ГЛАВА 19. РЕОВИРУСЫ И СХОДНЫЕ ГРУППЫ ВИРУСОВ
Помимо обширной группы реовирусов (семейство Reo- viridae),двунитевую PHK имеют многие другие группы вирусов: вирусы дрожжей и плесневых грибов с линейной непрерывной PHK (10 вирусов); вирусы грибов с двумя сегментами PHK — бипартитные (около 20 вирусов), трехсегментные— трипартитные (4 вируса), монопартитные бисег- ментные вирусы грибов (4 вируса), монопартитные бисег- ментные вирусы животных (4 вируса).
К ним надо добавить, дрожжевые киллеры, представляющие собой двунитевую PHK без белковых оболочек [Matthews R., 1982]. О названных 5 группах вирусов мы упомянем лишь кратко, так как сведения о них недостаточны для того, чтобы строить какие- либо представления об их эволюции.Вирусы первой группы имеют геном с молекулярной массой 3?106—5,7?106. Вирионы представляют собой полиэдры диаметром 35—40 нм, построенные из белковых молекул с молекулярной массой 73 000—160 000. Выделяют две подгруппы (рода): Helminthosperium maydisи Saccharomyces сег- υiridae.
Вирусы второй группы обнаружены в плесневых и других грибах. Их геном состоит из двух моноцистронных фрагментов двунитевой PHK с молекулярной массой 0,9?106— 1,6?106, которые заключены в разные частицы (бипартитный вирус). Кроме того, встречаются дефектные частицы с неполными фрагментами генома. PHK упакована в белки с молекулярной массой 42 000—72 000, частицы имеют форму икосаэдра диаметром 30—35 нм. Вторым белком является полимераза с молекулярной массой 56 000. У более изученного вируса этой группы (вирус Penicillium Solaniticumгруппы Aslz-S) имеется несколько видов частиц с разной плотностью— от 1,3 до 1,39 г/мл, зависящей от разного содержания в них РНК, включая пустые капсиды. По совокупности свойств среди бипартитных вируоов грибов выделяют около 10 групп в ранге видов. Все они, как и следовало
ожидать, кроме капсидного белка, имеют вирионную полимеразу.
Эти две группы вирусов недавно выделены в самостоятельные семейства — Totiviridaeи Partitiviridae [Brown F., 1986].Вирусы следующей группы имеют геном в виде двуните- вой РНК, являются трипартитными вирусами и обнаружены также у плесневых грибов. Сегменты моноцистронные, их молекулярная масса около 2?106, они упакованы в разные частицы икосаэдрической формы диаметром 35—40 нм, образованные белком с молекулярной массой 110 000—120 000. В частицах содержится также вирусная полимераза.
Отдельную группу образуют двухсегментные монопар- титные вирусы животных, содержащие двунитевую РНК. Вирусы поражают широкий круг «хозяев» (рыб, моллюсков, кур, уток, индюшек, дрозофилу). Один из наиболее изученных вирусов этой группы — вирус инфекционного панкреатического некроза лососей — морфологически выглядит в виде икосаэдра диаметром 60 нм, имеет плотность 1,32 г/мл. Его геном состоит из двух фрагментов с молекулярной массой 2,5?106и 2,3?106. PHK неинфекционна, а в вирионах содержится вирусспецифическая полимераза. В составе капсида имеются 4 белка с молекулярной массой 105000, 54 000, 31000 и 29 000 в разных соотношениях (соответственно 22, 544, 550 и 122 молекулы на вирион). Кроме того, в ходе репликации синтезируются невирионные белки, созревание структурных белков происходит путем протеолитического расщепления предшественников. Несмотря на широкий круг «хозяев»/, эти вирусы образуют компактную группу, имеют исходную характеристику нуклеоновых кислот и белков [Dobos P. et al., 1979]. Некоторые из них серологически родственны. Предлагается выделить их в самостоятельную группу Birnavirus,у вирусов которой капсид состоит из 132 морфологических единиц с кубической симметрией и Т-13 [Schwanz-Pfitzner I. et al., 1984]. Недавно их выделили в самостоятельное семейство Birnaviridaeс единственным одноименным родом [Brown F., 1986].
К бирнавирусам относят вирус Xдрозофилы. Его двуни- тевая PHK состоит из двух сегментов — А и В(2,3?106и 2,2?106).
Вирионы содержат 5 белков: большие (110000), средние (49 000 и 45 000) и малые (34 000 и 27 000). В зараженных клетках синтезируются 5 неструктурных белков (110 000, 67 000, 34 000 и 27 000), а также белок молекулярной массой 49 000, являющийся продуктом расщепления белка с массой 67 000. Стратегия генома этого вируса, таким образом, отлична от стратегии генома других вирусов с дву- нитевой PHK [Nagy E., Dobos P., 1984].Таким образом, упомянутые выше 4 группы вирусов, несмотря на их явную разобщенность, имеют много общего. В частности, стратегия их генома, по-видимому, весьма сходна, так как их PHK неинфекционна и функционирует лишь при наличии вирусной полимеразы, присутствующей либо в вирионах, либо в частицах дву- и трипартитных вирусов. Как будет показано ниже, это сближает их с рео- вирусами. Быть может, к ним надо добавить еще одного представителя—Фаг Ф6, поражающий псевдомонад. Его геном составляют три фрагмента двунитевой PHK с общей молекулярной массой 10,4?106. Молекулярная масса фрагментов 2,3?106; 3,1 XlO6и 5,0? IO6. Вирусная PHK кодирует синтез 11 белков, включая транскриптазу (молекулярная масса их от 6000 до 82 000). Вирус имеет капсид кубической симметрии (диаметр 60 нм) и липидную внешнюю оболочку (диаметр вирионов 75 нм) [Day L., Minilich L., 1980]. Вирус выделен в отдельное семейство Cystoviridae.
Дрожжевые киллеры также состоят из двунитевой РНК- Они, по-видимому, не имеют собственных белков. В этом отношении они больше напоминают плазмиды, хотя геномом их является не ДНК, а РНК. В их геноме находится ген для синтеза токсина, являющегося кислым белком с молекулярной массой 10 000—20 000, который убивает дрожжи без киллеров. По механизму действия — образование каналов в мембранах, через которые выходят ионы, — киллерный токсин напоминает бактерицины, хотя химически это разные вещества, различна и их молекулярная масса, и структура, не говоря уж о том, что колицины являются ДНК-содержа- щими плазмидами [Kagan В., 1983].
Семейство Reoviridaeвключает роды Reovirus(собственно реовирусы), Orbivirus(вирусы передаются кровососущими членистоногими), Rotavirus(возбудители кишечных заболеваний), Phytoreovirus(возбудители тканевых опухолей растений), Fijivirus(растительные реовирусы второй подгруппы), Cynovirus(возбудители цитоплазматических полиэдрозов). Все они являются вирусами с. двунитевой мульти- сегментной РНК.
Вирионы имеют сферическую форму, диаметр 75 нм, состоят из двухслойного капсида, внутри которого находится вирионная PHK- Внешний капсид является икосаэдром с 12 вершинами, число капсомеров точно не определено (от 92 до 180). Они объединены в гексоны и пентоны и представлены в виде цилиндров диаметром 18—20 нм с каналом диаметром 4—6 нм. Внутренний капсид, или сердцевина, имеет диаметр 52 нм, является также икосаэдром с 12 вершинами. Геном состоит из 10 фрагментов двунитевой PHK с общей молекулярной массой около 16?106. Соответ -
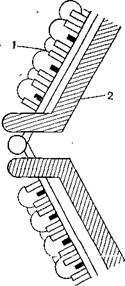
Рис. 29. Строение капсида реовируса (схема).
1 — наружный капсид; 2 — внутренний капсид.
ственно по размеру выделяют 3 класса молекул РНК: большие (Ll—L3)с молекулярной массой 2,7?106и 4500 пар нуклеотидов, средние (Ml—М3) с молекулярной массой 1,3XIO6и 2300 пар нуклеотидов, малые (Sl—S4)с молекулярной массой 0,6?106— 0,8?106и 1200 пар нуклеотидов. Каждый сегмент кодирует синтез одного структурного или неструктурного белка. Синтез белков сердцевины кодируется сегментами Llи L3.Белки обозначаются соответственно λl и λ3, а также сегментами Mlи М2 (μl и μ2) и S2 (σ2). Синтез белков внешнего капсида кодируется сегментами М2 (μlc), Slи S3 (σl, σ3). В вирионе внешний капсид образуют не только' ее белки, но также выступающие внутрен
ние белки (рис. 29). Белок σι является гемагглютинином. В ходе репликации образуются два неструктурных белка, кодируемые генами М3 (μNS)и S3 (σNS), с молекулярной массой 75 000 и 36 000.
Молекулярная масса белков λ составляет соответственно 155 000, 140 000 и 135 000, белков μ -80 000, 72 000 и70 000, белков σ — 42 000, 38 000 и 34 000.
Напомним вкратце характеристику основных групп (родов) этого обширного семейства.
Реовирусы (более 10 вирусов) поражают человека,, обезьян, крупный и мелкий рогатый скот, летучих мышей,, птиц, т. е. только теплокровных животных. Геном их состоит из 10 фрагментов двунитевой РНК, упакованных в двуслойный капсид икосаэдральной формы диаметром 60—76 нм с 12 полыми выступами на поверхности. Общая молекулярная масса генома 14?106—15?106, масса его отдельных фрагментов варьирует в пределах 0,2?106— 2,7?106. PHK имеет большие (L),средние (M)и малые, а также короткие- цепи, состоящие из 2—20 нуклеотидо’в. Плюс-нить вирионной PHK на 5'-конце содержит кэп-структуру m7GpppGmpCp [Урываев Л. В., 1982]. Соответственно обозначают и кодируемые ими белки λ, μ и σ. В составе вирионов имеется 10 полипептидов, включая транскриптазу, нуклеотидфосфогид- ралазу >и кэппинг-фермент. Серологические типы вируса перекрестно реагируют. Для реовирусов, включая орбивирусы, характерна консервативность З'-концевых последовательно
стей как разных генов, так и генов различных вирусов, относящихся к одному 'и тому же роду [Mertens P., Saugar D., 1985]. Другие роды в общем имеют сходные характеристики и некоторые отличия.
Большая группа (род) орбивирусов, насчитывающая более 100 представителей, подразделяется на 13 серологических групп, из которых наиболее многочисленны группы вирусов синего языка овец, Кемерово, Чангвинола, африканской болезни лошадей [Spence R. et al., 1985]. Тотальный геном орбивирусов, состоящий из 10 фрагментов, у вируса синего языка имеет молекулярную массу 14,9?106. Геном вируса лихорадки Колорадо содержит 12 сегментов с общей молекулярной массой 18?106. Сегменты генома имеют сходные последовательности на З'-концах, являясь, по-видимому, сайтами распознавания для вирусной транскриптазы.
Благодаря фрагментарности генома в естественных условиях и эксперименте образуют реассортанты, что показано на вирусах синего языка овец. Четыре мажорных и 3 минорных белка орбивирусов по размерам распределяются примерно так же, как у реовирусов. Всего же в ходе репликации образуется 12 белков с молекулярной массой от 20000 до 141 000.Группа (род) ротавирусов поражает человека, обезьян, крупный и мелкий рогатый скот, свиней, лошадей, диких млекопитающих (антилоп, оленей, грызунов, и пр.), собак, ,птиц. Общее число ротавирусов трудно определить, так как их разные серовары сходны у человека и животных. Ротавирусы — возбудители кишечных инфекций, возможно бессимптомное носительство. Геном ротавирусов состоит из 11 фрагментов с молекулярной массой от 0,2?106до 2,2? XlO6, общая молекулярная масса 10?IO6—-12? 106. В составе вирионов находятся 8—10 полипептидов. Вирионы имеют форму икосаэдров диаметром 65—75 нм. При смешанной инфекции двумя близкими ротавирусами возникают реассортанты, причем не только в культурах тканей, но и при заражении животных. Они являются источниками для селекции, проявляющейся по-разному в различных условиях [Gombold J., Ramig R., 1986]. У ротавирусов пересортировка генов происходит особенно легко. В частности, удается получить реассортанты между хорошо культивируемыми ротавирусами животных (например, коров) и «некультивируе- мыми» ротавирусами человека. Этот подход может быть использован для получения живых, аттенуированных для человека вакцин [Midthun К. et al., 1985]. Вполне естественно предположить, что эти процессы происходят в природе, и, может быть, в этом заключается причина обширных перекрестных антигенных связей между ротавирусами челове
ка и животных. Так, показано [Allen A., Dosselberger U., 1985], что ротавирусы, выделенные от детей с иммунодефицитом, хронически .инфицированных, являются реассортанта- ми между ротавирусами человека и коровы.
Серологические взаимоотношения между ротавирусами довольно сложны. Например, ротавирусы, выделенные от кошек, серологически близки к ротавирусам обезьян (SAll)и человека (тип 3), но не других сероваров [Birch С. et al., 1985]. Прикрепление к клетке и выработка вирус - нейтрализующих антител связаны с гликопротеидом внешней оболочки с молекулярной массой 38 OOQ — 41900 [Sahara М. et al., 1985].
Помимо этих групп (родов) вирусов, имеются неклассифицированные ротавирусы животных, например, синцитиальный вирус кроликов.
Среди обширной группы реовирусов растений выделяют роды Phyloreovirus(первая подгруппа растительных реовирусов), Fijivirus(вторая подгруппа), в каждую из которых входит несколько вирусов. Первый род включает более 30- вирусов раневых опухолей растений, а также вирусы карликовости риса. Геном их состоит из 12 фрагментов с молекулярной массой от 0,3?106до 3?106; общая молекулярная масса 16?106. В вирионах содержится 7 полипептидов. Вирионы представляют собой икосаэдры диаметром около 70 нм,. имеют двуслойную оболочку с сердцевиной диаметром 59 нм. Вирусы передаются тлями, размножаются как в растениях,, так и тлях. Род фидживирусов включает многие сотни сероваров, сгруппированных в группу вирусов карликовости маиса (около 400 разновидностей), дающих перекрестные серологические реакции, группу вирусов болезни Фиджи (119 разновидностей) и группу вирусов стерильной карликовости маиса (около 220 разновидностей). Геном состоит из 10 фрагментов с молекулярной массой от IO6до 2,9?106; общая молекулярная масса 18?106— 20?106. Впрочем,, некоторые авторы приводят меньшие цифры. Например, общая молекулярная масса генома вируса rice ragged stunt составляет 11,63XlO6 [Boccardo G., Milne R., 1980]. В вирионах содержится 7 белков, вирионы являются икосаэдрами диаметром 65—71 нм с12 выступами 5-кратной симметрии. Оболочка двуслойная, диаметр сердцевины 50—55 нм. Передаются тлями.
К реовирусам относятся также многочисленные возбудители цитоплазматических полиэдрозов (две группы). Первая группа включает 12 вирусов, типичным из которых является вирус цитоплазматического полиэдроза тутового шелкопряда. Его геном состоит из 10 фрагментов с молекулярной массой от 0,3?106до 2,7?106; общая молекулярная масса
около 15?106 [Payne C., Rivers C., 1976]. В вирионах содержится до 5 пептидов, а также транскриптаза, нуклеотидфосфогидролаза, кэппинг-фермент. Вирионы имеют диаметр 50—65 нм и 12 выступов. Вирусы поражают чешуе- и двукрылых, .других насекомых, а также ракообразных.
Циловирусы имеют геном, состоящий из 10 сегментов с общей молекулярной массой 13?106—16?106. Вирионы имеют диаметр 50—65 нм, 12 трубчатых выступов размещены на вершинах икосаэдра, внешний капсид не имеет этой структуры. Вирионы реовирусов всех родов имеют в своем составе РНК-зависимую РНК-полимер азу (транскриптазу), ферменты формирования кэп-структур (включая их метилирование).
Другая группа содержит около 150 недостаточно классифицированных вирусов, поражающих тлей, цикаделлид. Некоторые из этих вирусов могут также размножаться в растениях. Геном их состоит из 8 фрагментов с молекулярной массой от 0,5?106до 2,5?106; общая молекулярная масса около 12?106.
После адсорбции на клеточных рецепторах вирионы проникают в клетки посредством виропексиса (эндоцитоза), причем в этом процессе решающая роль принадлежит белку σl (гемагглютинину). При этом происходит частичная депротеинизация проникших вирионов клеточными протеазами, в результате чего внешний капсид удаляется и обнажается сердцевина. Вслед за этим осуществляется синтез мРНК с помощью вирионной транскриптазы (продукт λ3 гена Llи продукт μlcгена М2).Причем в первую очередь синтезируются и подвергаются процессингу (образование кэп-структур) мРНК, кодирующие синтез белков λl—3,62 и μNS.Вновь синтезированные PHK выходят из сердцевины через отверстия в выступах. Вслед за этим происходят трансляция и репликация генома, а затем транскрипция и трансляция поздних генов. Разные гены транскрибируются с разной скоростью: так, сегмент М2(кодирующий синтез белка μl) транскрибируется в 6 раз быстрее и транслируется в 30 раз интенсивнее, нежели сегмент Llи его продукт λ3. Сборка вирионов осуществляется в цитоплазме, вирионы покидают зараженные клетки после их лизиса.
Переходя к обсуждению возможных путей эволюции вирусов с геномом в виде двунитевой РНК, можно высказать следующие соображения. Прообразом этих вирусов можно считать дрожжевые киллеры, которые по своим свойствам сходны с плазмидами. Они представляют собой «голые» молекулы РНК, двунитевидность которых служит хорошей защитой от клеточных нуклеаз («голая» однонитевая PHK вряд ли. может сохраниться в цитоплазме). Размножение их
происходит с участием клеточных РНК-синтезирующих' систем, которые существуют у низших эукариотов (грибов), •поскольку собственных систем синтеза PHK эти структуры не имеют. Другие вирусы грибов с двунитевой PHK (моно-, би- и трипартитные), могут служить примером дальнейшей эволюции. Они имеют уже два «приобретения», делающие их классическими вирусами: собственный белок капсида и собственную РНК-репликазу — транскриптазу. Это весьма существенное «приобретение», позволившее вирусам с двунитевой PHK занять новые экологические ниши — не только древних по происхождению эукариотов (грибы), но и высших животных (бирнавирусы). Фрагментарность генома и мультипартитность имеют свои преимущества и свои недостатки и поэтому это могло явиться альтернативой в эволюции вирусов данной категории. В частности, по мере увеличения размеров генома он подразделялся на фрагменты, каждый из которых являлся отдельным геном, конечно, гораздо более устойчивым к действию внешних факторов, чем громадный геном с молекулярной массой 15?106— 20?10s. Не следует забывать, что такой геном не мог бы эффективно реплицироваться, поскольку величина 7?106—8?106 (правда, этот расчет сделан для однонитевой РНК) является предельным размером РНК.
Несмотря на сходство сравниваемых вирусов, между ними существуют и определенные отличия в стратегии генома. Дрожжевой вирус, имеющий ассоциированную с капсидом полимеразу, реплицируется консервативно, таким же путем осуществляется и транскрипция. У фага Ф6 и у других вирусов грибов транскрипция происходит полуконсервативно. У реовирусов и транскрипция, и репликация осуществляются консервативно [Nemeroff M., BruennJ., 1986].
Итак, собственный капсидный белок и вирионная полимераза явились весьма ранним «приобретением», которое, быть может, возникло около миллиарда лет назад. Не случайно поэтому среди вирусов с двунитевой PHK имеются примитивные представители (один капсидный белок — одна полимераза, иными словами, минимальное число генов), паразитирующие на наиболее древних эукариотах — грибах. Мы не знаем, вероятно, промежуточных групп, так как рео- вирусы — значительно более поздние образования, паразитирующие на членистоногих, теплокровных животных и растениях. Все это; могло возникнуть не ранее 300 млн лет назад, а скорее всего значительно позже, так как, несмотря на широкий круг «хозяев», они на редкость сходны и не слишком разнообразны.
Реовирусы служат хорошей- иллюстрацией извилистых и на сегодня мало понятных путей эволюции вирусов. Харак
терной особенностью является наличие в качестве генома фрагментарной двунитевой РНК, двуслойного капсида и вирионной РНК-зависимой РНК-полимеразы. Молекулярная масса генома, состоящего из 10—12 фрагментов, колеблется в пределах 10?106— 18? IO6, каждый ген кодирует синтез одного соответствующего вирусспецифического белка. Вирусы репродуцируются в цитоплазме, причем транскрипция и репликация их генома обеспечиваются собственным фрагментом.
Экологически эти вирусы образуют 5 групп. Собственно реовирусы паразитируют в клетках дыхательных путей и пищеварительного тракта человека, обезьян, крупного и мелкого рогатого скота, собак и птиц (около 20 вирусов). Другой род — ротавирусы — включает возбудителей диареи детей, болезни телят Небраски, диареи мышей и других животных [Дроздов С. Г. и др., 1982]. Третий род — орби- вирусы — являются типичными арбовирусами, передаваемыми комарами, мокрецами и клещами (около 15 вирусов). Среди них встречаются вирусы, имеющие большое значение для ветеринарии (вирусы синего языка овец, африканской болезни лошадей, клещевой лихорадки Колорадо, геморрагической болезни оленей). Четвертую группу составляют возбудители цитоплазматических полиэдрозов шелкопряда и других насекомых (около 150 вирусов). И, наконец, в пятую группу входят вирусы карликовости риса и маиса, болезни Фиджи и раневых опухолей растений (около 10 вирусов), передающиеся насекомыми (тлями).
Поразительное сходство геномов у всех 5 родов вирусов (по молекулярной массе и числу фрагментов), наличие в вирионах уникального фермента РНК-зависимой РНК-поли- меразы и в то же время отсутствие антигенных связей между вирусами разных групп позволяют заключить, что эволюция их как изолированных групп происходила в течение длительного времени. Вероятно, наиболее древней является группа вирусов цитоплазматических полиэдрозов. Это почти симбиотические вирусы, вызывающие не острые патологические процессы, а скорее хроническую персистенцию у чешуе- и двукрылых. Вирионы инкапсидированы в капсулообразующие белки. Обилие вирусов и поражаемых ими видов насекомых, а также описанный выше характер инфекционного процесса свидетельствуют о своеобразном экологическом - равновесии, установившемся между паразитом и «хозяином». Можно предположить, что эти вирусы или их предшественники явились источниками двух ветвей эволюции — реови- русов растений и орбивирусов. В обоих случаях насекомые (членистоногие) сохранились в качестве переносчиков, причем не механических, так как растительные вирусы размно
жаются в -организме тлей, а орбивирусы— в организме насекомых и клещей. Две остальные группы — собственно реовирусы и ротавирусы — имеют, вероятно, более позднее происхождение, и потеря переносчика является вторичной. Конечно, такое заключение сугубо гипотетическое, и предстоит объяснить, когда и почему представители разных эволюционных ветвей потеряли антигены. Для этого, в частности, следует изучать серологические взаимоотношения между ■ многочисленными вирусами цитоплазматических полиэдрозов насекомых, растительных рео- и орбивирусов.
Орбивирусные инфекции являются природно-очаговыми, как, например, эпизоотическая геморрагическая болезнь оленей, передаваемая мокрецами; вполне возможно, что олени являются вторичными «резервуарами» — индикаторами природных очагов этого орбивируса. К природно-очаговым инфекциям относится и клещевая лихорадка Колорадо. Некоторые орбивирусы пока ничем себя -не проявили, будучи выделенными из переносчиков (вирусы Кемерово, Охотский). В то же время имеются вполне сформировавшиеся зоонозы домашних животных, к которым относятся болезнь синего языка овец, африканская чума лошадей, болезнь Ибараки крупного рогатого скота. Болезнь синего языка, передаваемая мокрецами (Culicoides),вначале была распространена в Южной Африке, но затем распространилась на север Африки, Ближний Восток, юг Европы и в странах Америки. Известны 16 сероваров вирусов. Распространение болезни далеко за пределами первоначального ареала свидетельствует об «отрыве» ее от природного очага, что связано с повсеместным распространением переносчиков — мокрецов и участием в циркуляции вируса комаров (Aedes),кровососок (Malophagus), а также птиц, заносящих вирус в новые местности [Сюрин В. H., Фомина H. В., 1979]. Сформировавшимся зоонозом домашних животных является также африканская чума лошадей, передающаяся мокрецами и комарами. Известны 9 сероваров вируса. Болезнь из Южной Африки распространилась в страны Ближнего и Среднего Востока и на юг Европы. Это также относится к болезни Ибараки крупного рогатого скота, распространенной в Японии, Индонезии и на о. Тайвань. Ни один из этих вирусов не стал патогенным для человека.
Собственно реовирусы и ротавирусы являются, по-видимому, результатом далеко зашедшей эволюции. Реовирусы серовара 3наряду с человеком поражают широкий круг диких и домашних животных. Все эти вирусы вызывают заболевания в первую очередь кишечника, а также дыхательных путей (отсюда и название вирусов Respiratory Enteric Orphans),обычно доброкачественные, с тенденцией к бес
симптомному носительству. Отсутствие клинически выраженной картины заболевания, полиморфный характер патогенеза и антропозоонозный характер реовирусных заболеваний человека— все это создает впечатление, что реовирусы продолжают интенсивно эволюционировать. C этой точки зрения реовирусные болезни птиц носят более определенный характер. У кур реовирусы вызывают развитие тендосиновита и бурсита, у индюшат — синдром синего гребня, у уток—■септический процесс. Все эти заболевания вызываются разными реовирусами, непатогенными для человека. Заражение происходит через корм и воду [Сюрин В. H., Фомина H. В., 1979].
Еще по теме ГЛАВА 19. РЕОВИРУСЫ И СХОДНЫЕ ГРУППЫ ВИРУСОВ:
- Семейство Реовирусов (Family Reoviridae)
- 2 группа (беременные с клиническими проявлениями ВПЧ инфекции в виде аногенитальных бородавок, ассоциированных с наличием 6 и 11 типов вируса).
- 2 группа (беременные с клиническими проявлениями ВПЧ инфекции в виде аногенитальных бородавок, ассоциированных с наличием 6 и 11 типов вируса).
- 2 группа (беременные с клиническими проявлениями ВПЧ инфекции в виде аногенитальных бородавок, ассоциированных с наличием 6 и 11 типов вируса).
- группа (беременные с клиническими проявлениями ВПЧ инфекции в виде аногенитальных бородавок, ассоциированных с наличием 6 и 11 типов вируса).
- группа (беременные с клиническими проявлениями ВПЧ инфекции в виде аногенитальных бородавок, ассоциированных с наличием 6 и 11 типов вируса).
- 2 группа (беременные с клиническими проявлениями ВПЧ инфекции в виде аногенитальных бородавок, ассоциированных с наличием 6 и 11 типов вируса).
- 2 группа (беременные с клиническими проявлениями ВПЧ инфекции в виде аногенитальных бородавок, ассоциированных с наличием 6 и 11 типов вируса).
- 2 группа (беременные с клиническими проявлениями ВПЧ инфекции в виде аногенитальных бородавок, ассоциированных с наличием 6 и 11 типов вируса).
- Общие сведения о вирусе папилломы человека. Классификация папилломавирусной инфекции. Пути передачи вируса.
- Глава 2 ВИРУС ИММУНОДЕФИЦИТА ЧЕЛОВЕКА
- Параграф второй. Расхождение стенок и то, что с этим сходно
- Параграф двенадцатый. Мази для переломов и того, что с ними сходно
- Параграф пятьдесят восьмой. Признаки по качеству прыщей и того, что с ними сходно