ГЛАВА 18. АРЕНАВИРУСЫ
Семейство аренавирусов (Arenaviridae)объединяет четко очерченную группу вирусов. Большинство вирусов вызывает тяжелые геморрагические лихорадки, передается без кровососущих членистоногих и имеет между собой серологические связи.
Немногочисленные представители (13 вирусов) этого ■семейства объединены в один род (Arenavirus).Вирионы аренавирусов имеют размер IlO—130 нм, состоят из сердцевины (нуклеокапсида) и внешней оболочки с выступами длиной 10 нм. Внутри оболочки обнаруживаются одна или несколько рибосом клетки-хозяина — «песчинок» (arena). Вирионная PHK представлена двумя вирусспецифи- ческими фрагментами с молекулярной массой 1,1 ?106— l,6?106 (S) и 2,1 XIO6—3,2?106(L).Кроме того, в вирионах содержатся 3 фрагмента рибосомной РНК: 28S, 18S и 4—6S [Leung W. et al., 1981]. Вирионные Lи S-PHK содержат сходные последовательности длиной около 30 нуклеотидов с 3'-и б'-конца молекулы. Вирионы включают неглико- зилированный белок (63 000—72 000), образующий рибонуклеопротеид, и два гликозилированных белка GPl (54 000— 72 000) и GP2 (34 000—44 000). Их синтез кодируется S- фрагментом PHK- В составе вирионов имеются также транскриптаза и поли (У)—поли (А)-полимеразы. Особенностью транскрипции генов аренавирусов является биполярность S- фрагмента РНК: половина его, примыкающая к б'-концу, имеет негативную, а вторая половина — позитивную полярность, причем они разделены шпилькой. Поэтому с вирионной PHK транскрибируется, мРНК для белка N,а второй, неструктурный белок транскрибируется с правой половины ее комплементарной копии. Репликация происходит, как и у вирусов с негативным геномом, в два этапа: сначала синтезируется полная комплементарная нить, а затем дочерние РНК.
После адсорбции на рецепторах клеточной мембраны вирионы проникают в клетки посредством виропексиса, одновременно происходят слияние клеточных и вирусных мем
бран, а также освобождение нуклеопротеида, который начинает функционировать в цитоплазме.
Транскрибированные гены функционируют как мРНК, кодируя синтез вирусных белков. Затем осуществляются синтез вирионных дочерних РНК, сборка капсидов и формирование вирионов, покидающих клетку путем почкования.Число представителей с імейства, составляющих единственный род Arenavirus,нез мико (13). Все они в естественных условиях поражают гр дзунов, вызывая у них персистентную инфекцию. Наряду с горизонтальной передачей существует вертикальная. Хотя в эксперименте показано размножение аренавирусов в клещах, аренавирусы распространяются без переносчиков (табл. 14). По крайней мере 4 представителя этого семейства вызывают тяжелейшие забо-. левания, протекающие обычно с геморрагическим синдромом: лимфоцитарный хориоменингит (ЛХМ), лихорадки Ласса, Мачупо и Хунин. Все аренавирусы являются возбудителями персистирующих, частично бессимптомно протекающих инфекций грызунов. Лишь вирус Такарибе был, помимо грызунов, выделен от летучих .мышей и комаров. Возможно, эти лихорадки случайны [Murphy F., 1977].
Эндемичные очаги в настоящее время расположены в экваториальном, субэкваториальном, тропическом и субтропическом климатических поясах. Исключение составляет лимфоцитарный хориоменингит (см. ниже). Аренавирусы широко распространены в природе как среди диких, так и полуси- нантропных грызунов (луговые формы). Именно последние являются причиной вспышек заболеваний среди людей. Это происходит в результате сезонных миграций грызунов в жилье человека (лихорадка Ласса, боливийская геморрагическая лихорадка) или во время сельскохозяйственных работ (сбор маиса в очагах аргентинской геморрагической лихорадки). Заболеваемость носит сезонный характер. После ухода грызунов или их истребления заболевания прекращаются. Среди грызунов вирус распространяется как горизонтальным, таки вертикальным путем [Rawls W., Leung W., 1978]. Люди заражаются через пищу, воду, воздух, поврежденные участки кожи, загрязненные выделениями грызунов (главным образом мочой).
Формирование аренавирусов, вероятно, «закладывалось» в палеоцене, когда существовала связь между Южной Америкой и Африкой.
В экспериментах показана высокая частота рекомбинаций с ^-мутантами вируса Пичинде. Нельзя исключить роль рекомбинации в эволюции, хотя возможность репликации двух разных аренавирусов в организме одного грызуна мала. Следует учесть достаточную узкую приуроченность различных аренавирусов к отдельным видам грызунов.Таблица 14, Распространение аренавирусов и их связь с грызунами
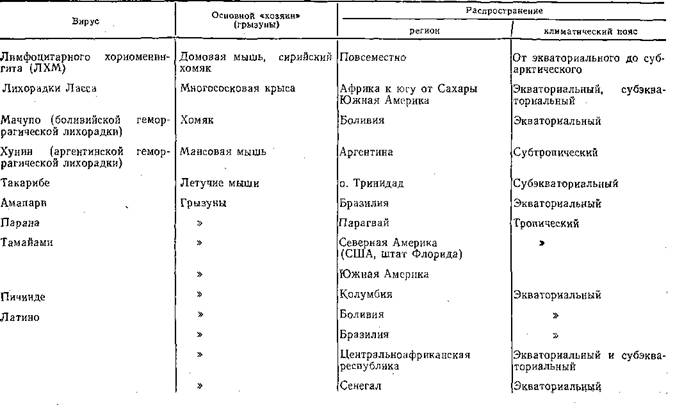
В Южной Америке аренавирусы экологически связаны с одной трибой семейства хомякообразных (Cricetidae), попавших в Южную Америку из Северной Америки в течение миоцена — плиоцена. Этот процесс сопровождался активным образованием видов. Возможно, с этим периодом связано формирование и южноамериканских видов аренавирусов [Ca- Iosher C., 1980]. Появление аренавируса Тамайами в Северной Америке (Флорида), по-видимому, носит вторичный характер. Вирус паразитирует в Sigmodon hispidus,имеющих южноамериканское происхождение в плейстоцене с вторичным проникновением через Центральную Америку в южные части Северной Америки [Arata A., Gratz M., 1975].
Основным природным резервуаром африканского аренавируса Ласса является многососковая крыса Praomys (mas- tomys) natalensis,таксономически занимающая промежуточное положение между домовой мышью и корабельной (черной) александрийской крысой. Грызуны обитают в ,зоне саванн к югу от Сахары. На африканском континенте это наиболее распространенный и многочисленный вид, населяющий кустарники и обработанные поля, легко перемещающийся от жилья человека к местам обитания диких грызунов. Вид ведет свое происхождение со времен плейстоцена и возник в ходе последовательных периодов изоляции в зонах перехода саванн в леса [Bellier, 1975]. В Старом Свете аренавирусы связаны с одним из 6- подсемейств мышинообразных Muridae.Обе указанные группы грызунов Старого и Нового Света являются наиболее ,многочисленными и жизнеспособными, в пределах своих ареалов по численности они преобладают над другими грызунами [Arata A., Gratz M., 1975].
Вероятно, существуют длительные экологические связи между южноамериканскими аренавирусами и группой близкородственных грызунов из родов Oryzomus, Calomus, Akodon, Neacomys, Thomasomys, Sigmodon.Видообразование вируса ЛХМ связано, вероятно, со становлением человеческого общества и повсеместным распространением домовых мышей. Этот вид мышей до начала 70-х годов считался единствённым резервуаром вируса. Однако в последние десятилетия описан ряд вспышек заболевания людей, заразившихся от содержавшихся в домашних условиях сирийских хомяков, переболевших ЛХМ чаще в виде персистирующей инфекции, иногда клинически выраженной. Природные очаги ЛХМ практически не изучены. Известны случаи выделения вируса от полевок Apodemus Sylvaticusи A. fIavicollis [Lehmnnn-Grube F. е. а., 1971]. Род Apodemus таксономически близок к роду Musи относится к палеарктическим формам. Первоначальные ареалы этих родов частично совпадают. Прародиной домовых мышей является
Центральная Азия, где, вероятно, ,и следует искать сохранившиеся в настоящее время природные очаги ЛХМ. Из центра формирования вид распространился в двух направлениях — в северно-западном (Al. musculus musculus)и юго-западном. (М. musculus domesticus).Эти популяции в настоящее время не смешиваются, но обе являются носителями ЛХМ.. Первичное их инфицирование, вероятно, произошло в центре видообразования — на юге Среднеазиатского региона.
Еще по теме ГЛАВА 18. АРЕНАВИРУСЫ:
- Глава 17 ВЫЧИСЛИТЕЛЬНЫЙ ЭКСПЕРИМЕНТ
- Система семейной поддержки (глава 19)
- Глава 1. Введение в иммунологию
- Глава 7. Этапы факоэмульсификации
- Встреча с Внутренним Наставником (глава 15)
- Глава 8. Болезни легких
- ГЛАВА 12 АНОМАЛИИ И ЗАБОЛЕВАНИЯ РОГОВИЦЫ
- ГЛАВА 6. ЭПИДЕМИОЛОГИЯ И ПРИНЦИПЫ ОРГАНИЗАЦИИ ПСИХИАТРИЧЕСКОЙ ПОМОЩИ
- Глава 6 Сопроводительная терапия в детской онкологии и гематологии
- ГЛАВА 6 ПАЛЬЦЕВАЯ ДЕРМАТОГЛИФИКА ПРИ ОГРАНИЧЕННЫХ ФИЗИЧЕСКИХ ВОЗМОЖНОСТЯХ
- Глава 3 КЛИНИКА, ДИАГНОСТИКА И ПРОГНОЗ АНДРОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ У ДЕТЕЙ И ПОДРОСТКОВ
- Глава З Патологические состояния сетчатой оболочки, выявляемые при ультразвуковой диагностике.
- Глава 1
- Глава 1
- Глава 2
- Глава 3
- Глава 4