КОЛИЧЕСТВЕННАЯ ХАРАКТЕРИСТИКА И ЭНЕРГЕТИКА ПОВЕДЕНИЯ
Внимание к этой стороне вопроса было привлечено в связи с изучением суточного ритма двигательной активности и влияния на него различных факторов. При этом оказалось возможным изучение суммарной двигательной активности животного за определенный отрезок времени и распределение этой активности на протяжении суток (Szymanski, 1918, 1920; Щербакова, 1937, 1938, 1949; Калабухов, 1938, 1940, 1950; Черкович, 1950, 1958 и мн.
др.).Большой фактический материал позволяет утверждать, что общая суммарная двигательная активность (ходьба, бег, прыжки) и их энергетический эквивалент остаются при одинаковых условиях у отдельных особей изо дня в день постоянными. Это впервые было показано О. П. Щербаковой (19496) на обезьянах, причем удалось отметить определенные взаимоотношения между горизонтальными прыжками и вертикальными. Общая величина выполненной работы в кг!м и ,н/кг при одинаковых условиях исследования (температура среды, освещение) оставалась постоянной. Суммарная активность животного и ее энергетический эквивалент являются довольно постоянным признаком, характеризующим данный вид и даже данную особь. На высших обезьянах — шимпанзе и на лемурах подобные отношения в течение суток были установлены Р. С. Персон и Н. Е. Мярков- скон (1953).
Тот факт, что суммарный энергетический эквивалент мышечной работы, выполняемой животным в течение суток, остается постоянным, заставил обратиться к изучению влияния навязанных мышечных нагрузок на спонтанную деятельность животного. Первым исследованием на эту тему явилась работа М. Р. Майзелис (1953), проведенная на больших песчанках. Оказалось, что непроизвольная, даже кратковременная, мышечная работа значительно сокращает спонтанную активность животного. Это уменьшение активности может продолжаться несколько суток; наблюдается снижение газообмена и снижение веса в дни сокращенной двигательной активности.
На фоне общего снижения двигательной активности выявились некоторые ее особенности. В течение суток можно было наблюдать определенные циклы, характеризующие такие движения, как бег и ходьба, питание и чистка меха; чередование этих элементов оставалось неизменным. Сокращалось только их время и увеличивались промежуточные периоды покоя. Некоторые эле-
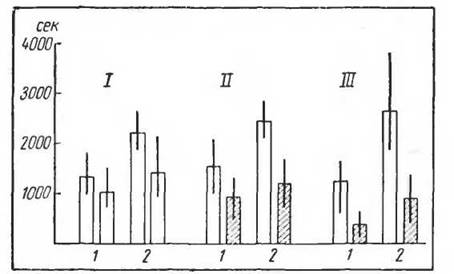
Рис. 16. Длительность игровой активности золотистых хомяков (по Склярчик, 1965). / — контроль; // — бег в течение 600 мин\ III—-бег в течение 3600 мин. Белые столбики— до бегл в третбане; заштрихованные—- после бега в третбане. Линин — размах» колебаний (минимум и максимум), столбики— средние величины:
1 — дни с большой длительностью игровой актив ности в первом исследовании. 2 — дни е меньшей длительностью игровой активности в первом ис
следовании
менты активности, связанные с питанием, мочеиспусканием, дефекацией, существенно не изменились. Впоследствии А. Г. Понугаева и Е. Л. Склярчик установили такие же отношения в изменении поведения после мышечных нагрузок и по отношению к циклам игры у молодых животных (золотистые хомяки) (рис. 16).
Следовательно, в поведении животного можно различать как облигатные, так и факультативные для каждого вида элементы. Облигатные элементы не могут быть изменены даже количественно (конечно, у нормальных особей данного вида); они характеризуются своеобразием не только качественным, но даже и количественным — удельным весом в общей двигательной активности
животного. По-видимому, пищевые рефлексы и пшцедо- бывапие не могут всегда считаться облигатными элементами поведения. Это видно хотя бы из опытов В. Г. Кассиля, А. М. Уголева и В. Н. Черниговского (Черниговский, 1962), установивших, что в зависимости от концентрации раствора и его состава (глюкоза 8% и 40%-ные растворы), а также в зависимости от состояния пищевой возбудимости количество выпитого раствора меняется.
При этом 40%-ный раствор глюкозы использовался как пищевое вещество. Значит, акт питья может изменяться в зависимости от состояния пищевой возбудимости и наоборот.Рассмотрение полученного материала убеждает в том, что между временем еды и калорийностью съеденного корма существуют обратные соотношения. Чем ниже калорийность пищи, тем больше время еды. При расчете на 1 кг веса тела \ растительноядных травоядных форм поступает 0,04—0,14 кал!кг/мин, у плотоядных 1,03— 9,66 кал/кг/мин, у всеядных 1,4—5,5 кал/кг/мин. Такова энергетическая основа пищевого поведения с количественной стороны. Разумеется, эти отношения, полученные в условиях неволи, совершенно элиминируют природную пищедобывательную деятельность. Однако наблюдения в условиях пастбища свидетельствуют также об определенных отношениях времени пшцедобывания у разных видов в разных условиях (Трайб, 1959; Слоним, 1962).
Таким образом, количественная сторона поведения определяется н энергетическими отношениями, отношениями поверхности тела и, по-видимому, именно эти условия определяют видовое постоянство.
Количественная характеристика поведения как общего (двигательного), так и специального (пищевого, пи- щедобывательного) в значительной мере зависит от температуры внешней среды. Это хорошо выявлено во многих исследованиях пастьбы сельскохозяйственных животных (Трайб, 1959). В лабораторных условиях изменения таких врожденных форм поведения, как роющая деятельность, грызение под влиянием температуры внешней среды, выявились в работах А. Г. Понугаевой (1960) и А. И. Щегловой (1958). Для грызущей деятельности большой песчанки, например, установлена оптимальная зона температур +20—22° С, при которых животные сгрызают наибольшее количество древесины. Высокие
температуры тормозят роющую деятельность у грызунов различных видов. Следовательно, как для общей двигательной активности, так и для специализированных ее форм существует определенный температурный оптимум.
Каков же физиологический механизм, лежащий в основе этого оптимума? Многие исследователи наблюдали сокращение времени пастьбы и сокращение времени всей пищедобывательной деятельности сельскохозяйственных животных.
Это обычно объясняют необходимостью снизить теплопродукцию в условиях нагревания, что и достигается уменьшением приема пищи. Возможно также, что повышенная теплопродукция во время двигательной активности является фактором, повышающим температуру тела, н в порядке регуляции теплового баланса наступает разлитое торможение в центральной нервной системе, ограничивающее двигательную активность. Пъ- видимому, между температурой внешней среды и величиной двигательной активности гомойотермного организма существуют более сложные отношения.А. А. Алимухамедовым (1960) и А. А. Алимухамедо- вым и К. П. Ивановым (1963) было установлено, что температурная реакция проявляется ранее, нежели развивается собственно терморегуляция. Новорожденные грызуны, взятые из гнезда, где они находились с матерью, реагируют на небольшое охлаждение сначала повышением произвольной двигательной активности, а затем ее понижением. В последующие после рождения дни картина резко меняется (рис. 17). Начальная фаза повышения обмена при охлаждении все более растягивается во времени, а затем тепло, которое в первые дни и часы после рождения вызывало возбуждение животного, начинает его угнетать. Появляется реакция, свойственная взрослому гомойотермному животному,— реакция, которая входит как обязательный компонент в поддержание температуры тела (терморегуляцию) в естественных условиях жизни. Очень трудно согласиться с высказываниями некоторых исследователей (Hart, 1960; Scholander, 1960), указывающими на то, что способность к поддержанию температуры тела при охлаждении следует рассматривать как явление адаптации только в состоянии полного покоя животного. Многие гомойотерм- ные организмы практически не остаются в покое длительное время (мышевидные грызуны, землеройки). Для
многих видов небольшая двигательная активность является как бы фоновой. Поэтому теплообразование при мышечной деятельности, особенно при умеренной, постоянно входит как неизбежная составная часть в общий энергетический баланс организма.
С этой стороны представляет значительный интерес влияние температуры среды на так называемую спонтан-
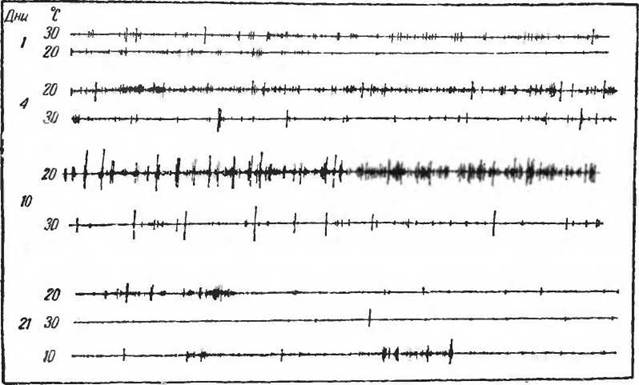
Рис. 17. Двигательная активность белой крысы при разных температурах среды в первые дни после рождения (по Алпмухамедову, 1961)
ную деятельность молодого животного, которая хорошо известна даже стороннему наблюдателю под общим названием игры, или игровой деятельности. А. Г. Попугаевой было показано, что интенсивность игры, изучаемая количественно (время игры/час), в процессе развития имеет характерную кривую. У молодняка золотистого хомяка игра начинается на 15—18-й день, достигает максимума на 26—28-й день и резко снижается на 36-й день. У других видов грызунов эти явления несколько более растянуты. Как показывают сопоставления данных, полученных А. Г. Понугаевой и Д. А. Рашевской, интенсивности игровой деятельности соответствуют в онтогенезе и потребление кислорода, и величины систолического и минутного объема сердца. Последние две величины опре-
делились как прямым путем но Фику, так и расчетом так называемого кислородного пульса (см. рис. 10).
Исследования А. Г. Попугаевой показали, что при содержании помета хомяка при температуре среды + 10— 15° С на игровую деятельность приходилось около 85% от общего времени опыта; у помета хомяков, содержавшихся при температуре +23—25° С, эта величина составляла только 65%.
Весьма важные данные получены в термограднент- ириборе при изучении игры. «Холодные» хомячки, содержавшиеся при температуре +10—15°С, играли в температурной зоне +12—16° С. «Теплые» хомячки, содержавшиеся при +23—25° С, играли в зоне +17—34° С. Интересно, что после мышечной работы — игры как теплые, так и холодные животные располагались и засыпали в термоградиентприборе при одинаковой температуре +22—23° С. Следовательно, предварительное содержание молодняка в различных температурах не только оказывает влияние на уровень спонтанной активности, но и позволяет выявить существенные различия в термопреферен- думе у этих животных.
Выполненная мышечная работа резко снижает термическую чувствительность организма (Слоним, 1952, 1962, 1962). Низкая температура среды (известный термический градиент теплоотдачи) является одним из важных факторов, стимулирующих мышечную активность. II хотя некоторые исследователи (Hardy, 1961; Hart, 1962) считают, что мышечная дрожь по своему калорнгенному эффекту более эффективна, нежели произвольные мышечные движения, она только в случаях крайне интенсивного охлаждения используется животными (едва ли кто-нибудь сознательно доведет себя до дрожи и откажется сделать несколько энергичных движений, если ему придется некоторое время пробыть на холоде). В этих условиях и человек, и животные начинают двигаться, и снижение температурной чувствительности играет, по-видимому, немаловажную роль.Изложенные выше данные, а также специальные исследования соотношений между температурой тела и явлениями утомления при мышечной работе животных (Склярчик, 1963) позволяют высказать гипотезу о термостатической регуляции мышечной работоспособности и интенсивности двигательной активности животных. Во всяком случае, для общей количественной оценки двигательного поведения животных температура внешней среды имеет преимущественное значение как в момент самого исследования, так и в предшествующий исследованию период (температурная адаптация).
Однако как стимуляцию, так и угнетение общей двигательной активности можно наблюдать не только при воздействии разных температур среды. А. И. Щегловой было установлено, что ограничение дачи воды и снижение содержания воды в корме резко повышает двигательную активность у пустынного грызуна большой песчанки. При этом распределение двигательной активности ее в течение суток не изменяется (песчанка — животное с дневной активностью). У ночного вида — полуденной песчанки (Pallasiotnys mcridianus) содержание иа такой скудной ио содержанию воды пище вызывает значительное снижение активности. Такие же данные были получены А. И. Щегловой и у лабораторных крыс. Ограничение двигательной активности при сухоядении у желтого суслика наблюдала Л. Г. Филатова (1944).
Таким образом, такой казалось бы неспецифический раздражитель, как изменение осмотической концентрации крови и тканей, вызывает совершенно различные изменения поведения у животных с разной экологической специализацией. Если ограничение движений в случае с желтым сусликом является пассивной адаптацией, вызывающей экономию потери воды, то увеличение активности у дневного вида большой песчанки, наоборот, связано, вероятно, с миграцией или увеличением роющей деятельности, что приводит к добыче частей растений, содержащих воду.
Количественное изучение поведения животного, проведенное в различных условиях, может дать очень много для понимания биологической значимости отдельных его форм, а иногда и вскрыть отдельные физиологические механизмы, лежащие в его основе. Однако только одна количественная характеристика не дает возможности вскрыть ряд зависимостей поведения различных видов от их экологической специализации. В этом плане значительные услуги оказывает исследование структуры отдельных элементов поведения — рефлекторных актов, включающих как двигательные, так и вегетативные функции.
4—1204
Еще по теме КОЛИЧЕСТВЕННАЯ ХАРАКТЕРИСТИКА И ЭНЕРГЕТИКА ПОВЕДЕНИЯ:
- 25. Количественные и качественные характеристики социальных отклонений.
- 2. Количественные ультразвуковые характеристики нормальной молочной железы в возрастном аспекте.
- Основные количественные характеристики сенсорных систем человека
- 32.Характеристика просоциального поведения личности.
- Условнорефлекторная характеристика трудового поведения человека.
- 1.Общая характеристика зависимого поведения.
- 17.Психологическая характеристика личности, склонной к успешному поведению.
- 17.Психологическая характеристика личности, склонной к успешному поведению.
- 31. Неактивные формы поведения животных: общая характеристика.
- 9.Характеристика уровня психической регуляции поведения и деятельности: произвольная, непроизвольная, волевая
- 2.2. Общие характеристики подходов и теорий поведения личности в организации.
- P.O.T. Влияние характеристик социальных сетей потребителей инъекционных наркоти- ков на рискованность их поведения
- 21. Делинквентное поведение как форма отклоняющегося поведения личности.
- 1. Предмет и содержание дисциплины «Организационное поведение». Модели поведения человека в организации
- 17. Понятие «девиантное поведение». Критерии определения понятия. Особенности отклоняющегося поведения личности.
- 8. Поведение. Личностное поведение.
- 6. Наследственность и среда – основные факторы количественной изменчивости.
- Количественный подход
- Количественная оценка ионизирующих излучений. Основы дозиметрии