И к содовые клещи.
У самок Ixodes ricinus и других видов подссм. Ixodinae имеется один тип секреторных альвеол. Каждая альвеола образована 7—9 устьевыми клетками, занимающими базальную половину вокруг альвеолярного выводного протока и расположенных над ними 15—20 заполняющих клеток.
У недавно перелинявших особей продукты секреции не выявляются, цитоплазматическая зона вокруг ядер узкая, а размеры последних близки в обоих типах клеток. Устьевые и заполняющие клетки можно различать на этой стадии лишь по их положению в альвеоле.По окончании послелиночного доразвития устьевые клетки и их ядра становятся несколько крупнее заполняющих. Цитоплазма устьевых клеток сильно вакуолизирована и плохо окрашивается гистологическими красителями (рис. 217). В ней появляются одиночные сферические вакуоли, окрашивающиеся азокармином в красный цвет. Ядра подобных клеток увеличиваются в размерах с 6 до 8 мк и часто приобретают неправильно многоугольную форму. В цитоплазме заполняющих клеток внешние проявления секреторной активности не выявляются (рис. 218).
Прикрепление к хозяину служит стимулом к началу секреции. Размеры альвеол быстро увеличиваются, причем созревание их не синхронно для железы в целом. У питающейся особи одновременно обнаруживаются альвеолы на всех фазах секреторной активности. Размеры заполненных секретом альвеол достигают 160 X140 мк. Многие из них, особенно в начале питания, освобождаются от секрета при значительно меньших размерах (рис. 219, 220).
Цитоплазма зрелых устьевых клеток заполнена вакуолями с секретом. Они достигают в поперечнике 2—4 мк, окрашиваются азокармином в красный цвет и чернятся железным гематоксилином. Перед самым опорожнением содержимое отдельных вакуолей сливается в более крупные капли. Последние, как и секрет в полости альвеол или в выводных протоках, приобретают после азокарминовой окраски различные оттенки синего цвета.
Ядра сморщиваются, и в них прослеживаются пикнотические изменения (рис. 221).Заполняющие клетки также заметно увеличиваются и принимают пирамидальную форму с вершиной в просвет альвеолы. Сферические ядра увеличиваются с 5—7 мк у голодных особей до 9 — 11 мк, а внутри них хорошо выявляются крупные ядрышки. В цитоплазме в большом количестве появляются базофильные волокнистые структуры, по-видимому, являющиеся элементами эндоплазматической сети. В петлях последней, особенно в апикальной части клетки, появляется большое количество мелких вакуолей (не более 1 мк), окрашивающихся азокармином в синий цвет.
Освобождение от секрета зрелой альвеолы происходит очень быстро, так что на срезах редко встречаются промежуточные этапы этого процесса. В первую очередь изливается секрет устьевых клеток, после чего они быстро сморщиваются и дегенерируют. В заполняющих клетках выведение секрета сопряжено с отторжением апикальных участков цитоплазмы. Базальные половины клеток, содержащие ядра, напротив, продолжают увеличиваться в размерах, сильно уплощаются и как бы наползают в направлении выводного протока, занимая место дегенерировавших устьевых клеток. В результате формируется обширная центральная полость и альвеола приобретает вид тонкостенного пузырька диаметром 190 X ХІЗО мк (рис. 222). В дальнейшем альвеолы спадаются, ядра перестают окрашиваться и происходит быстрая дегенерация всех клеточных структур.
У самцов /. ricinus строение слюнных желез сходно с самками, они отличаются лишь значительно меньшими размерами. В них обнаружены пирамидальные и секреторные альвеолы, причем последние образованы из устьевых и заполняющих клеток (рис. 246). Однако в отличие от самок секреторные альвеолы полностью развиты уже у недавно перелинявших голодных особей, а их цитоплазма заполнена продуктами секреции.
У самок Hyalomma asiaticum и других изученных представителей подсем. АтЫуоттіпае наблюдается усложнение строения слюнных желез по сравнению с Ixodinae. В слюнных железах самок мы обнаружили, включая пирамидальные, три типа альвеол, которые, как и в работе В.
Тилл (Till, 1961), в дальнейшем будут обозначаться римскими цифрами, а входящие в их состав клетки — буквами латинского алфавита. В передней части желез непосредственно на стенках главных выводных протоков находятся альвеолы I типа («пирамидальные альвеолы»). Альвеолы II типа встречаются в передних двух третях железы и открываются преимущественно в первичные разветвления главных выводных протоков, а альвеолы III типа, составляющие основную массу железистой ткани, встречаются по всей железе и открываются как в первичные, так и в концевые разветвления выводных протоков.Альвеолы II типа у голодных особей почти правильной сферической формы или слегка суживаются в направлении выводного протока. Диаметр их составляет 28—42 мк. Снаружи они, как и альвеолы других типов, одеты базальной мембраной соединительнотканного происхождения толщиной 400—700 ммк. Она очень хорошо видна на электронномикроскопических фотографиях, тогда как при световой микроскопии даже при максимальных увеличениях выглядит едва различимой одноконтурной линией. Внутри альвеолы находится центральный щелевидный просвет, окруженный цитоплазматической зоной с 17—22 ядрами трех типов (рис. 223).
Проксимальную-половину альвеолы занимают 4—5 крупных клеток, забитых множеством вакуолей секреторной природы («устьевые клетки»
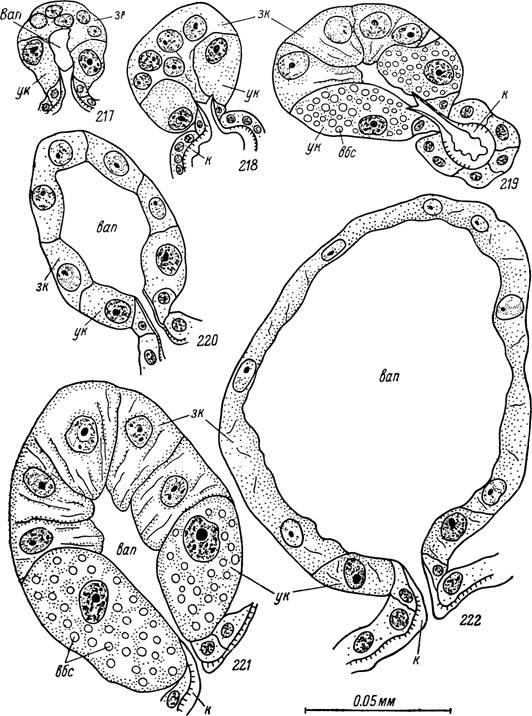
Рис. 217—222. Секреторные альвеолы слюнных желез самки Ixodes ricinus. 217 — начальная и 218 — конечная стадии дифференцировки альвеол у голодной особи; 219 — заполненная секретом альвеола в начале периода роста; 220 — опорожненная альвеола в начале периода роста; 221—222 — то же в конце периода роста.
ух — устьевые клетки; зк — заполняющие клетки; вап — внутриальвеолярная полость; х — кутикулярная выстилка выводных протоков; вбс — вакуоли с белковым секретом.
в прежних исследованиях и тип «а» в настоящей работе). Диаметр секреторных вакуолей колеблется от 1 до 3 мк. Они тесно прилегают друг к другу игюставляют лишь узкие прослойки цитоплазмы.
Азокармином они красятся в темно-красный цвет, а железным гематоксилином — в черный или темно-коричневый. Ядра клеток «а» чаще всего неправильной угловатой или лопастевидной формы, хотя встречаются и почти правильные сферические. Диаметр их составляет 9—14 мк, и они содержат очень большое количество мелких и средних глыбок ДНК, а также крупные сферические нуклеолы.В дистальной половине альвеолы голодного клеща клеточные границы в световом микроскопе обычно неразличимы, но хорошо выявляются при электронной микроскопии. В этой части альвеолы находятся 6—9 более крупных ядер (диаметр 6.5 — 12 мк) сферической формы, принадлежащих клеткам типа «Ь», и 7—9 более мелких (3.5—7 мк) слегка овальных ядер клеток типа «с» (рис. 223).
С началом питания размеры альвеол II типа начинают быстро увеличиваться и они достигают даметра 100—160 мк (рис. 224, 225). Объем внутреннего щелевидного просвета также значительно увеличивается, и в нем, как и в близлежащих частях выводных протоков, на гистологических срезах часто приходится наблюдать коагулировавшую массу секрета. При азокарминовой окраске она приобретает синий оттенок.
Рост альвеол происходит исключительно за счет увеличения размеров клеток «Ь». Диаметр их ядер, сохраняющих сферическую форму, увеличивается в несколько раз и достигает 30—50 мк. Ядра интенсивно окрашиваются различными красителями, а при реакции Фельгена в них выявляется большое количество мелких и средних глыбок ДНК. Особенно характерно для них сильное увеличение и частая фрагментация нуклеол. Клеточные границы внутри альвеол на этой стадии выявляются очень отчетливо. В цитоплазме большое количество волокнистых элементов с высокой базофилией. В ячейках образуемой ими сети появляется множество секреторных вакуолей диаметром до 2—4 мк. Во вполне созревших клетках они заполняют значительную часть цитоплазмы и местами, сдавлирая друг друга, могут приобретать полигональную форму (рис. 225). При аза- новой методике окраски, в отличие от секреторных вакуолей клеток «а», они приобретают различные оттенки синего цвета, тогда как цитоплазматическая сеть между ними становится фиолетово-розовой.
Клетки типа «а» с началом питания, напротив, начинают быстро уменьшаться в размерах и сморщиваться в связи с исчезновением в них секреторных вакуолей.
Ядра их проявляют дегенеративные изменения, сильно сжимаются и приобретают способность к сплошному диффузному прокрашиванию по Фельгену (рис. 225). С середины стадии питания клетки описанного типа вообще не обнаруживаются или же выглядят небольшими комочками дегенерирующего материала на границе с выводными протоками (рис. 226).Форма и размеры клеток типа «с» в период питания существенно не меняются, так же как и структура их цитоплазмы. В полностью созревших альвеолах они оказываются сильно сжатыми между гипертрофированными клетками «Ь». В цитоплазме их не удается выявить никаких вакуолей или включений секреторной природы, ядра приобретают овальную форму и почти полностью теряют способность окрашиваться по Фельгену.
По мере освобождения от секрета альвеолы II типа заметно сжимаются. В клетках «Ь» внутри цитоплазмы появляются обширные пустые полости, ядра приобретают неправильную форму и постепенно растворяются. В дальнейшем клетки подвергаются полному лизису, причем из-за разновременности созревания клеток даже в пределах одной альвеолы у заканчивающих питание клещей в слюнных железах можно видеть все стадии секреторной активности и конечные этапы дегенерации (рис. 226). Разрушение альвеол II типа завершается в первые дни после окончания питания. Внутри желез в этот период появляется большое количество фагоцитов, захватывающих продукты лизиса клеток.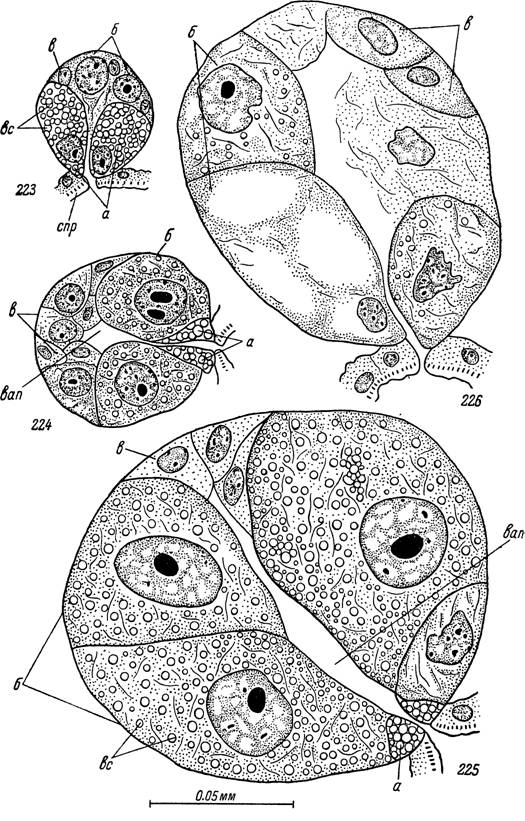
Рис. 223—226. Секреторные альвеолы II типа слюнных желез самки Ну а-
lomma asiaticum.
223 — голодный клещ; 224 — через двое, 225 — через пять суток после прикрепления к хозяину; 226 — полностью напитавшийся клещ сразу по
отпадении.
спр — стенки выводных протоков; вап — внутриальвеолярная полость; а — клетки типа «а»; б — клетки типа «Ь»; в — клетки типа «с»; вс — вакуоли с секретом.У голодных особей альвеолы III типа почти правильной сферической формы, диаметром в 30—40 мк. Внутри их находятся небольшие полости, сообщающиеся с выводными протоками слюнных желез.
В проксимальной половине альвеолы полость окружают 4—5 крупных клеток типа «d» (рис. 227). Они заполнены сферическими вакуолями с секретом, окрашивающимися азокармином, в отличие от клеток «а», в оранжево-красный цвет. Ядра сравнительно крупные (диаметр 9—12 мк), округлые или лопастевидные со сферическими нуклеолами и большим числом мелких и средних глыбок ДНК.Остальная часть альвеолы занята сравнительно мелкими клетками типа «е». Они пирамидальной формы, с вершиной, направленной в сторону просвета, и правильными сферическими ядрами диаметром в 6—9 мк. Объем цитоплазмы невелик, и в ней не удается обнаружить какие-либо секреторные вакуоли или гранулы.
Во время питания размеры альвеол III типа также значительно увеличиваются и их диаметр достигает 75—95 мк. Характерной их особенностью будет обширная внутренняя полость, формирующаяся за счет роста и уплощения клеток «е» (рис. 228, 229). На гистологических срезах в клетках данного типа начинают четко выявляться волокнистые структуры, а по достижении полного развития их апикальные концы, выступающие в просвет, приобретают зубчатую или неправильно изорванную форму (рис. 229). Местами происходит отторжение небольших участков апикальной цитоплазмы в полость альвеолы. Однако морфологически оформленные гранулы или вакуоли секрета в клетках «е» не обнаруживаются. По-види- мому, продуктом их секреции служит основное вещество цитоплазмы, поступающее в процессе медленного разрушения клеток в выводные протоки слюнных желез. Величина ядер в активно секретирующих клетках несколько увеличивается, они приобретают овальную форму и размеры их составляют 9—14x6—10 мк. Ядра хорошо прокрашиваются различными основными красителями, содержат много мелких и средних глыбок ДНК и сравнительно крупные овальные нуклеолы.
В клетках типа «d» основная масса секрета образуется еще до начала питания, но, в отличие от клеток «а», эти процессы продолжаются и в первые дни после прикрепления к хозяину. Объем цитоплазмы несколько увеличивается, и в ней продолжают появляться новые секреторные вакуоли. Выделение секрета в просвет железы продолжается на протяжении всего периода питания. Даже у заканчивающих кровососание самок в некоторых клетках сохраняются секреторные вакуоли (рис. 230). В цитоплазме таких клеток появляются обширные пустые полости, а ядра сморщиваются и подвергаются пикнотическим изменениям.
В заканчивающих цикл секреторной активности альвеолах клетки «е» сильно истончаются за счет излияния протоплазмы во внутренний просвет, ядра сморщиваются и почти полностью теряют способность окрашиваться по Фельгену. Стенки альвеолы спадаются, внутренняя полость уменьшается, а в дальнейшем, как и у альвеол II типа, происходит полная дегенерация.
У Rhipicephalus appendiculatus (Till, 1961) и Haemaphysalis spinigera (Chinery, 1965b), как и у Ы. asiaticum, в слюнных^ железах самок выявлены два аналогичных типа секреторных альвеол. Различия проявляются лишь в том, что у этих видов в альвеолах II типа только 2 разновидности секреторных клеток («а» и «Ь»), а в альвеолах III типа, наоборот, 3 («с», «d» и «е»). В то же время тонкое строение этих клеток и особенности их функционирования сходны у 3 изученных видов.
У самцов Hyalomma asiaticum, а также Rhipicephalus appendiculatus и Haemaphysalis spinigera обнаружены 4 типа альвеол. Альвеолы I типа, илие
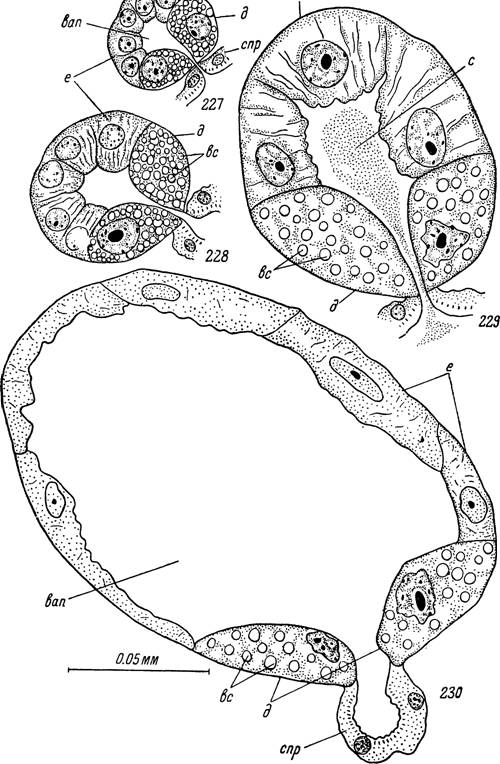
Рис. 227—230. Секреторные альвеолы III типа слюнных желез самки Hyalomma asiaticum.
227 — голодный клещ; 228 — через двое и 229 — через пять суток после прикрепления к хозяину; 230 — полностью напитавшийся клещ сразу по отпадении.спр — стенки выводных протоков; вап — внутриальвеолярная*полость; д — клетки типа «d»; е — клетки типа «е»; вс — вакуоли с секретом; с — секрет в полости альвеол.пирамидальные, обычного для иксодоидных клещей строения. Альвеолы II типа Hyalomma asiaticum образованы 2 клетками типа «а», 4—5 — типа «Ь» и 4—7 типа «с». Размеры альвеол голодных особей достигают 40x50 мк, а во время питания увеличиваются до 60x70 мк.
У голодных самцов клетки «а» заполнены вакуолями с секретом. Клетки «Ь» и «с» не содержат включений секреторной природы и практически не отличимы. Они занимают дистальную половину альвеолы, причем границы между клетками плохо различимы. Ядра сферические, диаметром 5—6 мк (рис. 231).
С началом питания клетки «а» освобождаются от секрета и спадаются. Большую часть альвеолы занимают гипертрофированные клетки «Ь». Их цитоплазма заполнена сетью волокнистых эндоплазматических структур, между которыми лежат многочисленные секреторные вакуоли, окрашивающиеся при азановой методике в синий цвет (рис. 232). Ядра увеличиваются до 18—19 мк, сохраняя сферическую форму. Клетки «с» представлены узкими прослойками цитоплазмы с мелкими вытянутыми ядрами. В процессе освобождения от секрета альвеолы сморщиваются, а цитоплазма клеток «Ь» перестает окрашиваться.
Альвеолы III типа, в отличие от таковых у самок, наряду с клетками «d» и «е» обладают еще одним типом железистых клеток — «f». Последние в количестве от 3 до 5 лежат между 1—2 клетками «d» и 5—8 клетками «е». Клетки «f» занимают большую часть альвеолы и, как и «d», заполнены уже у голодных особей крупными секреторными вакуолями (рис. 233). Последние, в отличие от клеток «d», окрашиваются азокармином не в красный, а в оранжевый цвет и не чернятся железным гематоксилином. Клетки «е» занимают узкий сектор вершины альвеолы. Цитоплазма их сильно волокнистая, но в ней не выявляются секреторные вакуоли.
Во время питания размеры альвеол III типа увеличиваются с 45 X 50 мк до 60х70мк, главным образом за счет клеток «f». Объем клеток «е» увеличивается незначительно, а диаметр ядер достигает 9 мк по сравнению с 6 мк у голодных. Превращение альвеол III типа в процессе освобождения от секрета в тонкостенные пузырьки, как у самок, не наблюдается (рис. 234).
Только у самцов найдены альвеолы IV типа. У голодных особей они имеют вид мелких пузырьков размером 28x31 мк из 7—10 кубических клеток, окружающих небольшую центральную полость, сообщающуюся с альвеолярным выводным протоком (рис. 235). Ядра сферические диаметром 7 мк. Какие-либо проявления секреторной активности на этой стадии отсутствуют.
Во время питания альвеолы этого типа достигают наиболее крупных размеров (230x240 мк). Клетки приобретают пирамидальную форму и тесно прижаты друг к другу, так что внутриальвеолярная полость исчезает. Большая часть цитоплазмы заполнена крупными сферическими секреторными вакуолями диаметром 3—6 мк, окрашивающимися азокармином в темно-красный цвет. Ядра увеличиваются до 32x45 мк и располагаются в узкой базальной цитоплазматической зоне, свободной от секреторных вакуолей (рис. 236). По окончании накопления секрета в цитоплазме ядра сморщиваются и подвергаются пикнотическим изменениям.
Рис. 231—236. Секреторные альвеолы слюнных желез самца Hyalomma asiaticum. 231 — альвеолы II типа у голодного клеща; 232 — то же у питающейся особи; 233 — альвеола III типа у голодного клеща; 234 — то же у питающейся особи; 235 — альвеола IV типа у голодного клеща; 236 — то же у питающейся особи.
а — клетки типа «а»; б — клетки типа «Ь»; в — клетки типа «с»; г — клетки типа «g»; д — клетки типа «d»; е — клетки типа «е»; ж — клетки типа «f»; вбс — вакуоли с белковым секретом; вус — вакуоли с углеводно-белковым секретом; к — кутикулярная выстилка стенок выводных протоков; вап — внутриальвеолярная полЛють.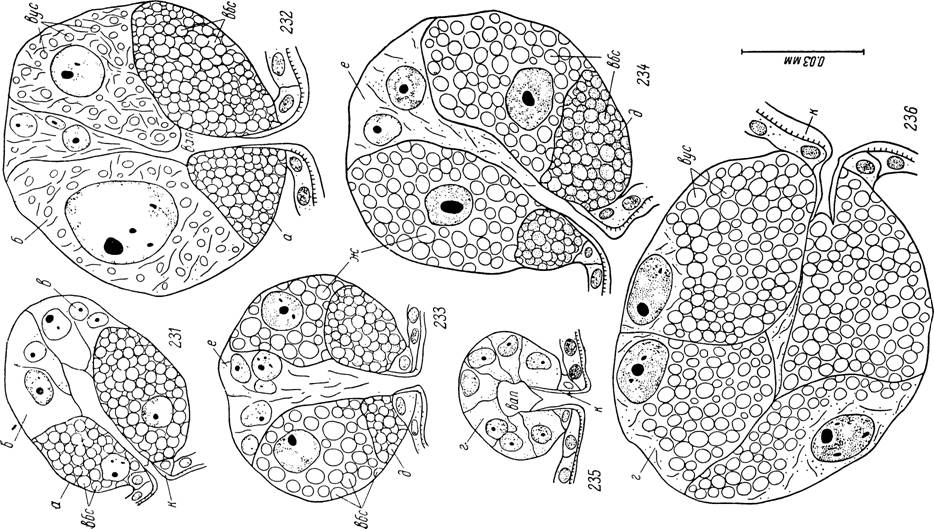 Ультраструктура секреторных клеток. При электронномикроскопиче-
Ультраструктура секреторных клеток. При электронномикроскопиче-
Еще по теме И к содовые клещи.:
- Вскрытие аргасовых и ик- содовых клещей.
- Гамазовые клещи (надсемейство Gamasoidea).
- Раздел IV КЛЕЩИ, ИМЕЮЩИЕ МЕДИЦИНСКОЕ ЗНАЧЕНИЕ
- Отряд Клещи
- КЛЕЩИ-КРАСНОТЕЛКИ (TrombicuIidae)
- ИКСОДОВЫЕ КЛЕЩИ (Ixodidae)
- Аргасовые клещи (сем. Argasidae).
- Глава IX ИКСОДОВЫЕ КЛЕЩИ И ВОЗБУДИТЕЛИ ИНФЕКЦИЙ
- Отряд Паразитоформные клещи - Parasitiformes.
- Иксодовые клещи (сем. Ixodidae).
- Надотряд КЛЕЩИ
- КЛЕЩИ И ВОЗБУДИТЕЛИ ТРАНСМИССИВНЫХ ИНФЕКЦИЙ ПОЗВОНОЧНЫХ
- Краснотелковые клещи (подотряд Trombidiformes).
- Отряд Акариформные клещи - Acariformes
- СЕМЕЙСТВО ИКСОДОВЫЕ КЛЕЩИ (IXODIDAE)
- Аргасовые клещи.
- Иксодовые клещи.
- КЛЕЩИ-КРАСНОТЕЛКИ