ГЛАВА 31. ВИРУСЫ ОСПЫ
Семейство вирусов оспы (Poxviridae)подразделяется на два подсемейства — вирусы оспы хордовых (Chordopoxvirinae) и оспы насекомых (Entomopoxvirinae).В первом подсемействе выделены 6 родов вирусов: Orthopoxvirus(оспы), Parapoxvirus(паравакцины), Avipoxvirus(оспы птиц), Capripoxvirus(оспы овец), Leporipoxvirus(миксомы) и Suipoxvirus (оспы свиней).
В составе каждого рода имеется либо один, либо (чаще) несколько вирусов. Во втором подсемействе выделены 3 рода на основе морфологических особенностей строения вирионов, молекулярной массы геномов, круга поражаемых «хозяев»: вирусы А оспы шейкокрылых (Colioptera), вирусы В оспы чешуекрылых (Lepidoptera)и вирусы Cоспы двукрылых (Diptera). В отличие от вирусов оспы позвоночных вирусы, поражающие насекомых, серологически неродственны как между родами, так и в пределах одного и того же рода. Поэтому и классификацию их следует считать предварительной. Кроме того, несколько вирусов оспы не классифицированы, среди них вирусы оспы плотоядных, слонов, раккунов, вирусы оспы Тана и Яба, а также контагиозного моллюска [Matthews R., 1982].
Морфология вирионов оспы настолько характерна у представителей обоих подсемейств, что не было смысла подразделять представителей, поражающих столь отдаленных филогенетически «хозяев», на таксономические группы более высокого ранга. Вирионы поксвирусов имеют овальную или кйрпи- чеобразную форму с соотношением длин осей 1,2—1,7. Их размер 140—170?220—450 нм, эта величина приближается к размеру наиболее мелких бактерий (риккетсии, хламидии, микоплазмы). Строение их не укладывается в рамки типов симметрии других вирусов и отличается деталями у представителей разных родов. Общим для всех вирусов данного семейства является наличие сердцевины в виде вогнутой линии, внешней оболочки и боковых тел, как бы сплющивающих сердцевину (нуклеоид).
Нуклеоид содержит ДНК, ассоциированную с белками.
Способ ее упаковки неясен, хотя из комплекса ДНК—белок выделены 3 белка с молекулярной массой 33 000, 28 000 и 12 000, последний белок богат аргинином. Напрашивается аналогия с упаковкой ДНК хромосом эукариотов гистоновыми и негистоновыми белками. Нуклеоид окружен внутренними оболочками. Внешние оболочки состоят из липидов и углеводов. Оболочки построены сложно и состоят из слоев трубчатых или глобулярных структур. Разные роды вирусов различаются преимущественно деталями строения внешних оболочек.Геном вирионов представляет собой линейную двунитевую ДНК с молекулярной массой 85?106—250?106, концы которой соединены ковалентно, образуя терминальные петли и инвертированные тандемные повторы. Молекулярная масса генома не может служить таксономическим признаком, так как у серологически родственных вирусов она колеблется от 85?106(вирус паравакцины) до 200?106(вирус оспы птиц). Содержание Г+Ц у вирусов оспы позвоночных составляет 5— 7,5% от массы вириона; состав нуклеотидов: А —29,5%, Ц — 20%, Г — 20,6%, T — 29,9%. Геном вируса осповакцины содержит всего 240 000 пар нуклеотидов и имеет длину 82 мкм. Необычная прочность ДНК этого вируса и других вирусов данной группы обусловлена ковалентными связями комплементарных нитей на обоих концах молекулы ДНК, примерно на расстоянии 50 нуклеотидов от ее концов [см. Слепуш- кин A. H., 1982]. Как было показано при изучении вируса осповакцины, ее ДНК, подобно ДНК многих других вирусов, содержит терминальные инвертированные повторы, которые, как предполагается, играют важную' роль в репликации ДНК. У вируса осповакцины эти повторы весьма велики — молекулярная масса около 7?106, или около 10 000 пар нуклеотидов. Они могут быть частично транскрибированы, кодируя синтез ранних мРНК [Wittek R. et al., 1980].
На геноме вирусов оспы закодировано несколько сотен белков (глико- и фосфопротеидов) с молекулярной массой 80 000— -200 000. По данным разных авторов, для различных вирусов число вирусспецифйческих белков варьирует.
Ранее указывалось, что вирионы осповакцины содержат более 80 белков, суммарная молекулярная масса которых составляет около 60% массы генома [Fenner F., 1979]. У вируса оспы Яба обнаружено 37 структурных белков с молекулярной массой 10 000—220 000 [Fenger.T., Rouhandch H., 1976]. Более поздние данные свидетельствуют, что вирус осповакцины имеет 279 белков, в том числе 13 гликопротеидов [Carrasco L., Bravo R., 1985]. Пока идентифицированы лишь немногие вирионные и и невирионные белки. Нейтрализация вируса связана с поверхностным белком (58 000), гемагглютинация — с гликопротеидом (85 000). Другие мажорные белки обозначают символами. 2Ь, 4a, 4b, 6a, 6b, 8, llb∖два белка (4a, 4Ь) составляют ■50% суммарных белков сердцевины, масса которой в свою очередь составляет около 50% массы вириона. В наружных •оболочках вириона найдено 8 белков, а на самой поверхности— 5 белков. В составе вирионов имеется или в процессе репродукции образуется не менее 10 ферментов, в том числе РНК-полимераза (вирионная транскриптаза), ферменты, катализирующие образование и метилирование кэп-структур, •которые обеспечивают синтез и процессинг мРНК, протеин- и тимидинкиназа. По-видимому, и синтез вирусной ДНК катализируется в основном вирусспецифическими ферментами [Sla- baugh M., Mattews C., 1984].Из нескольких (около 10) больших антигенов вирионов ■один перекрестно реагирует со всеми «членами» вирусов оспы •позвоночных. В пределах родов серологические связи разных вирусов выражены еще больше. На ранних стадиях репродукции вируса осповакцины синтезируется белок с молекулярной массой 19 000, сходный с фактором эмбрионального роста клеток млекопитающих (о возможном значении и происхождении этого белка см. ниже). В связи со сложными процессами синтеза и созревания вирусных белков и морфогенезом вирионов антигенная структура внутриклеточного и внеклеточного вирусов различна.
Репликация вирусов оспы (рис. 61) происходит в цитоплазме ступенчато, включая первичное раздевание (uncoatingI), «синтез ранних мРНК и белков, вторичное раздевание (uncoatingII), репликацию ДНК, синтез поздних мРНК и-белков, входящих в состав вирионов, их гликозилирование и фосфорилирование, образование предшественников вирионов, созре- .
вание их и выход из клетки [Shida H., 19861.Хотя имеются данные об участии клеточного ядра в репро-
■ дукции вирусов оспы [Miningan H. et al., 1985], но оно не
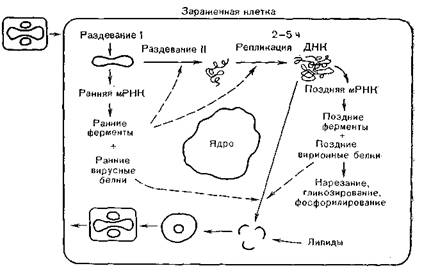
Рис. 61. Цикл репликации вируса осповакцины (схема).
обязательно, так как весь цикл репродукции вирусов оспы можно воспроизвести в безъядерных клетках [Villareal Е. et al., 1984]. Вместе с тем репродукция этих вирусов, если даже абсолютно исключить клеточно-зависимые синтезы белка, тесно связана с клеточным метаболизмом. Об этом свидетельствуют возможность репродукции штаммов вируса ocπσ- вакцины, дефектных по некоторым генам (например, по гену тимидинкиназы), возможность замены этого гена геном герпесвируса, а также наличие в геноме вируса достаточно обширных нереплицирующихся областей. Именно на использовании последних в значительной мере основаны генно-инженерные исследования, в результате которых вирус осповакцины со встроенными в него чужеродными генами становится экспрессирующим их вектором (см. ниже).
После адсорбции вирионов на клеточных мембранах и проникновения их в клетку вместе с образовавшимися эндоцитоз.- ными вакуолями происходят слияние клеточных и вирусных мембран, первичная депротеинизация вирионов и высвобождение сердцевин (нуклеоидов), которые после этой первой стадии раздевания становятся метаболически активными. Синтез ранних мРНК, образование кэп-структур и полиаденилирование обеспечиваются вирионными ферментами, при этом не происходит сплайсинга [Venkatesan S., Moss В., 1981]. Регулирующие области генов вирусов оспы имеют богатые аденином и тимином (TATAA) последовательности, но в остальном
отличаются от генов про- и эукариотов. В ранних стадиях транскрибируется 14% генома, и из ранних белков синтезируются тимидинкиназа, эндонуклеазы и другие ферменты вируса, а также «родственный белок». Таким образом, депротеинизация вириона включает две стадии: клеточно-зависимую, обеспечивающую удаление внешних оболочек, и вирусзависи- мую, обеспечивающую более полную депротеинизацию ДНК и возможность ее репликации.
Места, в которых происходят репликация ДНК и формирование вирионов, являются структурами клеточной цитоплазмы, модифицированными вирусом; их обозначают как вирусные фабрики. Более 100 белков, синтезирующихся в первые часы инфекции, обеспечивают вторую стадию раздевания, репликацию вирусной ДНК и транскрипцию поздних генов. Среди ферментов, синтезируемых в этой стадии, следует назвать ти- мидинкиназу, ДНК-полимеразу, полинуклеотидлигазу, ферменты синтеза ДНК.
Репликация вирусной ДНК связана с образованием разрывов терминальных петель, самопраймированием З'-концов; вирусной ДНК, но в общем многие детали синтеза вирусной ДНК мало изучены, и поэтому предлойсено несколько схем ее репликации. Поздняя транскрипция охватывает большую часты генома. На этой стадии транслируются структурные белки,, входящие в состав вирионов. Подсчитано, что 1A генома вирусов оспы кодирует ранние, 1A — структурные и 1A — поздние неструктурные белки [Pennington T., 1976]. В транскрипции поздних генов, по-видимому, участвует клеточная РНК-поли- мераза II. Часть белков, синтезируемых на этой стадии, подвергается протеолитическому расщеплению, которое связано; со сборкой вирионов и их предшественников.
Сборка вирионов происходит в «вирусных фабриках» и является многоступенчатым процессом. Сначала дочерняя ДНК ассоциируется с внутренними белками, образуются плотные нити и гранулы. Затем формируется оболочка вирионов в виде гладкой стенки, охватывающей электронно-плотный материал,, и образовавшиеся незрелые вирионы (провирионы) выглядят в виде пузырей, в которых формируются нуклеоиды (сердцевины) и боковые тела. Происходят уплотнение внутреннего- материала и оболочек и образование поверхностных структур, в результате чего формируются зрелые вирионы. Вирионы выводятся из клетки по каналам внутриклеточного транспорта, а часть их освобождается после гибели зараженных клеток.
Вирусы оспы вызывают глубокое угнетение синтеза клеточных макромолекул, причем эти синтезы блокируются как вирионными белками, так и белками, синтезирующимися в ходе репродукции вируса.
Некоторые представители семейства вирусов ocπt>ι вызывают пролиферацию зараженных клеток и их:неопластическую трансформацию (вирусы фибромы, Яба, контагиозный вирус моллюска). Для вирусов оспы характерны такие генетические процессы, как рекомбинация внутри родов, ■спасение маркера [Nakano Е. et al., 1982] и негенетическая ,реактивация между родами вируров оспы позвоночных. Вирусы оспы, циркулирующие в природе, подвергаются процессам рекомбинации. Одним из таких рекомбинантов является злокачественный вирус кроликов, явившийся рекомбинантом 10% ■генома вируса фибромы кроликов и 90% генома вируса миксомы кроликов [Block W. et al., 1985].
Более подробно описание особенностей генетики вирусов •оспы можно найти в монографиях и обзорах [см. Гендон Ю. 3., 1975]. Здесь же мы хотели бы отметить некоторые важные особенности, характерные, впрочем, не только для вирусов оспы, но и для других, ранее упоминавшихся ДНК-оодержа- -щих вирусов (герпесвирусы, бакуловирусы, аденовирусы и др.). Во-первых, ДНК вирусов оспы, близких между собой, или мутантов, легко рекомбинируются, образуя как гомологичные, так и гетерологичные рекомбинанты. В последнем случае образующиеся гетеро дуплексы могут включать в себя чужеродные гены. Во-вторых, рекомбинация может происходить не только между полными геномами двух вирусов оспы, но и между полным геномом и сравнительно небольшим его фрагментом (4?10δ), включенными в плазмиду, при чкотрасфекции. В-третьих, в геноме вирусов оспы имеются большие участки, ■которые условно обозначают как несущественные для репликации области генома. Они имеют размер около 25 000 пар .нуклеотидов, т. е. более 10% всего генома [Smith G., Moss. B., 1983]. По-видимому, этой областью кодируется синтез белков репликации и других, которые могут быть заменены клеточными белками, как например, тимидинкиназа, а может быть, и белки, обеспечивающие вирулентность вируса. Все эти особенности делают вирус осповакцины весьма удобным вектором для включения в него чужеродных генов. Мы не будем здесь цитировать уже многочисленную литературу, посвященную использованию вируса осповакцины для генно-инженерных операций; сошлемся лишь на меморандум экспертов ВОЗ по этому вопросу [WHO, 1985]. Важно подчеркнуть другое: при .генно-инженерных операциях, т. е. искусственных манипуля- щиях, используют естественные существующие генетические .взаимосвязи у этой и других групп вирусов.
Котати, именно эта особенность вирусов оспы послужила ^препятствием для применения ,рекомбинантного вируса осповакцины, содержащего экспрессируемый ген протективного .гликопротеида вируса бешенства, для иммунизации лисиц. При поедании мяса, смоченного суспензией такого вируса, у лисиц :и других хищников развивается осповакцинный стоматит, со
провождающийся выработкой иммунитета одновременно против осповакцины и бешенства. Однако комитет экспертов ВОЗ решительно возразил против такого проекта, намечавшегося к проведению в Аргентине. Имелась в виду опасность появления в природе рекомбинантов вирусов оспы с повышенной нейровирулецтностью, которая вызвана или связана с геном вируса бешенства, введенного в ДНК вируса осповакцины.
Вирусы оспы вызывают разнообразные заболевания, и все же наиболее выраженно они проявляются в виде высыпаний на коже и слизистых оболочках. Соответственно у животных заболевания передаются при контакте, через корм и питье, а у человека также воздушно-капельным путем. Особое место занимают неопластические процессы, поражающие либо наружные покровы, либо более глубоко лежащие ткани (фиброма). В последнем случае возможно участие в передаче кровососущих насекомых.
Громадная величина генома, сопоставимая с величиной генома у наиболее мелких бактерий (микоплазмы, риккетсии и особенно хламидии), автономность процессов репликации и транскрипции, обеспечиваемая вирусспецифическими ферментами, отсутствие сплайсинга — все это позволяет поставить одновременно два вопроса. Закономерно ли считать вирусы оспы вирусами? Не являются ли они продуктами дегенеративной приспособительной эволюции бактерий, причем даже менее далекими, нежели органеллы типа пластид и митохондрий? В самом деле, наиболее мелкие бактерии — хламидии — настолько деградировали эволюционно, что утратили собственные энергорегенерирующие системы и стали своеобразными энергетическимIj паразитами. По биологическим свойствам от вирусов оспы они-отличаются «только» наличием рибосомных белоксинтезирующих систем. Однако это «только» и является разделом между вирусами и клеточными формами жизни, поэтому пока нет веских оснований исключать вирусы оспы из царства вирусов.
К сожалению, еще не сделана проверка эволюционных соотношений вируса оспы с прокариотами (эукариотами и/или архебактериями),. так как все эти проверки, построение филогенетических древ и т. п. делались чаще всего на основе анализа рибосомных PHK (26S, 16S, 55). Последние имеются даже у потомков далеко эволюционировавших эндосимбионтов (митохондрии, пластиды). А этого как раз и нет у оспенных вирусов.
Кроме того, несмотря на большую автономию генома вирусов оспы от генома клеток «хозяев»-эукариотов, включая процессы репликации вирусной ДНК, имеются факты, свидетельствующие о генетических взаимодействиях между вирусами оспы и их «хозяевами». Так, при исследовании белка с моле-
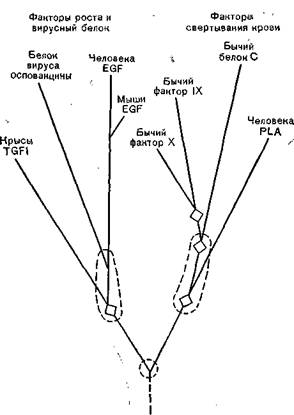
Рис. 62. Эволюционное древо белка вируса осповакцины сходных доменов у млекопитающих. Обведенные пунктиром овалы обозначают неопределенность ветвей древа.
кулярной массой 19 000, синтезируемого на ранних стадиях репродукции вируса осповакцины, было показано необычное положение цистеиновых и глициновых остатков в молекуле этого белка, сходное с положением этих остатков в двух клеточных белках — трансформирующем факторе роста (TGF)и факторе эпидермального роста (EGF).Эти два белка, обладающие сходными функциями и близкие по величине (50 и 53 аминокислотных остатков соответственно), стимулируют мутагенез. Первый обнаружен во многих опухолях и в эмбриональных тканях, второй — в подчелюстной железе и моче людей. Было высказано предположение, что все 3 белка и некоторые другие белки животных [Blomquist М. et al., 1984} имеют общее происхождение, было даже построено их эволюционное древо (рис. 62). Указанные 3 белка не имеют имму
нологического родства, но обладают сходной функциональной активностью. Белок вируса осповакцины, обладающий свойствами факторов роста клеток, является ранним секретируемым (неструктурным) белком, функции которого пока неизвестны.
C геномами эукариотических клеток геномы оспенных вирусов сближает наличие тандемных повторов в инвертированных терминальных повторах, что было показано при изучении вируса осповакцины. Предполагают, что они облегчают циклизацию однонитевых ДНК во время репликации. Эти повторы имеют размеры 70—150 пар нуклеотидов и повторяются тандемно 13—30 раз [Wittek R., Moss В., 1980].
Таким образом, происхождение вирусов оспы остается неясным. C бактериями его сближают громадный геном, независимая от клетки репликация и отсутствие сплайсинга. Однако структура их генома сходна и со структурой генома эукариотов, а некоторые гены имеют явно эукариотическое происхождение, если не считать их результатом далеко зашедшей молекулярной конвергенции.
Несмотря на большое сходство плана строения и морфологии вирионов оспы позвоночных и оспы насекомых, наличие по крайней мере 4 ферментов с одинаковыми функциями, размножение в цитоплазме и ряд других свойств, указывающих на общность происхождения обеих групп вирусов оспы, в эволюции всей группы вирусов остается много неясного. По-видимому, более древнее происхождение имеют вирусы оспы насекомых уж по одному тому, что насекомые — гораздо более древние формы организмов, нежели теплокровные позвоночные. Вирусы оспы не обнаружены у морских беспозвоночных, и если это факт,, а не недостаток знаний, то можно предположить, что вирусы оспы появились не ранее силура, т. е. около 400 млн лет назад, или еще позже, в карбоне.
Освоение ими новой экологической ниши можно связать с развитием кровососущих насекомых, так как некоторые вирусы оспы позвоночных до сих пор могут передаваться кровососущими насекомыми. Кстати, все до сих пор известные вирусы оспы позвоночных поражают теплокровных — млекопитающих или птиц, а вирусы насекомых — как некровососущих (чешуекрылые), так и кровососущих (двукрылые), в том числе комаров-кровососов теплокровных животных. Поэтому образование вирусов оспы позвоночных следует отнести к сравнительно позднему периоду — юрскому или меловому, т. е. 120—150 млн лет назад или еще позже. Именно этим можно объяснить умеренную дивергенцию, существующую между 6 группами (родами) вирусов оспы позвоночных, вплоть до сохранения общих антигенов и еще более широких антигенных связей между вирусами одного и того же рода.
! I
Эволюция этих вирусов происходила путем заполнений i ими новых экологических ниш. Она сопровождалась затем, узкой специализацией вплоть до поражения соответствующим, вирусом только одного вида. Так появились и эволюционировали вирусы оспы буйволов, верблюдов, кроликов, мышей (группа вирусов осповакцины), узелков доильщиц, контагиозной эктимы, пустулярного стоматита коров (группа вирусов, паравакцины), оспы канареек, голубей, перепелок, ласточек,, воробьев, индюшек (группа вирусов оспы птиц), оспы овец,, коз, фибромы и миксомы кроликов, оспы свиней. Приручение- человеком животных, с одной стороны, усилило специализацию вирусов (оспа овец, коз, верблюдов), с другой — интенсифицировало распространение вирусов, в результате чего одни и те же виды домашних животных стали поражаться разными вирусами оспы. Например, коровы поражаются вирусами оспо- вакцины, коровьей оспы, пара-вакцины, узелков доильщиц; кролики — вирусами кроличьей оспы, фибромы и миксомы. Впрочем, вопрос об узкой специализации не столь уж прост. Мы говорим о
Еще по теме ГЛАВА 31. ВИРУСЫ ОСПЫ:
- Вирус оспы кур
- Вирус оспы коров
- Вирус оспы мелкого рогатого скота
- Общие сведения о вирусе папилломы человека. Классификация папилломавирусной инфекции. Пути передачи вируса.
- Глава 2 ВИРУС ИММУНОДЕФИЦИТА ЧЕЛОВЕКА
- Диагностика ветряной оспы.
- Лечение и профилактика ветряной оспы
- Параграф пятьдесят пятый. Устранение следов оспы
- Параграф шестой. Следы язв и оспы
- Семейство Артеривирусов (от греч. arteria- артерия) состоит из 1 рода - Arterivirus(от греч. arteria- артерия): вирусы артериита лошадей*, респираторного и репродуктивного синдрома свиней и др. (4 вируса).
- ХАРАКТЕРИСТИКА ЭПИДЕМИЧЕСКОГО ПРОЦЕССА ВЕТРЯНОЙ ОСПЫ СРЕДИ СТУДЕНТОВ УЧРЕЖДЕНИЯ ОБРАЗОВАНИЯ «ГОМЕЛЬСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ»
- Вирус гриппа С.
- Вирусы.
- Имеет ли она общее с классификацией биологических вирусов?
- Кто и зачем создает вирусы?
- 11.3.1.6. Энцефалит, вызванный вирусом простого герпеса
- Вирус лейкоза птиц
- Почему нельзя дать точного определения компьютерного вируса?
- Существует ли сходство между биологическими вирусами и компьютерными?
- Вирусы висны-маэди овец