Биохимические различия, способствующие избирательности
Любую биологическую систему можно рассматривать как арену постоянной борьбы за следовые количества/катионов тяжелых металлов. При поражении позвоночных/микроорганиз- мами в организме-хозяине быстро понижается /уровень железа и цинка в плазме.
Из всех биологически значимых тяжелых металлов наиболее сильным действием на бактерии обладает железо, а цинк воздействует на дрожжи и другие грибы. Уровень железа в плазме млекопитающих понижается за счет переноса его в печень; в то же время уменьшается его всасывание в кишечнике [Weinberg, 1974]. При тяжелых хронических инфекциях уровень меди в плазме повышается на 50—90% [Weinberg, 1972]; это повышенное содержание меди, как полагают, является антипаразитическим.Аналогичные явления отмечают у онкологических больных, у которых понижается уровень содержания железа в плазме [Konaka, Matsuoka, 1967] и повышается концентрация меди [Mortazani et al., 1972]. Иногда из-за наличия злокачественной опухоли понижается концентрация цинка в плазме [Davies, Musa, Dormandy, 1968], что, по-видимому, является реакцией организма, так как у многих животных наблюдали ингибирование роста опухоли при исключении из рациона цинка [De Wys, Pories, 1972].
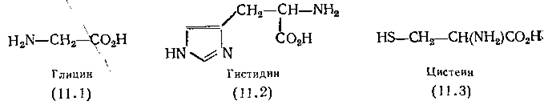
С помощью каких же веществ происходит связывание катионов тяжелых металлов в живых клетках? Следует отметить, что таких веществ великое множество, но в первую очередь внимания заслуживают аминокислоты. Большинство аминокислот, входящих в состав белков, имеет примерно такие же константы связывания, как глицин (11.1). Необычно высокой способностью связывать металлы обладают две аминокислоты: гистидин (11.2) благодаря наличию в его молекуле имидазольного кольца и цистеин (11.3) за счет тиольной группы [Albert, 1952]. К этим двум примерам можно добавить цистин — эффективный, специфически действующий агент, способный связывать медь [Hawkins, Perrin, 1963]. В настоящее время очевидно, что белки связывают металлы в основном с помощью гис- тидиновых и цистеиновых остатков.
Так рентгеноструктурный анализ миоглобина показывает, что между атомом железа и атомом азота имидазольного кольца существует прочная связь.Компьютерный анализ результатов потенциометрического изучения 160 равновесных взаимодействий между Cu2+, Zn2+ и 17 аминокислотами (все присутствовали одновременно) показал, что 85% меди образует комплекс гистидин—медь—цис-
теинв соотношении 1:1:1 и 67% цинка связано с цистеином и гистиіціном [Hallman, Perrin, Watt, 1971].
В кровотоке аминокислоты связаны с очень незначительным количеством меди и цинка. Около 90% всего количества меди находится в связанном состоянии, образуя с белком церулоплазмином комплекс ярко-синего цвета, причем связь эта настолько прочна, что никакого обмена не наблюдается. Концентрации обменивающихся ионов меди и цинка в плазме крови составляют 1,1 мкМ и 15 мкМ соответственно [Agarwal, Perrin, 1976]. Почти все обменивающиеся ионы меди связаны с концевыми остатками альбумина NH2-Asp-Thr-His (в плазме быка) и NH2-Asp-Ala-His (в плазме человека). В связывании участвуют ЫН2-группы, два пептидных атома азота и имидазольный атом азота гистидинового остатка. Условная константа связывания (выражаемая как log К1) составляет 12,1 при pH 7,4 для комплекса медь — альбумин (1:1) и 7,6 для комплекса цинк — альбумин (1 : 1), хотя характер связывания цинка и меди различается. Цинк в отличие от меди легко удаляется из; плазмы крови при инъекции комплекса кальций—ЭДТА. (11.27).
Металл в качестве кофактора необходим примерно 12% всех известных ферментов. Какую же роль он играет в этих, случаях? По-видимому, металл часто служит мостиком между субстратом и белком, причем последний способен активировать металл, акцептируя его свободные электроны. Образование избыточного положительного заряда на атоме металла приводит к тому, что электронная плотность смещается от субстрата к металлу.
При этом уменьшение свободной энергии активации субстрата происходит за счет изменения его электронной- структуры под действием металла.Подобная картина наблюдалась и при исследовании двух содержащих марганец ферментов — креатинкиназы и мышечной енолазы методом ЭПР. Было показано, что у первого из них марганец образует связь с коферментом (АДФ) и с субстратом, но не с белком, а у второго — атом марганца, по-видимому, служит мостиком между субстратом и белком [Cohn, Leigh, 1962]. В других случаях роль металла, по-видимому, сводится исключительно к обеспечению необходимой третичной структуры белка (по аналогии с дисульфидным мостиком S—S). При образовании требуемой третичной структуры часто происходит сближение двух или трех остатков аминокислот, образующих активный центр, тогда как в вытянутой полипептидной цепи эти остатки располагаются далеко друг от друга. Примером •может служить атом цинка в молекуле щелочной фосфатазы •бактерий Е. coli: при удалении атома цинка спектр кругового дихроизма этого фермента резко изменяется под действием 6М мочевины, так как природная конформация превращается в беспорядочную спираль [Trotman, Greenwood, 1971]. Подробнее об этом ферменте см. разд. 11.0.
Многие ферменты, например трипсин, функционируют без участия металла. Однако те ферменты, которым необходимы тяжелые металлы, обычно удерживают их очень прочно. Поэтому во многих случаях металл не удается удалить диализом и даже сильные хелатирующие агенты проникают в клетки микроорганизмов, не причиняя им вреда (разд. 11.7.1). Вероятнее всего, это обусловлено спецификой третичной структуры, а не высокими значениями констант устойчивости (см. выше).
Кроме аминокислот, белков и пептидов существует много других металлосвязывающих соединений, играющих важную роль во всех живых клетках. К ним прежде всего относятся птеридины (в том числе фолиевая кислота) и пурины, для комплексов которых с металлами были определены константы устойчивости [Albert, 1953; Albert, Serjeant, I960]; рибофлавин, ■обладающий наибольшим сродством к металлу в частично восстановленном состоянии [Hemmerich, Veeger, Wood, 1965], спермин (11.4) и диамины спермидин (11.5) и путресцин (11.6).
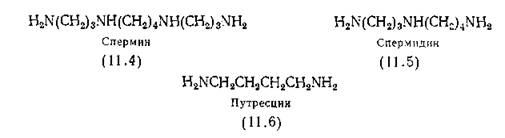
Все виды фосфатов способны связывать металлы в живых клетках. Говоря о неорганических фосфатах, следует отметить, что для орто-фосфатов (Са2+, Mg2+, Zn2+, Cu2+) значения констант устойчивости были заново определены и выяснилось, что комплексы, образованные в соотношении 1 : 1, например СаНРО4, имеют тенденцию к образованию димеров типа Са2Н2(РО4)2 [Childs, 1970]. Данные для аналогичных пирофосфатов можно найти в работе Wolhoff, Overbeck (1959). Определены также константы устойчивости комплексов АТФ с Mg2+, Са2+, Мп2+, Со2+, Ni2+, Zn2+ и Cu2+ [Perrin, Sharma,
1966] и аналогичные величины для АМФ, АДФ, УТФ, ГТФ, ЦТФ, ИТФ [Phillips, 1966]. Как показывают потенциометрическое титрование и метод ЯМР 31Р, ионы магния прочно связаны с фосфатными группами АТФ [Tuck, Baker, 1973]. Константы устойчивости комплексов Си2+, Са2+ и Fe3+ с ДНК имеют тот же порядок, что и комплексы этих металлов с АМФ, имею- 146
щим 'значительно меньшее сродство к катионам металлов, чем;- АТФ pBryan, Frieden, 1967]. Некоторые катионы более тяжелых металлов в природе связаны с молекулами нуклеиновых кислот. Предполагают, что ионы цинка образуют координационную связь с пуриновыми основаниями при обратимом скручивании и раскручивании спирали ДНК [Shin, Eichhorn, 1968]. Жирные кислоты цикла лимонной кислоты также связывают катионы металлов.
Насколько известно, константы устойчивости большинства перечисленных веществ близки по величине к константам устойчивости комплексов аминокислот (табл. 11.1). Только у порфи- ринов константы имеют более высокие значения; порфирины настолько прочно удерживают железо, что не удалось обнаружить никаких признаков обмена с его радиоактивным изотопом [Hahn et al., 1940].
Из вышесказанного ясно, что опыты с микроэлементами нельзя проводить в присутствии фосфатного или цитратного буфера. Для pH в пределах 7,0—8,2 удобным буфером, не образующим хелатных связей, служит N-этилморфин (температура кипения 138—139 °С). Подробнее о других буферах см. Perrin, Dempsey (1974).
Хелатообразующие соединения, избирательно действующие на организмы разных классов. Исследования в области сравнительной биохимии постоянно выявляют новые видовые различия в связывании и использовании металлов (разд. 4.2 и 11.0).
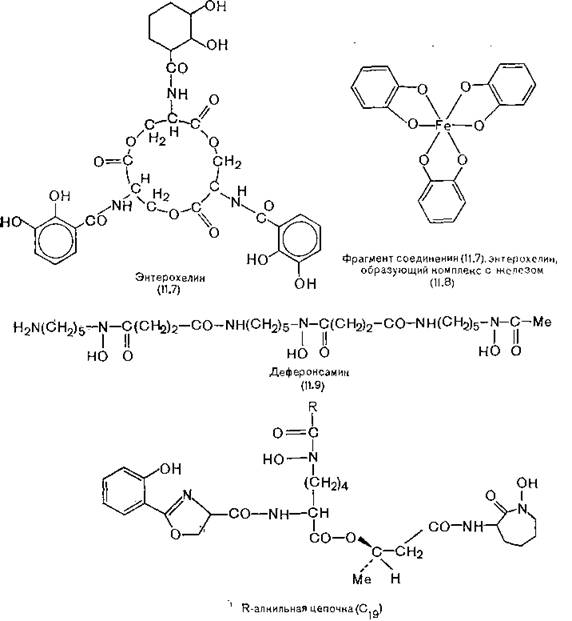
Аэробным бактериальным клеткам необходимо пополнять запас железа из окружающей их среды, где оно содержится, в основном в виде почти нерастворимого оксида железа (III) (количество растворяющегося вещества составляет 10~39 М). Однако существуют соединения, способные растворять и переносить железо, а также превращать его в биологически доступную форму. Эти так называемые сидерохромы содержатся в бактериях, дрожжах и грибах. Из них наиболее распространены феноляты, например энтерохелин (энтеробактин) (11.7) и гидрок- саматы, например дефероксамин В (11.9). Сидерохромы обоих типов представляют собой слабые кислоты (рКа около 9) и способны очень прочно связывать железо (III) (log К 45).
Наиболее важный сидерохром типа енолятов — энтерохелин, циклический триэфир N-(2,3-дигидроксибензоил)-L-серина. Это- соединение выделено из бактерий Е. coli [O’Brien, Gibson, 1970]. Gibson и Nielands (1974) установили механизм действия энтерохелина, использовав серию мутантов, каждый из которых был лишен способности осуществлять одну из биохимических стадий. Рентгеноструктурный анализ показал, что в молекуле энтерохелина каждый катион железа (III) связан с шестью атомами кислорода, предоставляемыми тремя фрагментами кате- холдианионов (11.8), что и придает комплексу пурпурную окраску [Anderson et al., 1976]. Бактерии выделяют энтерохе- 10*
лин, который образует комплекс с железом из окружакэщей среды; затем этот комплекс поглощается бактериями И/Гидро- лизуется специальными ферментами с образованием железа {O’Brien, Сох, Gibson, 1971J.
Микобактин Т (11.10)
Дефероксамины, подобные соединению (11.9), представляют собой гексапептиды, выделяемые из Streptomyces. Рентгеноструктурный анализ показал, что шесть атомов кислорода, принадлежащие трем группам гидроксамовой кислоты, образуют тетраэдрическую структуру, связывающую ион железа (III). Для этого соединения log К, равный 31, значительно выше, чем для ЭДТА (11.27) (log К 24), но ниже, чем log К энтерохелина [Schwarzenbach, Schwarzenbach, 1963].
Сидерохром микобактин Т (11.10) встречается у возбудителей туберкулеза человека. Восемь других микобактинов были получены из других разновидностей рода Mycobacteria. Эти соединения имеют только две гидроксаматных группы, однако наличие фенольной группы, участвующей в трехцентровом связывании иона железа, делает комплекс микобактинов с железом таким же прочным, как и комплексы с другими сидерохро- мами [Snow, 1970]. Это единственные природные соединения железа, имеющие хорошую липофильность.
Значительные различия в использовании железа бактериями и млекопитающими открывают большие возможности для химиотерапии. О применении сидерохромов в качестве антидотов при отравлениях железом см. разд. 11.6.
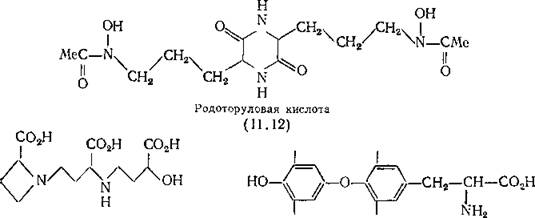
Многие грибы, например Aspergillus aerogenes, выделяют энтерохелин, тогда как другие (в том числе Aspergillus niger и Penicillium reticulosum) удерживают железо с помощью ферри- хрома, циклического гексапептида, в молекуле которого ион железа прочно связан тремя гидроксаматными группами. Грибы содержат множество хелатирующих пиронов, таких, как койевая кислота (З-окси-ч-пирон) (11.11), суммарное значение логарифма константы устойчивости (^3) которой для железа (III) равно 25. Биологическая роль этих соединений неизвестна, так же как и роль полигидроксиантрахинонов, содержащихся в грибах. Все дрожжи выделяют производное гидроксамовой кислоты — родоторуловую кислоту (11.12) для доставки железа в клетки.
Микробный метаболизм железа см. обзор Nielands (1974). О
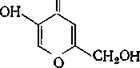
Койевая кислота
(11.11)
Мугинеивая кислота (11.13)
Тироксин
(П.14)
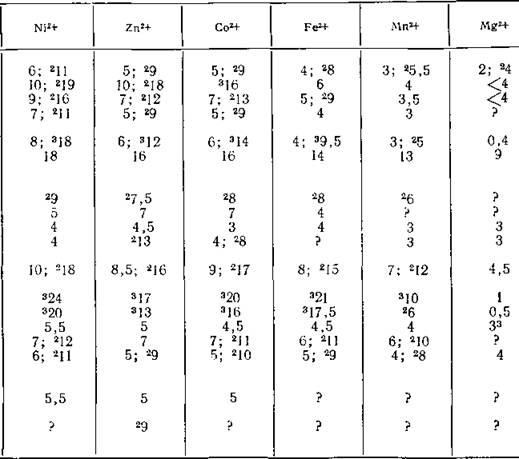
Таблица 11.1. Логарифмы констант устойчивости некоторых обозначены
| Лигаид | Р*а | Fe3* | Си2* | |
| Глицин (11.1) | 9,9; 2,4 | 10 | 8,5; 215 | |
| Цистеин (11.3) | 10,3; 8,4; 2,0 | см.1 | см.1 | |
| Гистидин (11.2) | 9,2; 6,0; 1,8 | ? | 10,5; 219 | |
| Гистамин (7.6) | 9,8; 6,0 | 4 | 10; 216 | |
| Этилендиамин | ||||
| (рис. 11.2) | 10,1; 7,0 | р | 11; 2202 | |
| ЭДТА (11.27) | 11,0; 6,3; 2,7; 2,0 | 24 | 19 | |
| Птероилглутаминовая кислота (фолиевая | ||||
| кислота (см. 9.20) | 8,3 | ? | 28 | |
| Гипоксантин (9.52) | 8,9; 2,0 | ? | 6 | |
| Гуанознн | 9,3 | ? | 6 | |
| Аденин (4.3) | 9,8; 4,3 | ? | 214 | |
| 8-Гидроксихннолин (ок- | ||||
| син) (11.30) | 11,2; 3,8 | 12; 224; 336 | 12; 223 | |
| орто-Фенантролин | ||||
| (11.18) | 4,9 | 314 | 220 | |
| Бипиридил (11.19) | 4,3 | р | 317 | |
| Щавелевая кислота | 4,2; 1,2 | 10 | 6 | |
| Салициловая кислота | 13,4; 3,0 | 16; 228 | 11; 219 | |
| Тетрациклин (11.36) | 9,6; 7,8; 3,4 | 10; 325 | 8; 213 | |
| Гидразид изоникотнно- вон кислоты (изониа- | ||||
| зид) (11.37) | 10,8; 3,5; 1,9 | ? | 8 | |
| Диметилдитиокарбами- | ||||
| новая кислота (3.57) | 3,2 | ? | 11; 222 |
1 Цнстеин окисляется этим катионом.
2 Ср. Ч1 для Си+.
4 Для Са2+ также ~3.
Комплекс марганца с бнпиридилом 2 : 1 (log р 6) обозначен как 2б, а ЭДТА 1 : 1
Данные для тетрациклина см. [Albert, Rees, 1956], изоииазида [Albert, 1956], сали днметнлдитнокарбаминовой кислоты [Janssen, 1958], аминокислот [Albert, 1950, 1952],
Растения также содержат специальные химические соединения, способные связывать железо. Многие злаковые культуры выделяют в почву мугинеивую кислоту (11.13), способную образовывать с ионом железа (Fe3+) комплекс, который затем поглощается и используется растениями. Мугинеивая кислота представляет собой производное азотсодержащего циклического соединения азетидина. В отсутствие необходимого количества железа у растений наступает хлоротическое состояние и они увядают [Mino et al., 1981].
Некоторые природные агонисты млекопитающих, а именно тироксин (11.14), норадреналин (7.5) и гистамин (7.6) способны связывать металлы. Только у млекопитающих в митохондриях накапливается кальций.
комплексообразующих агентов (в воде при 20 °С; насыщенные константы как Jog р)
(log р 13) — как 13.
цнловон кислоты [Perrin, 1958], бнпириднла и орто-фенантролнна [Irving, Mellor, 1962], остальные значения [Sillen, Martell, 1964, и дополнения].
11.2.
Еще по теме Биохимические различия, способствующие избирательности:
- Химические различия, способствующие избирательности
- Различия в ионизации, обеспечивающие избирательность
- Классификация ядов по избирательной токсичност
- Осаждение путем избирательной денатурации
- Избирательное смотрение и слушание
- Способствующие факторы.
- Избирательное вербальное внимание, семантические системы мозга и их связь с памятью
- Примечание: Р - уровень достоверности различий показателей по сравнению со здоровыми мальчиками; Рі - уровень достоверности различий показателей по сравнению с больными II и III группы.
- роль психологии в диагностике личности. приемы, способствующие получению информации
- Формирование избирательной функциональной организации нервных центров при зрительно-вербальной деятельности