Особенности мембран хрусталиковых волокон.
Контакты типа ’’пугови- ца—петля”. Мембранысоседствующих хру-
СТаЛИКОВЫХ волокон соединены при помощи разнообразных специализированных образований, изменяющих свое строение по мере смещения волокна с поверхности вглубь хрусталика.
В поверхностных 8-10 передних слоях коры волокна соединяются при помощи образований типа “пуговица—петля” ("шар и гнездо"), распределенных равномерно по длине волокна (рис. 2.16). Подобного типа контакты существуют только между клетками одного слоя, то есть клетками одного поколения, и отсутствуют между клетками разных поколений. Это обеспечивает возможность передвижения волокон относительно друг друга в процессе их роста.В более глубоко расположенных волокнах контакт типа“пуГ0ВИЦа—петля“ обнаруживается несколько реже и распределяется вдоль волокна неравномерно и непроизвольно. Видны они и между клетками разных поколений.
В самых глубоких слоях коры и ядра кроме указанных контактов (“пуговица—
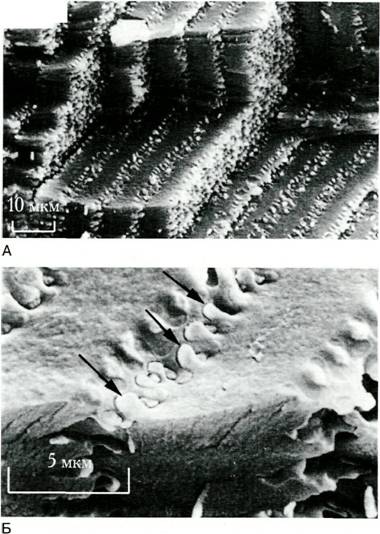
Рис. Характер расположения хрусталиковых
волокон. Сканирующая электронная микроскопия. Видны плотно упакованные хрусталиковые волокна (A) и контакты типа «пуговица—петля» (Б) петля") появляются сложные интердигита- ции в виде гребней, впадин и борозд. Обнаружены также иДесМОСОМЫ, но только между дифференцирующимися, а не зрелыми хрусталиковыми волокнами.
Предполагают, что контакты между хрусталиковыми волокнами необходимы для поддержания жесткости структуры на протяжении всей жизни, что способствует сохранению прозрачности хрусталика.
Еще один тип межклеточных контактов обнаружен в хрусталике человека. Это щелевой контакт. Предполагают, что такие контакты выполняют две роли. Во-первых, поскольку они соединяют хрусталиковые волокна на большом протяжении, сохраняется архитектоника ткани, тем самым обеспечивается прозрачность хрусталика.
Во- вторых, именно благодаря наличию этих контактов происходит распространение питательных веществ между хрусталиковыми волокнами. Это особенно важно для нормального функционирования структур на фоне пониженной метаболической активности клеток (недостаточное количество органоидов).Выявлено два типа щелевых контактов — кристаллические (с высоким омическим сопротивлением) и некристаллические (с низким). В некоторых тканях (печень) указанные типыЩвлевидных контактов могут преобразовываться один в другой при изменении ионного состава окружающей среды. В волокне хрусталика они не способны к подобному преобразованию. Первый тип щелевых контактов найден в местах прилегания волокон к эпителиальным клеткам, а второй — только между волокнами.
Второй тип щелевых контактов (низко- ОМНЬіе) имеютвнутримембранные частицы, не позволяющие соседним мембранам сближаться более чем на 2 нм. Благодаря этому в глубоких слоях хрусталика уровни ионов и молекул невысокие; последние достаточно легко распространяются между хрусталиковыми волокнами и их концентрация довольно быстро нормализуется.
Имеются и видовые различия в количестве щелевых контактов. Так, в хрусталике они занимают такую площадь от поверхности волокна: у человека — 5%, у лягушки — 15 %, у крысы — 30%, а у цыпленка — 60 %. Щелевых контактов нет в области швов (Kuszak и соавт., 1985).
Высокая рефракционная способность хрусталика достигается высокой концентрацией белковых филаментов, а прозрачность обеспечивается их строгой организацией, однородностью структуры волокон в пределах каждого поколения и небольшим объемом межклеточного пространства (ме- нее1% объема хрусталика). Способствует прозрачности и небольшое количество внутрицитоплазматических органоидов, а также отсутствие вХрусталикоВЫХ волокнах ядер. Все перечисленные факторы сводят к минимуму рассеивание света между волокнами.
Есть и другие факторы, влияющие на рефракционную способность. Одним из них является повышение концентрации белка по мере приближения к ядру хрусталика.
Именно благодаря этому отсутствует хроматическая аберрация.Не меньшее значение в структурной целостности и прозрачности хрусталика имеет и регуляция ионного содержания и степени гидратации волокон.
При рождении хрусталик прозрачен. С возрастом по мере его роста ядро приобретает желтоватый оттенок, что, вероятно, связано с влиянием на него ультрафиолетового излучения (длина волны315_400 нм). При этом в коре появляются флюоресцирующие пигменты. Предполагают, что эти пигменты экранируют сетчатку от разрушительного действия коротковолновой световой энергии. Пигменты накапливаются в ядре с возрастом, а у некоторых лиц участвуют в образовании пигментной катаракты. В старческом возрасте и, особенно, при ядерной катаракте в ядре хрусталика увеличивается количество нерастворимых белков, которые представляют собой кристал- ЛИНЫ со «сшитыми молекулами».
Метаболическая активность в центральных участках хрусталика незначительна. Отсутствует метаболизм белков. Именно поэтому они относятся к долгоживущим белкам и легко подвергаются повреждению окислителями, которые приводят к конформации белковой молекулы и образуют сульф- ГИДрильные группы. Развитие катаракты характеризуется увеличением зон рассеивания света. Это может быть вызвано нарушением регулярности расположения хрусталиковых волокон, изменением структуры мембран и рассеиванием, связанным с преобразованием белковых молекул. Отек хрусталиковых волокон и их разрушение приводят к нарушению водно-солевого обмена.
Еще по теме Особенности мембран хрусталиковых волокон.:
- ГЛАВА 2 ТОПОГРАФИЧЕСКАЯ АНАТОМИЯ ГЛАЗА
- Профилактика и мероприятия по устранению осложнений, возникающих в ходе операции
- Наружная оболочка глаза
- ОШИБКИ ПРИ МАНИПУЛЯЦИЯХ НА ХРУСТАЛИКЕ
- Внутриглазная коррекция афа-кии
- Форма и размер хрусталика.
- Процесс формирования хрусталиКОВЫХ волокон.
- Особенности мембран эпителиальных клеток.
- Особенности мембран хрусталиковых волокон.
- Неонатальные хрусталиковые вакуоли.
- Ленарственная натаранта