1.3. Физиологические функции белковых продуктов генов-кандидатов подверженности туберкулезу, их роль в патогенезе заболевания
Одним из генов предрасполагающих к развитию туберкулеза является NRAMP1 (от англ. Natural-Resistance-Associated Macrophage Protein 1 gene – ген макрофагального протеина 1, ассоциированного с естественной резистентностью).
Более того, R. Bellamy и соавт. (1998) отнесли NRAMP1 к основным кандидатным генам туберкулеза у человека. Белковый продукт этого гена имеет вес около 60 кД, он локализован в лизосомальном компартменте покоящегося макрофага, но во время фагоцитоза он работает на мембране фагосомы [Gruenheid S. et al., 1997]. Nramp1 участвует в процессах активации макрофагов, являясь ключевым звеном в механизме транспорта нитритов из внутриклеточных компартментов в более кислую среду фаголизосомы, где он способен вступать в химическую реакцию с образованием NO [Blackwell J. M., Searle S., 1999]. Белок входит в семейство функционально связанных мембранных белков (к этому семейству относят также Nramp2), ответственных за транспорт двухвалентных катионов, таких как Fe2+, Mn2+, Zn2+, Cu2+ [Jabado N. et al., 2000; Cellier M. et al., 2001]. Известно, что ионы металлов являются жизненно важными элементами, участвующими во многих метаболических реакциях, происходящих в каждой живой клетке. Следовательно, недостаток, избыток или отсутствие данных элементов может привести к развитию какого-либо патологического состояния или даже к гибели клетки. Постоянство ионов металлов в организме обеспечивается регуляцией их потребления, хранения и выведения. Для того чтобы поддерживалась необходимая концентрация ионов, каждая клетка обладает определенной системой, обеспечивающей транспорт веществ через мембрану. Сбой этой системы или ее части может повлечь за собой потерю равновесия между выведением и поступлением веществ, что приведет к изменению внутриклеточной концентрации ионов. Недостаточный транспорт ионов может оказаться причиной нехватки жизненно важных метаболических элементов, а чрезмерное их накопление может вызвать токсическое воздействие этих же веществ, ведущее к гибели клетки. Возможно, что антибактериальная функция Nramp1 заключается в создании неблагоприятной для бактерии окружающей среды внутри фагосомы [Gruenheid S. et al., 2000; Barton C.H. et al, 1999]. Во время фагоцитоза микроба макрофаг продуцирует активные кислородные метаболиты, которые являются токсичными для бактерии. Выживание патогена во время кислородозависимой перестройки метаболизма фагоцита обеспечивается микробными ферментами, большинство из которых содержат ионы металлов в своих активных центрах [Cellier M. et al., 1994]. В свою очередь истощение запаса ионов металлов в фагосоме, вызванное транспортной деятельностью макрофагального белка ассоциированного с естественной резистентностью, приводит к снижению продукции металлосодержещих ферментов поглощенной бактерией. Следовательно, дефекты продукции или функции Nramp1 могут приводить к нарушению его транспортной функции и, как следствие, к повышению чувствительности к внутриклеточным патогенам, таким как микобактерии (рис. 1) [Barton C.H. et al., 1999].
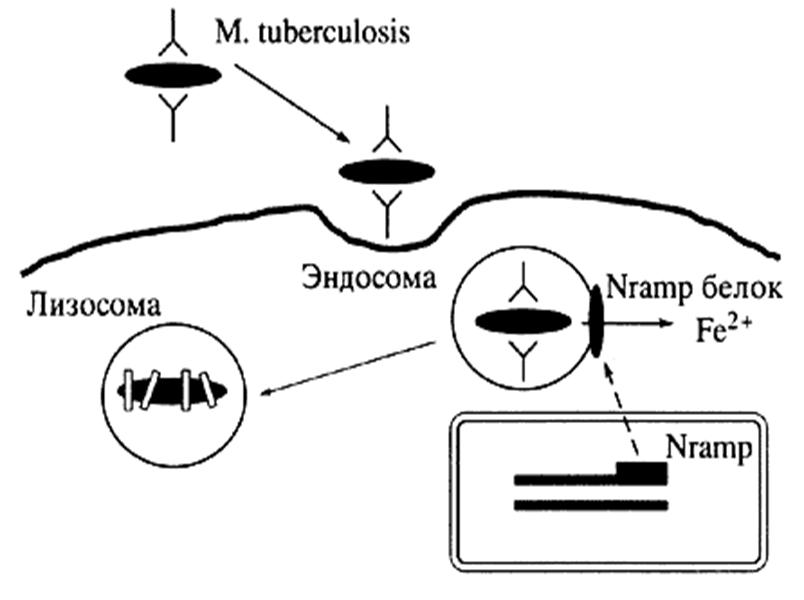 |
Рис. 1. Схема антибактериального действия NRAMP1 [по Пальцеву М.А., 2002]
Опыты, проведенные на инбредных мышах, показали, что уровень естественной резистентности к внутривенному заражению низкими дозами M. bovis (BCG) контролируется одним геном, локализованным в проксимальном регионе мышиной хромосомы 1. Этот локус обозначили как Bcg (также он известен как Lsh или Ity). Два различных фенотипа Bcg были ассоциированны с чувствительностью (Bcg-s) и с резистентностью (Bcg-r) на ранней стадии инфекции, вызванной M. bovis, M. avium, M. lepraemurium, Leishmania donovani, Salmonella typhimurium [Bredley D.J., 1977; Forget A. et al., 1981].
Экспериментальные исследования показали, что через 3 недели после заражения 10 КОЕ M.
bovis (BCG) из селезенки мышей Bcg-s высевается на 3-4 порядка больше микобактерий, чем из селезенки мышей Bcg-r [Gros P. et al., 1981].Результаты исследований на моделях мышей позволили утверждать, что высокая чувствительность линий мышей Bcg/Lsh/Ity к заражению внутриклеточными патогенами объясняется дефектом локализованного на 1-ой хромосоме гена в локусе Bcg [Blackwell S.M. et al., 1994; Skamene E., 1994].При помощи позиционного клонирования изолировали кандидатный ген и обозначили его как Nramp1 [Vidal S.M. et al., 1993]. Позже было подтверждено, что Nramp1 и ген, расположенный в локусе Bcg, идентичны [Govoni G. et al., 1996]. У лабораторных мышей ген Nramp1 имеет 2 аллеля Nramp1-s (восприимчивый, рецессивный) и Nramp1-r (резистентный, доминантный) [Malo D. et al., 1993].
Секвенирование матричной РНК Nramp1 от восприимчивых и резистентных линий мышей показало, что подверженность к инфекции связана с заменой глицина на аспарагиновую кислоту в позиции 169 (G169D) внутри 4-ого трансмембранного домена белка [Malo D. et al., 1994]. Элиминация функции Nramp1 у «нокаутированных» мышей (Nramp1-/-) приводит к повышению восприимчивости к группе бактериальных возбудителей, хорошо адаптированных к выживанию в макрофаге [Govoni G., Gros P., 1998].
Однако нельзя не учитывать, что в выше перечисленных экспериментах на мышах использовался штамм M. bovis (BCG), а он является авирулентным для человека. Более того, E. Medina и R. North (1998) показали, что в то время как Nramp1 действительно контролирует резистентность мышей к заражению M. bovis, резистентность к заражению M. tuberculosis, вероятно, не связана с мутациями данного локуса. Мыши с мутантным (чувствительным к заражению M. bovis) фенотипом не отличались по чувствительности к заражению M. tuberculosis от мышей с резистентным (дикого типа) фенотипом.
Учитывая полученные результаты, G. Govoni и P. Gros (1998) сделали вывод, что возбудители, не попадающие под контроль Nramp1, либо отличаются своим поведением внутри макрофага, либо не являются внутриклеточными паразитами. Эти данные свидетельствуют, что Nramp1 играет важную роль в резистентности к микобактериям и некоторым другим возбудителям инфекций у мышей, а его человеческий гомолог, вероятно, связан с подобными инфекциями у людей.
Такой человеческий гомолог гена Nramp1, обозначенный как NRAMP1, клонировали и картировали на человеческой хромосоме 2q 35 [Cellier M. et al., 1996]. В данном гене содержится 15 экзонов различной протяженности, разделенных интронами, размер которых также широко варьирует [Marquet S. et al., 2000]. Описано 9 полиморфных вариантов гена NRAMP1, которые, вероятно, влияют на функцию гена [Liu J. et al., 1995].
С целью изучения функции гена было проведено исследование различных полиморфизмов NRAMP1 у западных африканцев в Гамбии всвязи с туберкулезом в местной популяции. Четыре полиморфизма гена - 5`(CA)n, INT4, D543N, 3`UTR были ассоциированы с туберкулезом (р=0,03; р=0,009; р=0,008; р
Еще по теме 1.3. Физиологические функции белковых продуктов генов-кандидатов подверженности туберкулезу, их роль в патогенезе заболевания:
- Никотиновые ацетилхолиновые рецепторы: физиологические функции и роль в нейродегенеративных процессах
- Роль РНК-связывающих белков в нейродегенеративных заболеваниях на примере БАС-FUS
- Ключевые функции организма человека, позитивное воздействие на которые позволяет относить продукты к категории продуктов функционального питания:
- 1.2. Роль микроциркуляторных расстройств и дисфункции сосудистого эндотелия в патогенезе кардиометаболических заболеваний
- 3.4.2. Анализ ассоциаций исследованных генов с количественными признаками туберкулеза
- Нарушения образования и выведения конечных продуктов белкового обмена
- Переработка денуклеинизированной микробной биомассы с получением продуктов белковой природы
- 3.2. Анализ связи полиморфизма генов NRAMP1, IL12B, VDR, IL1B, IL1RN с туберкулезом
- 3.4.1. Анализ ассоциаций исследованных генов с качественными признаками туберкулеза
- Анализ ассоциации полиморфных вариантов генов-кандидатов с параноидной шизофренией и ответом на типичные нейролептики
- Роль фосфорилирования транспортных белков в нейронах
- Основные функции и структура белков