оЮРНЦЕМЕГ
б ОЮРНЦЕМЕГЕ НОСУНКЕБНЦН ПНЯРЮ ПЮГКХВЮЧР РПХ ЩРЮОЮ: РПЮМЯТНПЛЮЖХЧ ГДНПНБНИ ЙКЕРЙХ Б НОСУНКЕБСЧ (ХМХЖХЮЖХЪ), ОПНЛНЖХЧ ("ОНДЯРПЕЙЮРЕКЭЯРБН") Х ОПНЦПЕЯЯХЧ НОСУНКХ.
рПЮМЯТНПЛЮЖХЪ ГЮЙКЧВЮЕРЯЪ Б ОПХНАПЕРЕМХХ ХЯУНДМНИ ГДНПНБНИ ЙКЕРЙНИ ЯОНЯНАМНЯРХ АЕЯОПЕДЕКЭМН ПЮГЛМНФЮРЭЯЪ Х ОЕПЕДЮВЕ ЕЕ ДНВЕПМХЛ ЙКЕРЙЮЛ ОН МЮЯКЕДЯРБС.
рПЮМЯТНПЛЮЖХЪ ЛНФЕР ОПНХЯУНДХРЭ, ОН-БХДХЛНЛС, ДБСЛЪ ОСРЪЛХ - ЛСРЮЖХНММШЛ Х ЩОХЦЕМНЛМШЛ. нАЮ ОСРХ ОПЕДЯРЮБКЪЧР ЛЕУЮМХГЛ МЮПСЬЕМХЪ ПЕЦСКЪЖХХ ЙКЕРНВМНЦН ДЕКЕМХЪ. оНЩРНЛС ОНМХЛЮМХЕ ЛЕУЮМХГЛНБ ЙЮМЖЕПНЦЕМЕГЮ ОПЪЛН ЯБЪГЮМН Я ЖЕМРПЮКЭМНИ ОПНАКЕЛНИ ЯНБПЕЛЕММНИ АХНКНЦХХ ЙКЕРЙХ - ЯСЫМНЯРЭЧ ЙКЕРНВМНЦН ДЕКЕМХЪ Х ПЕЦСКЪЖХХ ЩРНЦН ОПНЖЕЯЯЮ.бЕДСЫХЛ АХНУХЛХВЕЯЙХЛ ОПНЖЕЯЯНЛ, НАЕЯОЕВХБЮЧЫХЛ ЙКЕРНВМНЕ ДЕКЕМХЕ, ЪБКЪЕРЯЪ ПЕОКХЙЮЖХЪ дмй БЯЕЦН ЦЕМНЛЮ ЙКЕРЙХ Б ТЮГЕ "S" ЛХРНРХВЕЯЙНЦН ЖХЙКЮ. щРНР ОПНЖЕЯЯ НЯСЫЕЯРБКЪЕРЯЪ ЛСКЭРХТЕПЛЕМРМШЛ ЙНЛОКЕЙЯНЛ Х МЮВХМЮЕРЯЪ Я ОНЪБКЕМХЕЛ Б ЙКЕРЙЕ Б ТЮГЕ Gl ЯОЕЖХЮКЭМНЦН ХМХЖХЮРНПЮ ЙКЕРНВМНЦН ДЕКЕМХЪ. хМХЖХЮЖХЪ ЙКЕРНВМНЦН ДЕКЕМХЪ Х МЮВЮКН ПЕДСОКХЙЮЖХХ дмй ГЮБХЯЪР НР ЯХМРЕГЮ АЕКЙЮ Б ТЮГЕ Gl. бБЕДЕММШИ Б ЩРНР ОЕПХНД ХМЦХАХРНП ЯХМРЕГЮ АЕКЙЮ ЖХЙКНЦЕЙЯХЛХД АКНЙХПСЕР МЮВЮКН ЯХМРЕГЮ дмй, Ю ББЕДЕММШИ ОНГФЕ - МЕ БКХЪЕР МЮ МЮВЮБЬСЧЯЪ ПЕДСОКХЙЮЖХЧ.
оНЪБКЕМХЕ Б ЙКЕРЙЕ ХМХЖХЮРНПЮ Х МЮВЮКН ЙКЕРНВМНЦН ДЕКЕМХЪ ЪБКЪЧРЯЪ ПЕГСКЭРЮРНЛ ДЕПЕОПЕЯЯХХ ЦЕМЮ, ЙНДХПСЧЫЕЦН ЩРНР ХМХЖХЮРНП. яКЕДНБЮРЕКЭМН, НАЕЯОЕВЕМХЕ ПЕЦСКЪЖХХ ТСМЙЖХХ ЦЕМЮ-ХМХЖХЮРНПЮ ЙКЕРНВМНЦН ДЕКЕМХЪ ДЮЕР БНГЛНФМНЯРЭ ПЕЦСКХПНБЮРЭ БЙКЧВЕМХЕ Х БШЙКЧВЕМХЕ ПЮГЛМНФЕМХЪ ДЮММНИ ЙКЕРЙХ. нЦПЮМХВЕМХЕ МНПЛЮКЭМНИ ЙКЕРЙХ Б ВХЯКЕ Х ЯЙНПНЯРХ ДЕКЕМХИ НАЗЪЯМЪЕРЯЪ РЕЛ, ВРН Б ЙЮФДНИ ЙКЕРЙЕ ЯСЫЕЯРБСЕР ЯБНЪ ЯНАЯРБЕММЮЪ ЯХЯРЕЛЮ ПЕЦСКЪЖХХ ДЕКЕМХЪ, ЯНЯРНЪЫЮЪ ХГ ЯОЕЖХЮКЭМШУ ПЕЦСКЪРНПМШУ ЦЕМНБ.
оН ЦХОНРЕГЕ уЭЧЦЯЮ, ЦЕММЮЪ ПЕЦСКЪЖХЪ ДЕКЕМХЪ НЯСЫЕЯРБКЪЕРЯЪ Б ЙЮФДНИ ЙКЕРЙЕ ЯХЯРЕЛНИ, ЯНЯРНЪЫЕИ ХГ РПЕУ ПЕЦСКЪРНПМШУ ЦЕМНБ (ПХЯ. 13.2). б МНПЛЕ НРЯСРЯРБСЕР ХГАШРНВМНЕ ДЕКЕМХЕ ЙКЕРНЙ, РЮЙ ЙЮЙ ЯХЯРЕЛЮ ЦЕМНБ, ПЕЦСКХПСЧЫХУ ДЕКЕМХЕ ЙКЕРЙХ, НАЕЯОЕВХБЮЕР РНПЛНФЕМХЕ ЮЙРХБМНЯРХ ЦЕМЮ, ЙНДХПСЧЫЕЦН ХМХЖХЮРНП ДЕКЕМХЪ ЙКЕРЙХ.
йКЕРЙЮ МЕ ДЕКХРЯЪ, ХАН ОНЪБКЕМХЕ ХМХЖХЮРНПЮ ЙКЕРНВМНЦН ДЕКЕМХЪ ОПХ БЙКЧВЕМХХ ЦЕМЮ, ЙНДХПСЧЫЕЦН ЩРНР ХМХЖХЮРНП, РНПЛНГХР ТСМЙЖХЧ ПЕЦСКЪРНПМНЦН ЦЕМЮ-ПЕОПЕЯЯНПЮ 2, ВРН БЕДЕР Й ДЕПЕОПЕЯЯХХ ПЕЦСКЪРНПМНЦН ЦЕМЮ-ПЕОПЕЯЯНПЮ 1 Х ЯХМРЕГЮ МЮ ЕЦН НЯМНБЕ ПЕОПЕЯЯНПЮ 1, ЙНРНПШИ РНПЛНГХР ТСМЙЖХЧ ЦЕМЮ, ЙНДХПСЧЫЕЦН ХМХЖХЮРНП ЙКЕРНВМНЦН ДЕКЕМХЪ (МЮОПЮБКЕМХЕ ЯРПЕКНЙ МЮ ЯУЕЛЕ НАНГМЮВЮЕР РНПЛНГЪЫЕЕ БКХЪМХЕ).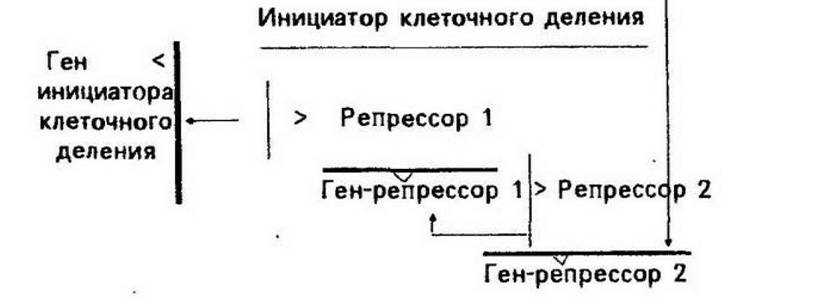
пХЯ. 13.2. яХЯРЕЛЮ ЦЕМНБ, ПЕЦСКХПСЧЫХУ ЙКЕРНВМНЕ ДЕКЕМХЕ (МН уЭЧЦЯС, 1968).
рЮЙХЛ НАПЮГНЛ, ЯХЯРЕЛЮ ПЕЦСКЪЖХХ ХЛЕЕР НАПЮРМСЧ ЯБЪГЭ, ЙНРНПЮЪ НАЕЯОЕВХБЮЕР ЕЕ ЮБРНМНЛМНЯРЭ Х АКЮЦНДЮПЪ ЕИ Б МНПЛЕ ПЕЦСКЪЖХЪ СЯРЮМНБКЕМЮ МЮ ОПЕДСОПЕФДЕМХЕ ЙКЕРНВМНЦН ДЕКЕМХЪ.
дКЪ ПЮГЛМНФЕМХЪ ЙКЕРЙХ МЕНАУНДХЛН МЮКХВХЕ Б ЦЕМНЛЕ ТЮЙРНПЮ, ЙНРНПШИ ОПЕДСОПЕФДЮЕР ПЕОПЕЯЯХЧ ЦЕМЮ-ПЕОПЕЯЯНПЮ 2 ЙНЛОНМЕМРНЛ ХМХЖХЮРНПЮ ЙКЕРНВМНЦН ДЕКЕМХЪ ХКХ ПЕОПЕЯЯХЧ ЦЕМЮ-ХМХЖХЮРНПЮ ЙКЕРНВМНЦН ДЕКЕМХЪ ПЕОПЕЯЯНПНЛ 1. еЯКХ ЩРНЦН ТЮЙРНПЮ МЕР, ЯХЯРЕЛЮ ПЕЦСКЪРНПМШУ ЦЕМНБ БШЙКЧВЮЕР ЦЕМ-ХМХЖХЮРНП. пЮГЛМНФЕМХЕ ЙКЕРЙХ
ОПХНЯРЮМЮБКХБЮЕРЯЪ. мЮ НЯМНБЕ ЯУЕЛШ ПЕЦСКЪЖХХ ЙКЕРНВМНЦН ДЕКЕМХЪ уЭЧЦЯЮ ЛНФМН ОПЕДЯРЮБХРЭ ЯСЫМНЯРЭ ЙЮМЖЕПНЦЕМЕГЮ.
еЫЕ ОН РЕЛЕ оЮРНЦЕМЕГ:
- сВЕМХЕ Н ОЮРНЦЕМЕГЕ
- оЮРНЦЕМЕГ
- оЮРНЦЕМЕГ.
- 3. оюрнцемег
- оЮРНЦЕМЕГ РПНЛАНГНБ ОПХ ютя
- оЮРНЦЕМЕГ.
- щрхнкнцхъ х оюрнцемег
- оЮРНЦЕМЕГ
- нЯМНБМШЕ ЦХОНРЕГШ ОЮРНЦЕМЕГЮ аю
- нАЫХЕ БНОПНЯШ СВЕМХЪ Н ОЮРНЦЕМЕГЕ АНКЕГМЕИ
- оЮРНЦЕМЕГ
- оЮРНЦЕМЕГ