Слюнные железы
Слюнные железы таежного клеща, как и у других иксодид, отличаются большой сложностью строения и выполняют многообразные функции, связанные с процессами питания и осморегуляции. Наиболее полно их строение изучено у клещей А.
атегі- сапит (Krolak et al., 1982), В. microplus (Binnington, 1978; Megaw, Beadle, 1979), H. spi- nigera (Chinery, 1965, 1973), H. asiaticum (Атлас..., 1979), R. appendiculatus (Fawcett et al., 1981a, 1981b; Walker et al., 1985), /. holocyclus (Binnington, Stone, 1981),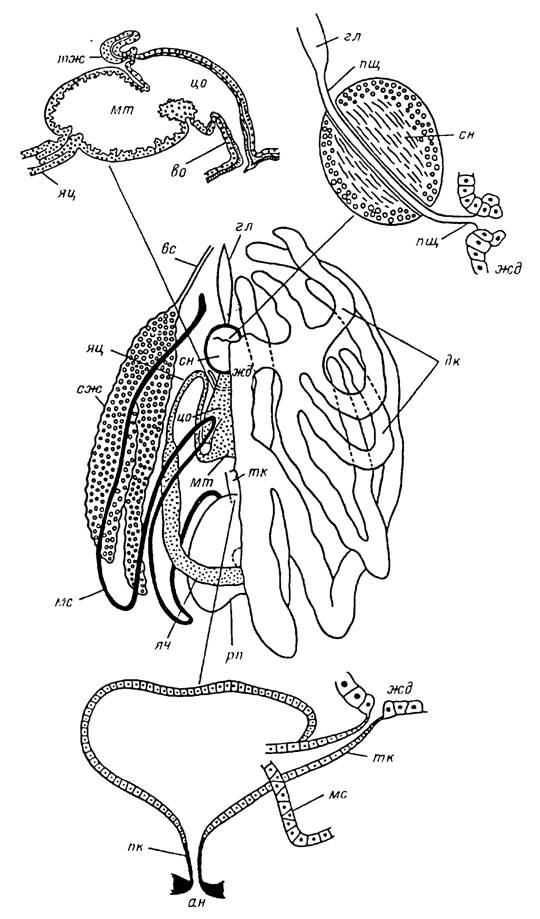
Рис. 9. Внутреннее строение самки Ixodes persulcatus.
ан — анальное отверстие; во — вестибулярный отдел влагалища; вс — выводной проток слюнной железы; гл — глотка; дк — дивертикулы средней кишки; жд — желудок; мс — мальпигиев сосуд; мт — матка; пк — прямая кишка; пщ — пищевод; рп — ректальный пузырь; сж — слюнная железа; сн — сннганглнй,- тж — трубчатая придаточная железа; тк — тонкая кишка; цо — цервикальный отдел влагалища; яц — яйцевод; ян — яичннк.
/. persulcatus (Балашов, 1979; Таежный клещ..., 1985). Все эти виды обладали сходным строением и клеточным составом слюнных желез при существовании небольших различий у представителей разных родов и особенно подсемейств Amblyommi- пае и Ixodinae.
Слюнные железы представлены парой массивных гроздевидных органов, занимающих латеральные части идиосомы от уровня заднего конца глотки до перитрем (рис. 9). У голодной самки в зависимости от видовой принадлежности их длина варьирует от 2.0—2.4 мм у /. persulcatus до 3—4 мм у Н. asiaticum. У напитавшихся самок их размеры увеличиваются соответственно до 3—4 и 6—8 мм. У нимф и личинок размеры слюнных желез в несколько раз меньше, чем у взрослых особей (рис. 10).
Каждая железа состоит из 2 главных лопастей, распадающихся на многие более мелкие дольки, и весь орган объединяет несколько сотен железистых пузырьков (альвеол).
Альвеолы сидят непосредственно на стенках главных выводных протоков желез (диаметр 50—100 мкм) и их многочисленных первичных (20—40), вторичных (15—25) и третичных (6—10) разветвлений (табл. XVIII, 7; XIX, 2). Конечные разветвления главных выводных протоков — третичные протоки несут по несколько десятков секреторных альвеол. Каждая альвеола открывается коротким альвеолярным протоком, который у альвеол II и III типов снабжен конусовидной клапанной структурой (рис. И). Изнутри полость протока выстлана кутикулярным слоем толщиной от 3—4 мкм в главных до 1 мкм в третичных протоках. В толще кутикулярной выстилки, по ее наружной поверхности, вдоль всей длины протока проходит спиральная нить из более твердого материала (табл. XVI, 2). По своему строению она сходна с аналогичной тенидиальной нитью в стенках трахей и, как и последняя, выполняет опорную функцию, предотвращая спадение стенок. Под кутикулярной выстилкой располагается слой эпителиальных клеток цилиндричес*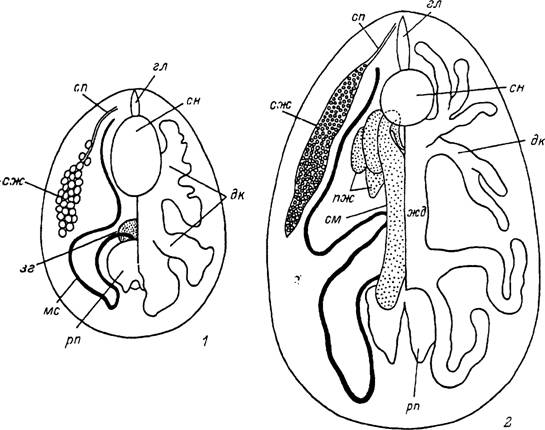
Рис. 10. Внутреннее строение личинки и самца Ixodes persulcatus.
I — личинка; 2 — самец, зг — недифференцированная закладка гонады; пж — комплекс половых придаточных желез; см — семенник; сп — слюнной проток. Остальные обозначения как на рис. 9.
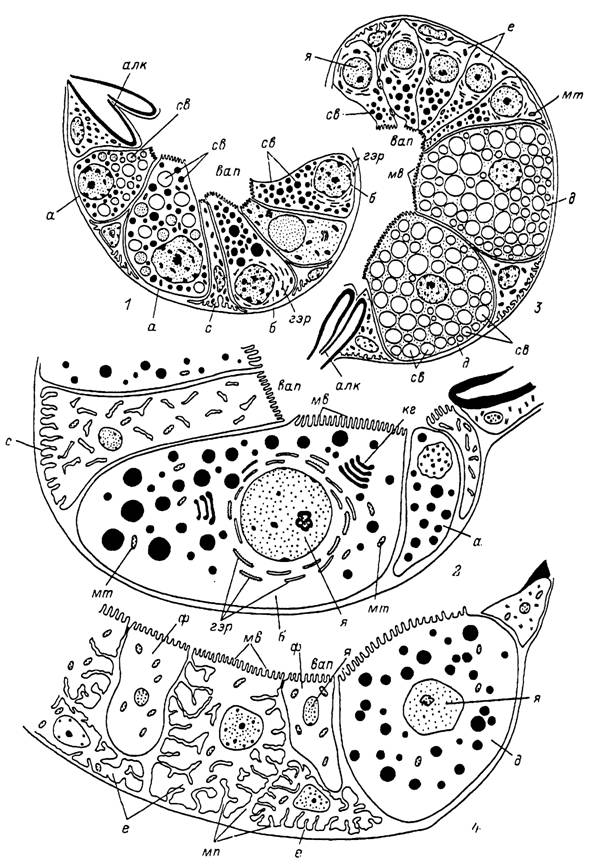
Рис. 11. Строение гранулосекретирующик альвеол самки Ixodes persulcatus.
[1] — альвеолы I типа голодной (7) и питающейся (2) особи; 3,4 — альвеолы II типа голодной (3) и питающейся (4) особи, г — клетка типа а; алк — альвеолярный клапан; б — клетки типа Ь\ в ап — внутри альвеолярная полость; гзр — гранулярный і-пдоплазматический ретикулум; д — клетки типа d\ е — клетки типа е; кг — комплекс Гольджи; мв — микроворсинки; — митохондрии; мп — межклеточные пространства; с — клетки типа с; св — секреторные включения; ф — клетки типа
/; я — ядро.кой или уплощенной формы, сходных по строению с гиподермальными клетками наружных покровов тела клеща.
Каждая из слюнных желез переплетена множеством трахей и пленок соединительной ткани.
Она иннервируется ответвлениями пальпального и I—III ножных нервов. Индивидуальные аксоны этих нервных окончаний входят внутрь каждой из секреторных альвеол, где находятся их синаптические зоны.У самок, нимф и личинок иксодид слюнные железы образованы 3 типами альвеол, у самцов амблиомин — 4 и у непитающихся самцов иксодин — 2 типами альвеол. В слюнных железах личинок, нимф и самок таежного клеща обнаружено 3 типа многоклеточных секреторных альвеол, а у самцов — 2 типа. Анатомическое строение слюнных желез сходно у всех иксодовых клещей. Имеются лишь небольшие межродовые различия у представителей подсемейств Amblyomminae и существенные отличия в наборах секреторных клеток и продуктах их секреции у амблиомин и иксодин.
Всем иксодовым клещам свойственны альвеолы I типа (пирамидальные, или не образующие гранул, альвеолы). Они встречаются только в передней части железы, где открываются непосредственно в главные выводные протоки или в их первичные ответвления (табл. XVI, 2). У самок Н. asiaticum в одной железе насчитывают 200—300 подобных альвеол и у самцов — около 150, а у /. persul- catus — соответственно 100—120 и 20—30. Строение этих альвеол не меняется во время питания, и их размеры составляют 50—70 мкм. В световом микроскопе эти альвеолы выглядят неклеточными органами с периферической волокнистой цитоплазмой и центральной зоной, заключающей одно крупное и несколько более мелких ядер. По данным электронной микроскопии (Barker et al., 1984), наружная цитоплазматическая зона этих альвеол представляет лабиринт из глубоких впячи.- ваний мембран периферических клеток (табл. XIX, 4). Эти структуры окружают единственную центральную клетку с крупным ядром. Предполагают, что в первые дни питания эти альвеолы выделяют липоидный секрет. Однако главная функция альвеол I типа связана с секрецией гипертоничной гигрофильной слюны для восполнения водного баланса в организме голодного клеща (Knulle, Rudolph, 1982).
Большую часть ткани слюнной железы образуют гранулосекретирующие альвеолы, названные так по признаку накопления в цитоплазме их железистых клеток крупных секреторных гранул.
В свою очередь входящие в состав этих альвеол железистые и интерстициальные клетки обозначают строчными буквами латинского алфавита (a, b, с, d, е, f g).Гранулосекретирующие альвеолы II типа встречаются в передней половине железы, расположены на главных выводных протоках и их первичных разветвлениях. Они лежат вперемежку с альвеолами III типа и по числу значительно уступают последним. У Н. asiaticum и других амблиомин они имеют овальную форму и их размеры у голодных самок 50x70 мкм (табл. XVI, 1, 3). Внутри они имеют вытянутую внутриальвеолярную полость. В каждой из желез самки Н. asiaticum от 200 до 300 альвеол II типа. Альвеола образована 3 типами секреторных клеток (а, ЬУ с). У голодных особей базальную часть альвеолы занимают несколько крупных клеток а, заполненных крупными секреторными гранулами. Меньшая по размерам апикальная часть образована несколькими мелкими, еще не дифференцированными клетками b и с. Секреторные клетки отделены друг от друга, от внутриальвеолярной полости и ог наружной оболочки альвеолы тонкими прослойками цитоплазмы интерстициальных клеток (аблюминальных и адлюминальных).
У питающихся клещей диаметр альвеол II достигает 140—200 мкм (табл. XVII, З, 4\ XVIII, 2, 3). С начала питания клетки а постепенно свобождаются от секреторных включений и спадаются. Клетки b и с с начала питания начинают увеличиваться в размерах. В цитоплазме их происходит синтез продуктов секреции, и она заполняется готовыми секреторными гранулами. В конце питания эти клетки становятся единственными гранулосекретирующими элементами альвеол II. Интерстициальные клетки альвеол питающихся клещей также увеличиваются в размерах и заполняют пространство между секреторными клетками системой цитоплазматического лабиринта-из тяжей и лакун.
У таежного клеща в альвеолах II обнаружено только 2 типа гранулосекрети- рующих клеток, и их гомологизация с соответствующими типами клеток амбли- омин затруднительна. В промежутках между секреторными клетками у них также располагаются сильно вытянутые интерстициальные клетки, отделяющие секреторные друг от друга.
Сам цикл секреторной активности сходен с описанным для амблиомин (рис. 11; табл. XIX, 2, 3). В первые дни питания полностью освобождаются от секрета базально расположенью клетки а, а клетки b приступают к накоплению и выделению секрета.Наиболее многочисленны в слюнных железах альвеолы III типа, число которых у самок Н. asiaticum достигает 1000—1200 и I. persulcatus — около 500. Из них построены задние 2/3 железы и дистальные боковые лопасти передней их трети. У голодных самок Н. asiaticum альвеолы III представляют собой почти сферические пузырьки диаметром 50—60 мкм (табл. XVI, 4\ XVIII, 7). Эти альвеолы образованы гранулосекретирующими клетками d, е, f и интерстициальными. Базальную часть альвеолы занимают несколько крупных клеток типа d, цитоплазма которых заполнена сферическими секреторными включениями. Далее располагаются несколько меньших по размерам клеток типа е и занимающих апикальное положение клеток /. У голодных особей два последних типа клеток свободны от секреторных гранул. Друг от друга секреторные клетки отделены узкими прослойками из интерстициальных клеток.
С начала питания альвеолы III быстро увеличиваются в размерах и на 3—4-е сутки достигают своей окончательной величины, превращаясь в тонкостенные пузырьки (табл. XVII, 7, 2; XVIII, 4, 5). У питающихся особей клетки d и е освобождаются от секреторных гранул и дегенерируют. Клетки е после освобождения от секреторных гранул вступают в стадию интенсивного роста, приобретают форму сильно уплощенных пирамид и образуют стенки у сильно увеличившейся в размерах внутриальвеолярной полости. Промежутки между этими клетками занимают также сильно увеличившиеся в размерах интерстициальные клетки. В цитоплазме клеток / образуется лабиринт из переплетающихся цитоплазматических тяжей и межклеточных полостей. Вместе с имеющими сходное строение интерстициальными клетками лабиринт из полостей и тяжей формирует клеточную стенку, ограничивающую обширную внутриальвеолярную полость от гемолимфы. Предполагают, что на этой стадии главной функцией альвеол III становится регуляция водно-солевого баланса в организме питающихся клещей.
Альвеолы III осуществляют активный транспорт избыточной воды и солей из гемолимфы внутрь альвеол и далее вместе со слюной в организм хозяина.Сходным образом функционируют и альвеолы III типа у таежного клеща (рис. 11; табл. XIX, 7). Из образующих их трех типов секреторных клеток гомологичны таковым амблиомин только клетки /, осуществляющие транспорт воды и солей. В строении и гистохимических особенностях двух других типов секреторных клеток между иксодинами и амблиоминами наблюдаются определенные отличия (Таежный клещ..., 1985).
У самцов Н. asiaticum кроме альвеол I, II и III типов имеется также около 300—400 альвеол IV типа, являющихся наиболее многочисленными в их слюнных железах. У голодных особей они свободны от секреторных включений. У питающихся клещей видно, что они образованы единственным типом клеток — g, цитоплазма которых заполнена большим количеством секреторных гранул. Эти клетки активны на протяжении всего многодневного периода питания. Для них характерно совмещение процессов синтеза новых секреторных гранул и их освобождения в полость альвеолы. У самцов 7. persulcatus секреторные клетки альвеол заполнены секреторными гранулами вскоре после линьки и для их функционирования нет необходимости в предварительном питании.
Во время питания со слюной выводится большое количество биологически активных компонентов, воды и солей. Объем слюны, выделяемой клещом за весь период питания, значительно превышает таковой тела напитавшейся особи (Kaufman, 1989), и путем модуляции состава и объема выделяемой слюны клещ не только поддерживает гомеостаз в своем организме, но и регулирует свои отношения с хозяином. В слюне клещей обнаружены компоненты цементного футляра, образующегося между кожей хозяина и ротовыми органами клеща (см. главу V). В ней выявлены несколько энзимов и их ингибиторов, стимуляторы выделения гистамина тучными клетками хозяина и антагонисты гистамина, антигемостатические факторы, простагландины, паралитические токсины, факторы развития и подавления иммунных реакций хозяина (Sauer et al., 1995).
На протяжении каждой фазы развития слюнные железы проходят закономерный цикл развития, включающий состояние покоя у голодных особей, интенсивного роста и секреции у питающихся клещей и дегенерации старых альвеол по окончании питания. Например, у самок А. атегісапит за время питания масса слюнных желез и содержание в них белков возрастают в 25 раз и более (Sauer et al., 1995). На стадии линьки на стенках выводных пртоков слюнных желез происходит формирование новых альвеол следующей фазы развития.
Процесс слюноотделения у питающихся особей находится под допаминергичес- ким нервным контролем. Выделение слюны можно искусственно стимулировать у недавно снятых с хозяина недопитавшихся самок при введении им интрацеломаль- но пилокарпина, допамина и других медиаторов нейрогенного типа, стимулирующих активность окончаний нейронов в клетках слюнных желез. Использование этой методики позволило проследить интенсивность выделения слюны самками таежного клеща в разные периоды питания. Оказалось, что наиболее интенсивное слюноотделение происходит в последние сутки питания, когда «водяные клетки» альвеол III типа выделяют особенно большое количество воды и солей, поступающих в гемолимфу в конце питания клеща.
Еще по теме Слюнные железы:
- Тема занятия. БОЛЕЗНИ ЭНДОКРИННЫХ ЖЕЛЕЗ. САХАРНЫЙ ДИАБЕТ. ЗАБОЛЕВАНИЯ ЩИТОВИДНОЙ ЖЕЛЕЗЫ. ОПУХОЛИ ЭНДОКРИННЫХ ЖЕЛЕЗ
- 1.6. Злокачественные опухоли слюнных желез
- 11. Рак грудной железы у мужчин? Лечение рака грудной железы у мужчин?
- Анатомо-морфологические особенности застенных слюнных желез жвачных
- Молочная железа
- Поджелудочная железа.
- СТРОЕНИЕ СЛЮННЫХ ЖЕЛЕЗ
- Поджелудочная железа
- Депонирование железа.
- 1.Нормальная анатомия поджелудочной железы.
- Рост массы слюнных желез
- Железо – основа клеточного дыхания
- ОПУХОЛИ СЛЮННЫХ ЖЕЛЕЗ
- 1.Рак поджелудочной железы.
- Транспорт железа в организме.
- 3.4. Поджелудочная железа.
- 73. Аномалии развития слезной железы
- 1. Пероральное назначение препаратов железа