Медиаторные синаптические системы дорзального стриатума и их роль в регуляции моторного поведения
Одно из принципиальных различий между физиологической реакцией медиаторной и модуляторной передачи состоит, кроме всего прочего, во времени реакции. Синаптические реакции - очень быстрые, они измеряются миллисекундами, тогда как только латентные периоды модуляторных реакций измеряются сотнями миллисекунд (от 200 до 500 мс, в зависимости от модуляторной системы).
Синаптические системы осуществляют процессы быстрого реагирования. Они представляют группу поведенческих исполнительных реакций или быстро организуемых рефлекторных актов, осуществляемых за очень короткое время. Информация по кортико-стриатным, стриато-таламическим и таламо-кортикальным сетям передается в миллисекундных интервалах (Mink, 2003). Как указывалось выше, одной из ключевых структур, участвующих в реализации поведения, является дорзальный стриатум, или неостриатум. Выше упоминалось, что по сравнению с другими мозговыми областями неостриатум является в высшей степени однородным. Приблизительно 95% его нейронов имеют однотипную морфологию и являются звездчатыми нейронами среднего размера. Это единственные проекционные нейроны в неостриатуме, и их функция связана с интеграцией всех входов в эту мозговую область (Hikosaka et al., 2000; Mink, 2003). Звездчатые нейроны неостриатума получают возбуждающую глутаматергическую синаптическую информацию через AMPA, каинатные и NMDA-рецепторы практически из всех областей неокортек- са (Mink, 2003). Сами звездчатые нейроны ГАМК-ергические, то есть используют ГАМК как передающий информацию медиатор, реакция которого осуществляется через постсинаптический ГАМКА-рецептор. Постсинаптические ГАМКА-рецеп- торы локализованы как на дендритах и соме средних звездчатых стриатных нейронов, так и на нейронах бледного шара, энтопедункулярного ядра и ретикулярной части черной субстанции (Mink, 2003).
Средние звездчатые нейроны хвостатого ядра (nucleuscaudatus) и скорлупы (putamen, или неостриатум) морфологически идентичны, но отличаются нейрохимически.
Одна популяция этих нейронов содержит ГАМК, динорфин, и вещество P как котрансмиттеры и преимущественно экспрессирует Dl-рецепторы. Эти нейроны посылают аксоны во внутренний паллидум (ГПв) и в ретикулярную часть черной субстаниции (ЧСрч) (см. рис. 1) и дают начало прямому пути, который, тормозя интернейроны локальной сети таламуса, активирует нейроны таламокорти- кального входов. Вторая популяция средних звездчатых нейронов содержит ГАМК и энкефалин как котрансмиттер и преимущественно экспрессирует D2-рецепторы. Эти нейроны проецируются к внешнему паллидуму (ГПвне), дают начало непрямому пути и тормозят нейроны таламокортикального входа. Эти две популяции средних звездчатых нейронов, являясь морфологически неразличимыми, также не отдельны топографически в пределах стриатума (Mink, 2003). Такое совместное распределение нейронов предполагает, что они получают общий вход и, таким образом, они могут передать сходную информацию своим соответствующим целям.Однако различие целей и различие типов передатчика этих двух популяций обеспечивают разделение функций в пределах стриатума.
В том случае, когда непрямой ГАМК-ергический путь оканчивается на нейронах ретикулярного ядра таламуса (РЯТ, см. рис. 1), то ингибирование этих нейронов вызывает пачечную активность и осцилляции в таламо-кортикальных нейронах (burstneurons). Это очень интересное явление, так как обычно нейроны активируются с помощью возбуждающих систем. А здесь торможение, или гиперполяризация, вызывает пачечную активность в таламо-кортикалных нейронах. Причиной этой реакции является то, что в нейронах ретикулярного ядра таламуса и соматосенсорной коры локализованы два потенциал-зависимых канала: ^-канал (Clapham, 1998; Bazyan, van Luijtelaar, 2013) и низкопороговый Са2+-канал T-типа (ICa2+T) (Buzsaki et al., 1990; Bazyan, van Luijtelaar, 2013). Активируемый гиперполяризацией катионный 1ь-канал, воротный механизм которого зависит от вторичных посредников, или канал Франчески If (для периферической системы), названный в честь его первооткрывателя (DiFrancesco, 1985), активируется при гиперполяризации, когда мембранный потенциал достигает -80 мВ (Clapham, 1998).
Низкопороговый Ca^-канал T-типа ICa2+T открывается при гиперполяризации, когда мембранный потенциал достигает -90 мВ (Irisawaetal, 1993). Эти два канала взаимосвязаны, так как ингибирование I^+^канала уменьшает пачечную активность If пейсмекерного канала (Hagiwara et al., 1988).Подробное описание функционирования Ih приведены в двух наших обзорах (Базян, Сегал, 2010; Bazyan, van Luijtelaar, 2013). Активируемые гиперполяризацией и модулируемые цАМФ HCN каналы способствуют непосредственной ритмической активации соматосенсорной коре и в ретикулярном ядре таламуса. Известны 4 изоформы субъединиц этого канала (HCN1-HCN4). Каналы HCN, или иначе Ih-каналы, похожи на потенциал-зависимые К+-каналы, они являются тетрамерными и составлены из мономерных субъединиц, состоящих из шести трансмембранных сегментов (S1—S6) с формирующей пору P-петлей между S5- и S6-сегментами или доменами. Различные HCN-субъединицы могут быть совместно объединены друг с другом и формировать гетеромультимерный комплекс, значительно увеличивая разнообразие нативных ^-каналов. Канал HCN1 показывает минимальное изменение в ответ на связывание цАМФ (+4 мВ) со специфическим цАМФ-связы- вающим участком, тогда как HCN2-канал показывает выраженное изменение (+17 мВ). Коэкспрессия HCN1 и HCN2 формирует гетеромультимерный канал и приводит к Ih-токам, которые имеют тенденцию приобретать промежуточные свойства относительно HCN1 и HCN2 гомомерных каналов.
Экспериментальные результаты позволяют предположить, что ^-канал и низкопороговый Са2+-канал Т-типа (ICa2+T) работают в тандеме. Гиперполяризация открывает ^-канал и катионный ток деполяризует мембрану до порогового уровня и вызывает спайк. Гиперполяризация открывает также ICa2+T канал. Вошедший внутрь Са2+ индуцирует Са2+-зависимый синтез цАМФ, который, связываясь с CNBD локусом HCN-субъединиц, резко усиливает активность канала.
Так как HCW-субъединица слабо реагирует на связывание цАМФ, то уменьшение HCN1- субъединиц в составе канала повышает пейсмекерную активность, а увеличение HCNl-субъединиц в составе канала понижает пейсмекерную активность. Анализ приведенных экспериментальных данных показывает, что одним из основных механизмов длительной регуляции активности Ih-пейсмекера является модификация количества HCNl-субъединиц в композиции или в составе пейсмекерного канала. Предполагается, что источником сонных веретен и пик - волновых разрядов в ЭЭГ, является Ih-пейсмекерный канал, локализованный в ретикулярном ядре таламуса и в пирамидных нейронах 3, 4 и 5 слоя соматосенсорной коры. Фармакологические исследования подтверждают это предположение.Ретикулярное ядро таламуса является структурой, которая получает ГАМК-ер- гическое торможение из базальных ганглий - ключевых структур, которые участвуют в реализации поведения. Обзор экспериментальных работ показал, что благодаря локализации Ih и ICa2+ т-каналов в нейронах ретикулярного ядра таламуса, неокортекса гиперполяризация и деполяризации вызывают пачечные разряды в нейронах, которые активируют таламокортикальные сети и способствуют процессам реализации поведения.
Взаимодействие Ih и ICa2+ т-каналов (рис. 3) описано с помощью схемы и количественной модели (Bazyan, van Luijtelaar, 2013).
Тандем Ih и ICaT работает следующим образом.
Гиперполяризация открывает ^-канал мембрана деполяризуется на величину N. Одновременно гиперполяризация открывает ICa2+ т-канал. Са2+ входит в клетку, усиливает аденилат циклазу, повышает концентрацию цАМФ. цАМФ связывается с CNBD-доменом и усиливает воротный механизм на величину M.
Общая величина деполяризации IhDep = N+M.
Когда ^-канал состоит из 4 HCNl-субъединиц, при взаимодействии цАМФ с CNBD HCNl-субъединицы воротный механизм усиливается и деполяризация повышается на 4 mV (Chenetal, 2001).
При этом: 4(HCN1); IhDep = N+(4x4) mV = N+16 mV.
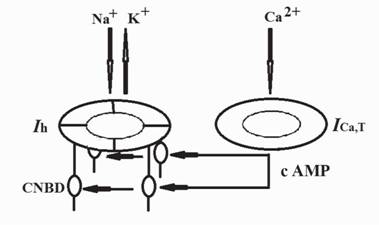
Рис. 3. Схематическое представление взаимодействия Ih и ^+,,-каналов. Ih - активируемый гиперполяризацией, циклический нуклетид активируемый катионный канал, ICaT - низкопороговый Са2+-каналов Т-типа, CNBD (cyclicnu cleotide - binding domain) - область связывающая циклический нуклеотид. Объяснения в тексте (Bazyan, van Luijtelaar, 2013)
При взаимодействии цАМФ с CNBD НС№-субъединицы воротный механизм усиливается и деполяризация повышается на 17 mV (Chen et al., 2001).
Когда уменьшается содержание HCNl-субъединицы в композиции канала, то увеличивается содержание НС№-субъединицы, то есть за счет кругооборота Ih- канал теряет HCNl-субъединицы и замещает их HC^-субъединицами Субъединичные композиции:
3(HCN1) и 1(HCN2); IhDep = N+(3 х 4)mV + (1 х 17)mV = N+29 mV 2(HCN1) и 2(HCN2); IhDep = N+(2 х 4)mV + (2 х 17)mV = N+42 mV l(HCNl) и 3(HCN2); IhDep = N+(1 х 4)mV + (3 х 17)mV = N+55 mV 4(HCN2); Ih Dep = N+ (4 х 17)mV = N+68 mV ГАМК-ергические, поэтому потеря HCN1 субъединиц повышает пейсмекерную активность Ih-канала.
Таким образом, аксоны нейронов неостриатума вызывают пачечную активность в таламических и корковых нейронах и запускают таламо-кортикальные сети, которые участвуют в регуляции моторного поведения. Устойчивый уровень активности нигростриарной дофаминергической (ДА) системы обеспечивает устойчивое торможение стриатных выходов.
3.2.
Еще по теме Медиаторные синаптические системы дорзального стриатума и их роль в регуляции моторного поведения:
- Регуляция моторного поведения
- Медиаторные синаптические системы дорзального стриатума и их роль в регуляции моторного поведения
- Заключение