Топографическое деление коры.
Ширина всей коры, количество клеток и ширина каждого слоя клеток на фронтальном разрезе, а также клеточный состав, т. е. величина, форма, расположение клеток в каждом участке коры чрезвычайно варьируют (рис.
5). Эти вариации занимают определенные, ясно отграниченные территории. В большом мозге млекопитающих насчитывают 11 хорошо выявленных больших корковых полей. На основании гистологических и физиологических исследований каждое такое поле было разделено еще на несколько самостоятельных участков. Более значительная дифференцировка коры отмечается у высших млекопитающих — обезьян (рис. 6). Она особенно велика у человека. Поверхность коры у человека может быть разделена на 200 полей (Ц. Фохт и О. Фохт, 3).Приводимая дифференциация в строении коры в общем совпадает с данными физиологии и клинического материала о локализации функции в коре мозга. Особенно показательны исследования неврологов Ц. Фохт и О. Фохт с электрическим раздражением коры обезьяны церкопитека. Они обнаружили полное соответствие полученных этим путем физиологических результатов с гистологически установленной архитектоникой мозга животных. На основании своих исследований эти авторы составили предположительную карту полей для человеческого мозга. Она оказалась очень сходной с той, которая была составлена О. Ферстером (4) для человеческого мозга на основании раздражения коры при операциях.
9

Рис. 3. Аксонные разветвления главных типов пирамидных нейронов с нисходящими аксонами (а)
/ и 5 — вставочные нейроны, их аксоны не выходят из пределов коры; 4, 7, Ю w tf — ассоциационные пирамиды, 8 — проекционный нейрон (Лоренте де Но)
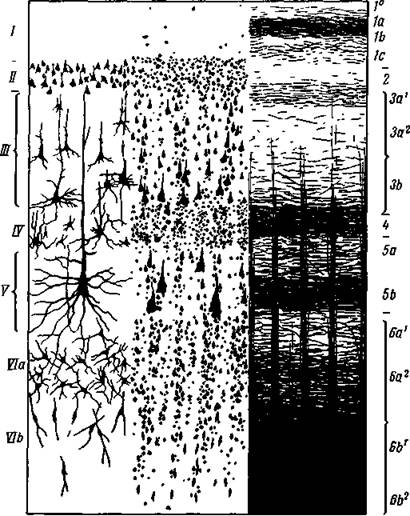
Рис. 4. Слои нервных клеток и пучков нервных волокон в коре большого мозга человека
Схематическое^изображение наиболее типичной для коры шестислойной архитектоники.
С левой стороны даны клеточные слои: I — зональный или молекулярный слой; II — внешний зернистый; III — пирамидный; IV— внутренний зернистый; V — ганглионарный или внутренний пирамидный; VI — полиморфный. С правой стороны дано расположение нервных пучков в тех же слоях (по Бродману и Фохту)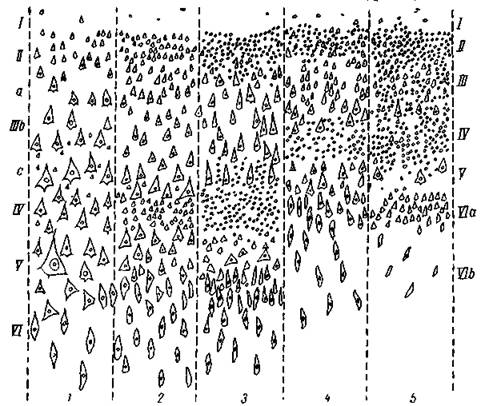
Рис. 5. Типы архитектонического строения коры больших полушарий человека
1 — тип двигательной области; 2 — фронтальный тип; з — париетальный тип; 4 — полярный тип; 5 — гранулярная, зернистая кора (Экономо)
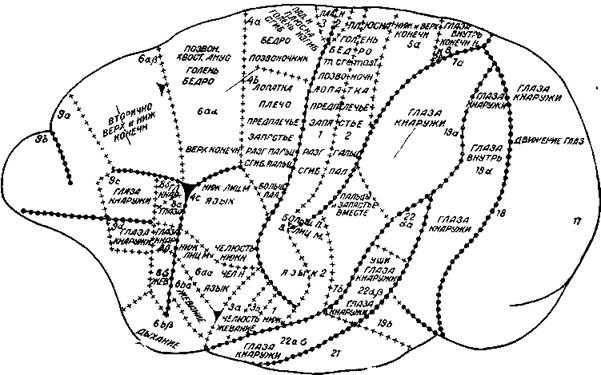
Рис. 6. Деление коры церкопитека (обезьяны) на отдельные поля на основании физиологического и гистологического исследований
Указывается, какие именно части тела реагировали на электрическое раздражение. Цифры обозначают нумерацию отдельных полей по гистологическим исследованиям (Ц. и О. Фохт)
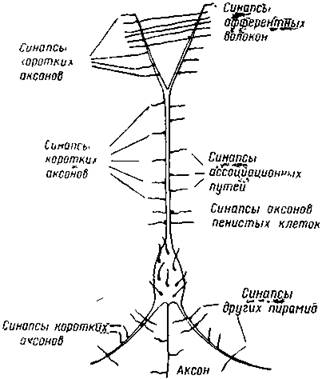 |
| Рис. 7. Пирамидный нейрон с двумя базальными дендритами и с верхушечным дендритом На дендритах и на теле клетки оканчивается множество синапсов от разнообразных нейронов (Лоренте Де Но) |
Микроскопическая структура коры высших позвоночных была изучена Лоренте де Но (5) на разных млекопитающих, включая и обезьяну. Подобно Р. Кахалу (6), он показал, что нейроны всех слоев коры мозга млекопитающих образуют своими сильно ветвящимися дендритами густую войлочную сеть. Многообразный нейронный состав двух наружных слоев коры мозга совсем не дает выходящих из коры аксонов. Эти аксоны клеток коротки и оканчиваются частью в том же слое, где лежат клетки, частью в соседних слоях, выше или ниже этого слоя. Иначе говоря, все нейроны первых двух слоев являются внутрикорковыми. Только в нижних четырех слоях аксоны значительного количества пирамидных клеток выходят из коры, но и здесь немало нейронов с короткими аксонами, осуществляющими связь нейронов этого слоя между собой и с вышележащими слоями.
Очень характерно существование так называемых «возвратных» аксонов и коллатералей. Аксоны некоторых нейронов сначала идут вниз, а затем поворачивают обратно и заканчиваются в вышележащих слоях. Во многих случаях поворачивает обратно не весь аксон, а его коллатерали (рис. 3). Все эти аксоны и их коллатерали оканчиваются конечными или касательными синапсами в том или другом слое коры на поверхности тел клеток и их дендритов. На клетках и дендритах эти синапсы располагаются сотнями, иногда сплошным слоем (рис. 7).Афферентные волокна, приходящие в кору из нижних отделов мозга, так называемые таламические волокна, заканчиваются преимущественно в четвертом слое, лишь немногие из них оканчиваются в третьем и пятом слоях и редко дают коллатерали к остальным слоям (рис. 1, 2). Лоренте де Но (5) находит, что эти афферентные волокна своими оканчаниями приходят в множественный синаптический контакт с находящимися здесь дендритами и телами клеток. Отсюда следует, что пирамидные нейроны нижних слоев коры могут иметь прямую связь с афферентными таламическими волокнами главным образом в своей дендритной части, распространяющейся во втором и третьем слоях. Но ввиду многочисленных аксонных связей вставочных пирамидных нейронов второго, третьего и четвертого слоев с пирамидами нижних слоев эти последние должны быть связаны с афферентными волокнами также и через эти нейроны.
Еще по теме Топографическое деление коры.:
- Развитие гигиены в Беларуси
- АНАТОМИЯ И РЕНТГЕНОАНАТОМИЯ СОСУДОВ головы
- Билет 33. Когнитивно-поведенческое направление в психотерапии.
- Когнитивно-поведенческое направление в психотерапии.
- Топографическое деление коры.
- Эмоциональные реакции при раздражении миндалевидногокомплекса.
- ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
- ЭЛЕКТРОННОЕ ОГЛАВЛЕНИЕ
- АРХИТЕКТОНИКА КОРЫ БОЛЬШИХ ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
- Глава 6 Высшие психические функции и их нарушения
- Медицинская помощь беременным.
- Зрительный путь и путь зрачкового рефлекса
- ГЛАВА 2 ТОПОГРАФИЧЕСКАЯ АНАТОМИЯ ГЛАЗА
- Наружная оболочка глаза
- ОШИБКИ ПРИ МАНИПУЛЯЦИЯХ НА ХРУСТАЛИКЕ
- СОСУДИСТЫЕ БОЛЕЗНИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ.
- Медиаторные синаптические системы дорзального стриатума и их роль в регуляции моторного поведения
- Роль холинергической системы стриатума в регуляции двигательного поведения