А. Механизм действия метотрексата.
Метотрексат ингибирует ДГФР, выделенную из клеток млекопитающих и бактерий,
несмотря на то, что обычно он не проникает в бактериальные клетки [Nichol, Welch, 1950; Werkheiser, 1953].
Действие метотрексата высокоспецифично: он вызывает 50% ингибирование ДГФР в концентрации 10_9 М, при которой не влияет на любой другой фермент. Этот ингибитор связывается с ДГФР примерно в 104 раз прочнее, чем субстрат (это очень необычное явление) (разд. 9.1), н, следовательно, его действие наименее обратимо по сравнению со всеми обратимыми ингибиторами. Отличия в ингибирующем действии метотрексата на ДГФР, выделенную из разных организмов, очень невелики (в отличие от диамино- лиримидинов, см. ниже) [Werkheiser, 1963]. Отсутствие у метотрексата способности проникать в одноклеточные организмы обусловлено тем, что у них нет системы переноса, имеющейся у млекопитающих. Поскольку метотрексат имеет низкий log Р (—1,85), он не может свободно проникать через цитоплазматические мембраны.Дигидрофолатредуктазы разных типов могут катализировать восстановление как фолата, так и дигидрофолата, хотя дигидрофолат реагирует с ферментами позвоночных в 4-—10 раз быстрее, чем фолат. Скорость восстановления фолата бактериальными ферментами очень незначительна [Blakley, 1969].
Рентгеноструктурный анализ дигидрофолатредуктазы (ДГФР), закристаллизованной с метотрексатом, позволил глубже понять механизм действия этого ингибитора и наглядно представить взаимодействие лекарственного вещества с рецептором [Matthews et al., 1977], позднее Bolin и сотр. (1982) использовали прибор с разрешением до 0,17 нм. Диаграмма фермента ДГФР, его кофермента (NADPH) и метотрексата представлена на рис. 9.4. Изображенный на нем фермент получен из бактерий Lactobacillus casei и те же авторы сообщили о ДГФР (закристаллизованной с метотрексатом), извлеченной из бактерий Е.
coli. Однако им не удалось закристаллизовать метотрексат вместе с ДГФР, извлеченной из организма какого- либо позвоночного.Метотрексат связывается с ферментом, полученным из L. casei, следующим образом. Анион Asp-26 фермента связан с катионом амидиния, образованным из атомов N-1 и 2-NH2 лекарственного вещества (значения рКа приведены в колонке N-1 табл. 9.3). Аминогруппы, связанные с атомом С-4 лекарственного вещества, образуют водородную связь с карбонильными группами Leu-4 и А1а-97. а-Дарбоксильная группа глутаминового фермента метотрексата образует ионную связь с основной боковой цепью Arg-57. Птеридиновое кольцо проникает в полость, ограниченную Leu-4, Ala-6, Leu-27, Phe-ЗО и А1а-97.
пара-Аминобензоильный фрагмент располагается в соседней полости, окруженной липофильными боковыми цепями Leu-27 и Phe-ЗО, с одной стороны, и Phe-49, Pro-50 и Leu-54 — с другой.
Молекула кофермента НАДФН полностью вытянута; никоти- намидная часть располагается напротив птеридинового кольца 38
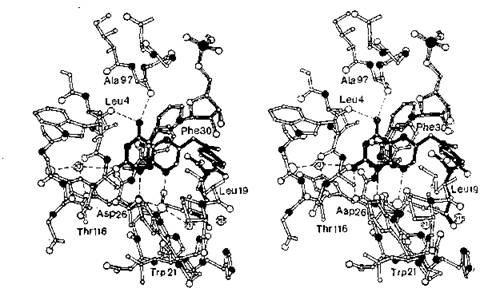
Рис. 9.4. Стереодиаграмма птеридинсвязывающего участка днгвдрофолатрс- дуктазы L. casei. Ингибитор метотрексат выделен жирными линиями, белок — светлыми, молекула НАДФ (показана только ее часть) — пунктиром; атомы углерода изображены небольшими, а атомы кислорода — большими светлыми кружками, атомы азота — черными кружками; цифры в кружках — связанные молекулы воды [Bolin et al., 1982].
в положении, облегчающем перенос гидрид-иона от пиридинового цикла к атому углерода в положении 6 птеридинового кольца [Filman et al., 1982].
Диаграмма для фермента, полученного из бактерий Е. coli, практически идентична представленной на рис. 9.4 [Bolin et al., 1982].
Отличительной особенностью метотрексата является то, что при связывании с ферментом птеридиновые ферменты антиметаболита и природного субстрата ориентированы противоположно. В отличие от метотрексата (см.
выше) для дигидрофолата характерно следующее [Bolin et al., 1982]: атом N-1 не связан, 2-NH2, так же как 0-4, связана только с молекулами воды, атом N-3 образует водородную связь с Asp-26, атом N-5 не связан, а атом N-8 связан с Leu-4 силами ван-дер-ваальса. Таким образом, птеридиновый цикл нормального субстрата в отличие от метотрексата связан с ферментом значительно слабее. Связи, образуемые пара-аминобензоильным и глутаминовым фрагментом метаболита и его антагониста, идентичны. Именно в этом заключается причина необычно высокого индекса ингибирования (1:10 000) метотрексата по сравнению с дигидрофолиевой кислотой.Полагают, что главная причина различной ориентации метотрексата и субстрата при связывании с ферментом заключается в разных положениях и основности наиболее основного атома азота в молекуле (см. табл. 9.3). Метотрексат почти в сто раз более сильное основание, чем дигидрофолиевая кислота, благо-
Таблица 9.3. Величины рКа фолиевой кислоты и ее производных (вводе), полученные: А — с помощью спектрофотометрии, В — с помощью ЯМР-образ- цов, обогащенных 13С по атому С-2 [Рое, 1977; Cocco et al., 1981]
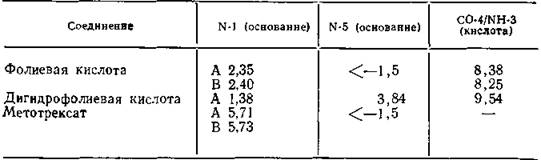
даря замене циклической амидной группы (CO-4/NH-3) аминогруппой. Следует отметить, что наиболее основным центром метотрексата является амидиновая группа N’ = C2—NH2, находящаяся в сопряжении, в то время как наиболее основный центр в молекуле дигидрофолиевой кислоты расположен в другом цикле птеридинового ядра и не соприкасается с каким-либо противоположно заряженным центром молекулы фермента.
Наиболее основный центр молекулы метотрексата имеет рКа 5,71 и, следовательно, только 2% его будут существовать в виде катиона при pH 7,3 в воде (табл. 17.1). Поэтому если даже предположить, что молекула метотрексата образует лишь одну ион-дипольную связь с молекулой аспарагиновой кислоты, то и этого было бы достаточно для того, чтобы обеспечить большое конкурентное преимущество метотрексата по сравнению с дигидрофолиевой кислотой.
Тем не менее следует ожидать, что величина рКа, определенная в воде, в действительности будет значительно выше внутри выстланной липидами полости, которой точно соответствует птеридиновое кольцо молекулы метотрексата, но не таковое дигидрофолата. Это повышение основности обусловлено понижением диэлектрической постоянной и отсутствием конкурирующих молекул воды. Предположение о повышении основности молекулы метотрексата при связывании с ферментом подтверждают также данные ЯМР-спектров, снятых для метотрексата, обогащенного изотопом 13С (до 90%) по атому С-2 [Cocco et al., 1981]. Установлено, что характеристичный сигнал атома С-2 в катионах сохраняется вплоть до pH 10, в то время как в аналогичных условиях основный рКа фолиевой кислоты, равный 2,4, не изменяется. Неизвестно, что именно происходит с дигидрофолиевой кислотой в этих условиях, так как ее недостаточно высокая стабильность не позволяет сделать обоснованных выводов. (Все исследования проводились на ДГФР, полученной из Streptococcus faecium.)
Еще по теме А. Механизм действия метотрексата.:
- Механизм токсического действия
- Механизм действия.
- Механизм токсического действия
- Б. Упрощение молекулы метотрексата.
- Механизм токсического действия
- Механизм токсического действия
- Механизм действия НПВП
- Механизмы радиозащитного действия.
- Механизм токсического действия.
- Механизмы лечебного действия групповой психотерапии.
- Механизм токсического действия
- Механизм действия ТЦА
- Механизм токсического действия
- Механизм токсического действия
- Механизм токсического действия
- Механизм токсического действия