАПЮГНБЮМХЧ ДЕЯЪРЙНБ Х ЯН╜РЕМ ПЕЮЯЯНПРЮМРНБ, МЕ ХЯОШРШБЮЧЫХУ ХЛЛСММНЦН ДЮБКЕМХЪ ЯН ЯРНПНМШ ╚УНГЪХМЮ╩-ВЕКНБЕЙЮ Х ПЮЯОПНЯРПЮМЪЧЫХУЯЪ Б ОНОС╜КЪЖХХ.
щЙНКНЦХВЕЯЙЮЪ ∙ ЙНМЖЕОЖХЪ БНГМХЙМНБЕМХЪ ОЮМДЕЛХВЕЯЙХУ БЮПХЮМРНБ БХПСЯЮ ЦПХООЮ ю МЕ НАЗЪЯМЪЕР МЕ РНКЭЙН НЦПЮМХ╜ВЕММНЯРЭ ХУ ВХЯКЮ, МН Х ПЕЖХЙКХВМНЯРЭ ОНЪБКЕМХЪ Б ВЕКНБЕВЕ╜ЯЙНИ ОНОСКЪЖХХ. оНПЪДНЙ ЯЛЕМШ БЮПХЮМРНБ ОПНЯКЕФЕМ МЮ ОПН╜РЪФЕМХХ IOO КЕР. яЕПНКНЦХВЕЯЙХЕ ХЯЯКЕДНБЮМХЪ ЯШБНПНРНЙ ЙПНБХ ОНФХКШУ КЧДЕИ ОНЙЮГЮКХ, ВРН БЮПХЮМР H2N2,ОН-БХДХ╜ЛНЛС, ЖХПЙСКХПНБЮК Б ОНЯКЕДМЕИ ВЕРБЕПРХ XIX Б. мЮ ЯЛЕМС ЕЛС ОПХЬЕК БЮПХЮМР H3N2,ДНЛХМХПНБЮБЬХИ ,ОПЕДОНКНФХРЕКЭ╜МН Б 1900≈1918 ЦЦ. оПХАКХГХРЕКЭМН Б 1907 Ц. Й БЮПХЮМРС H3N2 (РЮЙ ФЕ ЙЮЙ Х Б МЮЬЕ БПЕЛЪ) ОПХЯНЕДХМХКЯЪ БЮПХЮМР HlNl. оНЯКЕДСЧЫЮЪ НВЕПЕДМНЯРЭ ЯЛЕМШ ЬРЮЛЛНБ ПЮЯЯЛЮРПХБЮКЮЯЭ БШЬЕ.
R. е. Hope-Simpson, D. б. Golubev (1987) НЯРЮМЮБКХБЮЧР╜ЯЪ МЮ ПЕЖХЙКХВМНЯРХ ЬХТРНБШУ БЮПХЮМРНБ БХПСЯЮ ЦПХООЮ ю ВЕКНБЕЙЮ Х НЦПЮМХВЕММНЯРХ ХУ ВХЯКЮ МЕ ДКЪ РНЦН, ВРНАШ ОНД╜ВЕПЙМСРЭ ЯКЮАНЯРЭ ЩЙНКНЦХВЕЯЙНИ ЦХОНРЕГШ. щРХ ТЮЙРШ ХЯ╜ОНКЭГСЧРЯЪ ДКЪ НАНЯМНБЮМХЪ ОПЕДКНФЕММНИ ХЛХ ЙНМЖЕОЖХХ.
юБРНПШ ПЮЯЯЛЮРПХБЮЧР ЩОХДЕЛХНКНЦХВЕЯЙХЕ ДЮММШЕ, ЯНЦКЮЯМН ЙНРНПШЛ ПЮГБХРХЕ БЯОШЬЕЙ Х ДБХФЕМХЕ ГЮАНКЕБЮЕЛНЯРХ ОПХ ЦПХООЕ МЕКЭГЪ НАЗЪЯМХРЭ РНКЭЙН МЕОПЕПШБМНИ Х ОНЯКЕДНБЮ╜РЕКЭМНИ ОЕПЕДЮВЕИ БХПСЯЮ НР АНКЭМНЦН Й АНКЭМНЛС, ЙЮЙ ЩРН ОПНХЯУНДХР ОПХ ЙНПХ. лМНЦНВХЯКЕММШЕ ОНОШРЙХ ОНЯРПНХРЭ ЛЮ╜РЕЛЮРХВЕЯЙХЕ ЛНДЕКХ ЩОХДЕЛХИ ЦПХООЮ, НЯМНБЮММШЕ МЮ ЩРНЛ ОПХМЖХОЕ, ГЮЙНМВХКХЯЭ АЕГПЕГСКЭРЮРМН. дБХФЕМХЕ ГЮАНКЕБЮЕ╜ЛНЯРХ ОПХ ЦПХООЕ Б ПЪДЕ ЯКСВЮЕБ МЮОНЛХМЮЕР ЛНДЕКЭ, ОПХ ЙН╜РНПНИ ХМТЕЙЖХЪ БНГМХЙЮЕР БН ЛМНЦХУ ОЕПБНМЮВЮКЭМШУ ТНЙС╜ЯЮУ Х ГЮРЕЛ ГЮРСУЮЕР ОПХ ОЕПЕДЮВЕ НР ╚УНГЪХМЮ╩ Й ╚УНГЪХМС╩. щРН ОНДРБЕПФДЮЕРЯЪ ЩОХДЕЛХНКНЦХВЕЯЙХЛХ МЮАКЧДЕМХЪЛХ НА НДМНБПЕЛЕММНЛ БНГМХЙМНБЕМХХ ЯКСВЮЕБ ЦПХООЮ МЮ АНКЭЬХУ РЕППХРНПХЪУ Х Б ПЮГНАЫЕММШУ ПЕЦХНМЮУ. уЮПЮЙРЕП РЮЙХУ ЩОХ╜ДЕЛХИ Б ЯБНЕ БПЕЛЪ ЯНГДЮБЮК ХККЧГХЧ ГЮМНЯЮ БХПСЯЮ ЦПХООЮ ХГ ЙНЯЛНЯЮ.
юБРНПШ ОПЕДОНКЮЦЮЧР, ВРН БХПСЯ ЦПХООЮ ю МЕ ЯОНЯНАЕМ АЕЯ╜ЙНМЕВМН ДНКЦН ОЕПЕДЮБЮРЭЯЪ НР ВЕКНБЕЙЮ Й ВЕКНБЕЙС Х ЯНУПЮ╜МЪЕРЯЪ Б ЛЕФЩОХДЕЛХВЕЯЙХИ ОЕПХНД МЕ ,Б НПЦЮМХГЛЕ ФХБНРМШУ Х ОРХЖ, Ю Б НПЦЮМХГЛЕ ВЕКНБЕЙЮ Б ТНПЛЕ КЮРЕМРМНИ ХКХ ОЕПЯХ╜ЯРЕМРМНИ ХМТЕЙЖХХ. вЕПЕГ ЛМНЦХЕ ЛЕЯЪЖШ Х ДЮФЕ ЦНДШ ОНД БКХЪМХЕЛ ТЮЙРНПНБ БМЕЬМЕИ ЯПЕДШ Х ОПХ ХГЛЕМЕМХХ ЯНЯРНЪМХЪ НПЦЮМХГЛЮ ╚УНГЪХМЮ╩ БХПСЯ ПЕЮЙРХБХПСЕРЯЪ С ЯЙПШРШУ МНЯХ╜РЕКЕИ Х ПЮЯОПНЯРПЮМЪЕРЯЪ Б ВЕКНБЕВЕЯЙНИ ОНОСКЪЖХХ. бМЕЬМХЕ ТЮЙРНПШ ДЕИЯРБСЧР МЮ БЯЧ ОНОСКЪЖХЧ ╚УНГЪЕБ╩. оНЩРНЛС БХ╜ПСЯ БШДЕКЪЕРЯЪ НР ЛЮЯЯШ МНЯХРЕКЕИ Х ЩОХДЕЛХЪ МЮВХМЮЕРЯЪ ХГ ЛМНЦХУ ОЕПБНМЮВЮКЭМШУ ТНЙСЯНБ. гЮ ЯВЕР ЩРНЦН ЛЕУЮМХГЛЮ МНБШЕ ОНПЖХХ БХПСЯЮ БЙКЧВЮЧРЯЪ Б ЖХПЙСКЪЖХЧ Б ОПНЖЕЯЯЕ ПЮГБХРХЪ ЩОХДЕЛХХ. пЕЮЙРХБХПНБЮММШИ БХПСЯ БЯРПЕВЮЕРЯЪ Я ХЛЛСММНИ ГЮЫХРНИ ╚УНГЪХМЮ╩, Б ПЕГСКЭРЮРЕ ВЕЦН ОПНХЯУНДХР ЮМРХЦЕММШИ ДПЕИТ.
хМРЕПЕЯМН НРЛЕРХРЭ, ВРН ЩОХГННРХВЕЯЙХИ ОПНЖЕЯЯ ОПХ ЦПХО╜ОЕ С ДНЛЮЬМХУ ФХБНРМШУ ЛНФЕР МНЯХРЭ ОНДНАМШИ УЮПЮЙРЕП. б 50-У ЦНДЮУ Б яью НРЛЕВЮКНЯЭ НДМНБПЕЛЕММНЕ БНГМХЙМНБЕМХЕ БЯОШЬЕЙ ЦПХООЮ ю С ЯБХМЕИ МЮ ПЮГНАЫЕММШУ РЕППХРНПХЮКЭМН ТЕПЛЮУ. оПЕДОНКЮЦЮКНЯЭ, ВРН БХПСЯ ОЕПЯХЯРХПНБЮК С ФХБНР╜МШУ Х ЮЙРХБХПНБЮКЯЪ ОНД БКХЪМХЕЛ ЛЕРЕНПНКНЦХВЕЯЙХУ ТЮЙ╜РНПНБ.
оЕПХНДХВЕЯЙЮЪ ЮЙРХБЮЖХЪ БХПСЯЮ С ЯЙПШРШУ МНЯХРЕКЕИ Х ЕЦН ПЮЯОПНЯРПЮМЕМХЕ Б ВЕКНБЕВЕЯЙНИ ОНОСКЪЖХХ ОПХБНДЪР ЙЮЙ Й ПЮГБХРХЧ ЩОХДЕЛХИ, ЙНРНПШЕ Я ХМРЕПБЮКНЛ Б 1≈2 ЦНДЮ ЯКЕ╜ДСЧР ДПСЦ ГЮ ДПСЦНЛ Б ОЕПХНД ОПЕНАКЮДЮМХЪ РНЦН ХКХ ХМНЦН ЬХТРНБНЦН БЮПХЮМРЮ, РЮЙ Х Й ЮМРХЦЕММНЛС ЯДБХЦС Х БНГМХЙ╜МНБЕМХЧ ОЮМДЕЛХИ. бЯЕ 5 ЬХТРНБШУ БЮПХЮМРНБ БХПСЯЮ ЦПХО╜ОЮ ю ВЕКНБЕЙЮ ЯНУПЮМЪЧРЯЪ Б ОНОСКЪЖХХ Х ОЕПХНДХВЕЯЙХ ПЕ╜ЮЙРХБХПСЧРЯЪ. нЯНАЕММН ДКХРЕКЭМШИ КЮРЕМРМШИ ОЕПХНД ХЛЕКЮ ОЮМДЕЛХЪ БХПСЯЮ ЦПХООЮ A (H3N2^),ЙНРНПШИ ЯНУПЮМЪКЯЪ Б ВЕКНБЕВЕЯЙНИ ОНОСКЪЖХХ Я МЮВЮКЮ XX Б. Х МЕ ДЮБЮК ЩОХДЕЛХВЕ╜
ЯЙХУ ОПНЪБКЕМХИ ДН 1988 Ц. нРДЕКЭМШЕ БХПСЯНКНЦХВЕЯЙХЕ Х ЯЕ╜ПНКНЦХВЕЯЙХЕ МЮУНДЙХ СЙЮГШБЮЧР МЮ БНГЛНФМНЯРЭ ОПНДНКФХ╜РЕКЭМНИ ЙНМЯЕПБЮЖХХ ЮПУЮХВМШУ БЮПХЮМРНБ.
бЮФМСЧ ПНКЭ Б НАНЯМНБЮМХХ ЙНМЖЕОЖХХ, ОПЕДКНФЕММНИ R. E .Hope-Simpson Х D. б. Golubev (1987), ХЦПЮЧР ДЮММШЕ, ОНКСВЕММШЕ Б ЛЕФЩОХДЕЛХВЕЯЙХИ ОЕПХНД. б ОЕПХНДШ ЛЕФДС ЩОХДЕЛХЪЛХ МЮ ТНМЕ ОПЕЙПЮЫЕМХЪ ЖХПЙСКЪЖХХ БНГАСДХРЕКЪ НАШВМН ПЕЦХЯРПХПСЧРЯЪ НРДЕКЭМШЕ ЯКСВЮИ ЦПХООЮ, ХЛЕЧЫХЕ БХПСЯНКНЦХВЕЯЙНЕ ОНДРБЕПФДЕМХЕ. хУ БНГМХЙМНБЕМХЕ ЛНФМН ХЯРНКЙНБЮРЭ ЙЮЙ ПЕГСКЭРЮР ЮЙРХБЮЖХХ БХПСЯЮ С МНЯХРЕКЕИ Х ЕЦН ОНЯКЕДСЧЫЕЦН НЦПЮМХВЕММНЦН ПЮЯОПНЯРПЮМЕМХЪ.
яКЮАШЛ ЛЕЯРНЛ ЮМРПНОНМНГМНИ ЙНМЖЕОЖХХ Н ЦПХООЕ ю ВЕ╜КНБЕЙЮ Б ОПНЬКНЛ АШКН НРЯСРЯРБХЕ ДНЙЮГЮРЕКЭЯРБ ДКХРЕКЭМНЦН БХПСЯНМНЯХРЕКЭЯРБЮ. б МЮЯРНЪЫЕЕ БПЕЛЪ ОНКНФЕМХЕ ХГЛЕМХ╜КНЯЭ. йЮЙ ЯОПЮБЕДКХБН НРЛЕВЮЧР R. е. Hope-Simpson, D. б. Go╜lubev (1987), БНГЛНФМНЯРЭ КЮРЕМРМНИ Х ОЕПЯХЯРЕМРМНИ ХМТЕЙ╜ЖХХ ВЕКНБЕЙЮ БХПСЯНЛ ЦПХООЮ ю МЕ БШГШБЮЕР ЯНЛМЕМХИ, УНРЪ ТНПЛШ МНЯХРЕКЭЯРБЮ РПЕАСЧР ХГСВЕМХЪ. оН-БХДХЛНЛС, БЮФМСЧ ПНКЭ Б ЯРЮМНБКЕМХХ МНЯХРЕКЭЯРБЮ ХЦПЮЕР ДЕИЯРБХЕ ДЕТЕЙРМНЦН БХПСЯЮ. гМЮВХРЕКЭМШИ ОНГХРХБМШИ БЙКЮД Б ОПНАКЕЛС БМНЯЪР ДЮММШЕ Н ОЕПЯХЯРЕМРМНИ ХМТЕЙЖХХ БХПСЯЮ ЦПХООЮ ю Б ЙСКЭРС╜ПЮУ ЙКЕРНЙ. щРНР ТЮЙР МЕ ЪБКЪЕРЯЪ ОПЪЛШЛ ДНЙЮГЮРЕКЭЯРБНЛ Б ОНКЭГС ОЕПЯХЯРЕМЖХХ БХПСЯЮ Б НПЦЮМХГЛЕ Х ЕЦН ЯНУПЮМЕМХЪ Б ОНОСКЪЖХХ ╚УНГЪЕБ╩ ГЮ ЯВЕР СЙЮГЮММНЦН ЛЕУЮМХГЛЮ. рЕЛ МЕ ЛЕМЕЕ ДЮММШЕ, ОНКСВЕММШЕ МЮ СПНБМЕ ЙСКЭРСП ЙКЕРНЙ, СЙЮГШ╜БЮЧР МЮ РЮЙСЧ БНГЛНФМНЯРЭ.
нАЯСФДЮЪ ОПХВХМШ Х ОЕПХНДХВМНЯРЭ ПЕЮЙРХБЮЖХХ БХПСЯЮ ЦПХООЮ ю Б НПЦЮМХГЛЕ ВЕКНБЕЙЮ, ЯКЕДСЕР НРЛЕРХРЭ ЖХЙКХВМНЯРЭ ОПХПНДМШУ ЪБКЕМХИ. щРН НАЫЕАХНКНЦХВЕЯЙЮЪ ГЮЙНМНЛЕПМНЯРЭ. оНБРНПЪЕЛНЯРЭ Х ЖХЙКХВМНЯРЭ ОПНЖЕЯЯНБ ФХГМЕДЕЪРЕКЭМНЯРХ БШПЮФЮЧРЯЪ Б ЛМНЦНКЕРМХУ ЙНКЕАЮМХЪУ ВХЯКЕММНЯРХ ФХБНР╜МШУ, НАПЮРХЛШУ ОЕПЕЯРПНИЙЮУ БХДНБШУ ЯРПСЙРСП АХНЖЕМНГНБ, ОСКЭЯЮЖХХ НВЮЦНБ ГННМНГМШУ ХМТЕЙЖХИ, ХГЛЕМЕМХХ ЯБНИЯРБ БНГ╜АСДХРЕКЪ, ОНБШЬЕМХХ ЕЦН БХПСКЕМРМНЯРХ Х Р. Д. жХЙКШ ОПХПН╜ДШ НАСЯКНБКЕМШ БКХЪМХЕЛ ТЮЙРНПНБ БМЕЬМЕИ ЯПЕДШ, ЙНРНПШЕ ДЕИЯРБСЧР Х МЮ ВЕКНБЕЙЮ. яПЕДХ ЩРХУ ТЮЙРНПНБ БЮФМНЕ ЛЕЯРН ГЮМХЛЮЕР ЯНКМЕВМЮЪ ЩМЕПЦХЪ. мЮ ГМЮВЕМХЕ ОНРНЙЮ ЩМЕПЦХХ яНКМЖЮ СЙЮГШБЮК ЙПСОМЕИЬХИ СВЕМШИ ю. к. вХФЕБЯЙХИ (1976). б ЯБНЕИ ЙМХЦЕ ╚гЕЛМНЕ ЩУН ЯНКМЕВМШУ АСПЭ╩, БШЬЕДЬЕИ Б ЯБЕР Б 1937 Ц., НМ НРЛЕВЮК, ВРН ЯНКМЕВМЮЪ ЮЙРХБМНЯРЭ ЪБКЪЕРЯЪ ПЕ╜ЦСКЪРНПНЛ ЩОХДЕЛХВЕЯЙХУ ОПНЖЕЯЯНБ Б ЛЮЯЬРЮАЕ ОКЮМЕРШ. юМЮ╜КХГХПСЪ ЛЮРЕПХЮКШ Н БНГМХЙМНБЕМХХ Х РЕВЕМХХ ЦПХООНГМШУ ЩОХДЕЛХИ ОНВРХ ГЮ 500 КЕР, ю. к. вХФЕБЯЙХИ (1976) ОПХУНДХР Й БШБНДС, ВРН ОЕПХНД ОНБРНПЪЕЛНЯРХ ЩОХДЕЛХИ ЦПХООЮ (ХЛЕ╜ЧРЯЪ Б БХДС ОЮМДЕЛХХ) Б ЯПЕДМЕЛ ПЮБЕМ 11,3 ЦНДЮ. нРЙКНМЕ╜МХЕ МЮВЮКЭМШУ КЕР ЩОХДЕЛХХ НР ЛЮЙЯХЛСЛЮ ЯНКМЕВМНИ ДЕЪ-
еЫЕ ОН РЕЛЕ гюйкчвемхе:
- гЮЙКЧВЕМХЕ
- гЮЙКЧВЕМХЕ
- гЮЙКЧВЕМХЕ
- гЮЙКЧВЕМХЕ
- гЮЙКЧВЕМХЕ
- гЮЙКЧВЕМХЕ
- гЮЙКЧВЕМХЕ
- гЮЙКЧВЕМХЕ
- 2.5. пЮЖХНМЮКЭМНЕ РПСДНСЯРПНИЯРБН АНКЭМШУ ОН ГЮЙКЧВЕМХЧ йщй
- оЮРНЛНПТНКНЦХВЕЯЙНЕ ГЮЙКЧВЕМХЕ
- бЛЕЯРН ГЮЙКЧВЕМХЪ
- оЮПЮЦПЮТ ОЪРШИ. нА НАЫХУ ГЮЙКЧВЕМХЪУ ОН ДЕИЯРБХЪЛ ЛНГЦЮ
- лЕУЮМХГЛШ ГЮЙКЧВЕМХЪ ДНЦНБНПНБ
- гЮЙКЧВЕМХЕ: НР ОНМХЛЮМХЪ Й ДЕИЯРБХЧ
- оЮПЮЦПЮТ НДХММЮДЖЮРШИ. гЮЙКЧВЕМХЪ ОН ТНПЛЕ ЦНКНБШ
- оЮПЮЦПЮТ ВЕРБЕПРШИ. гЮЙКЧВЕМХЪ ОН ЯХЛОРНЛЮЛ КХУНПЮДНЙ
- гЮЙКЧВЕМХЕ
- гЮЙКЧВЕМХЕ
- гЮЙКЧВЕМХЕ