ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ НЫРЯЮЩИХ ОРГАНИЗМОВ
Физиологические исследования ныряющих организмов, в частности млекопитающих, касаются главным образом особенностей их легочного дыхания, естественно прекращающегося на продолжительное время во время ныряния.
Способность нырять довольно характерна для различных видов млекопитающих. Продолжительность пребывания млекопитающих под водой колеблется от 50 сек до 2 ч (Irving, 1939) (табл. 23).Продолжительность ныряния млекопитающих (по Jrving, 1939; Harrison, 1960)
| Объс кт | Время | Объект | Время |
| Утконос (Ornithor- hynchus anatitius) . . | 10 мин | Кашалот (Physcfer catodon) .......................... | 1 ч — 1 ч |
| Морской слон (На- lichocrus grypus) . . | 6 MUH | Бутылконос (Ну- | 15 мин |
| Нерпа (Pysa hispid а) ....................................... | 48 сек | peroodon ampullatus) | 2 ч |
| 15 мин | Гренландский кит (Ba.la.ena mysticetus) | 1 ч | |
| Тюлень-тевяк (На- lichoerus grypus . . | 15 мин | Блювал (Balaenop- tera mu scut us) . . . | 49 мин |
| Водяная крыса (Аг- vicola terrestris) . . | 2 мин | Японский кит (Eu- balacna glacialis) . . | 50 мин |
| Ондатра (Ondatra zibethica) | 17 сек 12 мин | Большой полосатик (Balaenoptera niuscu- lus) ......................................... | 30 мин |
| Бобр (Castor fiber) | 15 мин | Финвал (Balaenop- | 20 мин |
| Кролик (Oryctola- | tcra nodosa) . . . . | ||
| gus cuniculus) . . . | 3 мин | Горбатый кит (Me- | 30 мин |
| Собака домашняя | 4 мин | gaptcra nodosa) . . | |
| Бегемот (Hippopo- | 25 сек | Человек (искатели жемчуга и губок) | 2 мин |
| tamus amphibius) . . | 50 сек | 30 сек | |
| Морж (Odobcnus rosmarus) . ... , | 16 мин 20 егк |
Сравнение приведенных величин показывает, что наибольшей устойчивостью к прекращению дыхания обладают китообразные: некоторые из них способны находиться под водой от одного до двух часов. Насекомоядные (выхухоль), грызуны (ондатра), а также ластоногие (тюлени) способны оставаться под водой очень ограниченное время.
Среди водных млекопитающих, способных к нырянию, т. е. обладающих повышенной устойчивостью к гипоксии, можно встретить представителей многих отрядов. Так, из низших млекопитающих пыряет утконос, из насекомоядных— выхухоль, из грызунов — водяная крыса, ондатра, нутрия, бобр, нз наземных хищных — выдра, из нежвачных парнокопытных — бегемот. Ныряют, разумеет- 262
ся, все ластоногие и китообразные и ныне почти вымершие сиреповые.
Физиологические особенности пыряющих животных касаются изменении дыхательного аппарата, изменении дыхательной функции крови, содержания миогемоглобина, а также некоторых особенностей регуляции дыхания н кровообращения в условиях гипоксии.
/Морфологические отличия легких заключаются в том, что объем их у пыряющих организмов значительно превосходит объем легких у наземных форм. Например, у дельфинов легкие составляют по весу 27% веса тела, тогда как у наземных млекопитающих только около 12% (Клейненберг, 1956). Количество легочных альвеол почти в три раза превосходит таковое у человека (150 млн. у человека и 437 у морской свиньи). Сильно развита у дельфинов хрящевая ткань, окружающая не только трахею, но и бронхи и бронхиоли и не позволяющая легким спадаться. Такая структура легких в условиях ныряния и повышения даі тения предохраняет животное от кессонной болезни (Scholander, 1940). Вес сердца дельфина несколько меньше, чем у волка того же веса (около 30 кг) (Клейненберг, 1956). Скорость кровотока у ныряющих организмов понижена. Дыхательная функция крови некоторых ныряющих животных отличается несколько повышенной кислородной емкостью.
В табл. 24 приведены некоторые данные сравнения дыхательной функции крови человека, собаки и ныряющих млекопитающих.
Таблица 24
Дыхательная функция крови
(по Irving, 1939)
| Объек г | Кр mb (m.j о2;юо.«.?) | Эритр ЦП ы (.мл Oj/100 л/л) |
| Человек ..................................................... | 20.7 | 45 |
| Собака ...................................................... | 21,8 | — |
| Тюлень....................................................... | 29.3 | 61,3 |
| Морской лев.............................................. | 19,8 | 68 |
| Бобр........................................................... | 17.7 | 40 |
| Ондатра..................................................... | 25,0 | 50 |
| Дельфин .................................................... | 20.5 | 57.7 |
| Кит............................................................. | 14.1 |
Запасы кислорода в теле тюленя к моменту ныряния (вес животного 29 кг)
(по Scholander, 1940)
| Место связывания О2 | Общее количество Оа (лог) |
| Альвеолярный воздух (350 мл с 16 об. % Ог) Кровь (4500 мл с 25 об.% О->).......................................... Мышцы (6000 г с 4,5 об.% О2)........................................... Ткани (кроме мышц)........................................................... | 50 1100 270 100 |
| Весь запас О2 . . . | 1520 |
С уверенностью можно говорить о большой кислородной емкости крови только у некоторых ныряющих форм. У многих она нисколько не выше, чем у наземных организмов. Одно увеличение дыхательной функции крови не может обеспечить возможности ныряния. Содержание миогемоглобина в мышцах типичных ныряющих животных, особенно китов, резко повышено. У тюленя количество его в 7 раз выше, чем у быка (Robinson, 1939). Это же обнаружено и в сердечной мышце (Theorell, 1934). Эти факты были затем подтверждены для китов и тюленей (Scholander, 1940; Коржуев, 1949). Таким образом, огромные массы мышц крупных морских млекопитающих во время ныряния оказываются снабженными кислородом за счет восстановления мышечного гемоглобина. Есть предположение, что у китообразных мышечный аппарат снабжается кислородом во время ныряния именно за счет миогемоглобина.
Регуляция дыхания и кровообращения у ныряющих форм отличается исключительно малой чувствительностью дыхательного центра к накоплению СОг.
Это было установлено еще в прошлом столетии (Bert, 1878; Richet, 1898) и подтверждено многочисленными современными исследователями. Отсутствие реакции на накопление СОг характерно и для таких ныряющих форм, как тюлени, бобры, ондатры, т. е. животных, далеко отстоящих в систематическом ряду друг от друга (Irving, 1939). Напряжение СОг в альвеолярном возду-хе во время ныряния резко возрастает, но это не служит раздражителем для дыхательного центра. Следовательно, кислород, имеющийся в легких, может быть использован в большей мере, чем у не ныряющих организмов. Однако в последнее время прямая нечувствительность дыхательного центра к накоплению углекислоты подвергается сомнению. У уток при децеребрации и трахеотомии (т. е. при выключении влияний с высших отделов центральной нервной системы и назо-фарингеальных рецепторов) наблюдается ярко выраженная реакция при вдыхании воздуха с 6—10% СОг. Следовательно, имеет место постоянное торможение дыхательного центра, возрастающее при раздражении интраторакальных рецепторов (Jukes, Jukes a. Banihfa, 1968).
У ныряющих животных во время ныряния постоянно наблюдается уменьшение частоты пульса. По-видимому, это связано с уменьшением кровоснабжения мускулатуры, которая у всех ныряющих животных (кроме китообразных) во время ныряния расслабляется.
Вопрос об изменениях сердечной деятельности при нырянии на разных глубинах был изучен детально (Harrison, 1960). При небольших погружениях длительностью не более 15 мин у тюленей наблюдалась лишь скоропроходящая брадикардия, сменявшаяся тахикардией. Если период брадикардии продолжался 3 мин, то «восстановительная» тахикардия—до 5—10 мин. Обнаруживается и связь брадикардии с возрастом животного. У старых тюленей эти явления развиваются медленнее, чем у молодых. Более глубокие погружения (от 24 до 91 л*) осуществлялись в специальной камере, наполненной водой, где повышалось давление. При этом у всех животных (как старых, так и молодых) наступает сразу брадикардия (пульс от 4 до 15 в минуту), сменяющаяся через 2—3 мин тахикардией, доходящей до 30 пульсовых ударов в минуту.
Интересно, что в воздушной среде аналогичные повышения давления вызывают лишь очень слабый эффект со стороны сердца. Только при очень больших повышениях давления в воздушной среде (соответствующих глубине погружения 60 м) наблюдались брадикардия и одновременное урежение дыхания.Минутный и систолический объем сердца у тюленей сразу после 10 минутного пребывания под водой оказывается резко повышенным (минутный с 5,5 л до 14,7 л и систолический с 38 мл до 100 мл) (Давыдов и Макарова, 1964). Столь значительное повышение систолического объема сердца свидетельствует об особенностях сердечной мышцы. Расширение сердечной мышцы у человека и у наземных животных не превышает двукратного объема систолы во время работы по сравнению с покоем.
Регуляция щелочно-кислотного равновесия у ныряющих животных отличается, кроме особенностей невозбу- димости дыхательного центра к накоплению СОг, повышенными щелочными резервами крови. Суммарный запас кислорода к моменту ныряния складывается из содержания его в легких, крови и мускулатуре. Кроме того, имеется некоторое количество его в подкожном жире, где может раствориться кислорода в 5 раз больше, нежели в воде (Irving, 1939). Полное использование кислорода из легких благодаря отсутствию реакции на накопление СОг, использование запасов кислорода из миогемоглобина и, наконец, значительные изменения кровообращения с преимущественным снабжением мозга обусловливают способность животного к существованию в условиях дчительного выключения акта дыхания. По-видимому, раздражителем дыхательного центра этих животных служит недостаток кислорода, оказывающий влияние на рецепторы каротидного синуса.
В конечном счете, продолжительность ныряния определяется запасами кислорода в организме и способностью тканей, в том числе и нервной системы, функционировать в условиях резкой гипоксемии. У ныряющих животных содержание кислорода в тканях и крови значительно.
Совершенно особое значение в физиологии ныряющих животных имеют сетевидные сосудистые сплетения, так называемая чудесная сеть. Эти образования встречаются у китообразных в грудной клетке вдоль сонной и позвоночной артерий. По мнению некоторых исследователей, чудесная сеть обеспечивает кровоснабжение мозга во время ныряния. При этом резко сокращается кровообращение в мускулатуре и возрастает в центральной нервной системе. Эти явления, возникающие при нырянии, в последнее время стали характеризоваться как своеобразный «рефлекс ныряния». Последний предполагает трансформацию кровообращения в «сердечно-мозговую
2G6
цепь», направляющую все резервы Ог к сердцу и мозгу, почти полностью приостанавливая кровообращение даже в интенсивно работающих мышцах.
Рефлекс начинается с раздражения рецепторных окончании носа и через бульбарные механизмы резко усиливает реакцию, возбуждаемую через хеморецепторы сосудов. Кожные артериовенозные анастомозы обеспечивают быстрое возвращение крови к сердцу без потерь Ог в тканях. Такой круюоборот дает возможность полностью использовать резервы О2 мозгом и сердцем в период ныряния. При всплытии наблюдается быстрое увеличение минутного объема сердца (до 50 раз против уровня покоя) (Folkow, 1968).
Развитие ныряния в онтогенезе совершенно не изучено. Известно, что молодняк тюленей — бельки, остается длительное время (до 1 месяца) после рождения на льду. Однако наблюдалось нормальное плавание белька гренландского тюленя, непосредственно перед этим извлеченного оперативным путем из матки (В. К- Шепелева). Неизвестно, в какой мере это относится также и к грызунам— бобрам, ондатрам.
Известно также непродолжительное произвольное ныряние у детеныша тюленя, извлеченного из матки (Уждавини, Шепелева, 1966). Время ныряния не превышало 1 мин. Детеныши гренландского тюленя сразу после рождения способны оставаться под водой (при насильственном погружении) лишь самое минимальное время (2—5 мин). В более старших возрастах (стадии «хохлуша» и «серка») возможность погружения возрастает до 10 мин\ отдельные животные оставались неподвижно под водой до 7,5 мин (Давыдов и Макарова, 1964).
Нервные регуляции ныряния следует рассматривать как систему рефлекторных актов, связанных с раздражением определенных рецепторных образований на поверхности кожи. іМожно полагать, что большое значение в этих актах имеет терморецепцня, так как благодаря значительной теплопроводности воды раздражения возникают при каждом погружении. У лягушек, как было показано выше, имеет преимущественное значение специальная хеморсцепция.
Особенности центральной нервной регуляции у ныряющих животных проявляются в описанном виде рас- 267
слабления мускулатуры. Это установлено для утки (Bert, 1878), ондатры, бобра и тюленя (Irving, 1938). Утки при однократном погружении в воду способны выживать в течение 5—15 мин (Richet, 1898). После многократных погружений способность выживать под водой возрастает до 12—17 мин. Децеребрация утки (Huxly, 1913) не устраняет типичной задержки дыхания при погружении головы и клюва в воду, как не устраняет общего для ныряющих форм замедления пульса. Экстирпация лабиринтов снижает этот эффект, но не устраняет его. По-видимому, он осуществляется в этом случае с рецепторов шейных мышц.
Есть основания полагать, что у ныряющих и наземных млекопитающих и у птиц нет принципиальной разницы в реакциях при нырянии. Эти различия носят количественный характер и зависят от особенностей как рецепторов, так и центров, регулирующих дыхание. Ваго- томированный утенок становится таким же чувствительным к асфиксии, как цыпленок (Richet, 1899). Ритм дыхания у ныряющих организмов отличается прерывистостью. Тип периодического дыхания (Чейн-Стокса) характерен в норме для таких организмов, как бегемот, но не встречается у тюленей.
Изменения мышечного тонуса во время ныряния были установлены электромиографическими исследованиями, проведенными над молодняком гренландского тюленя (А. Ф. Давыдов и А. Р. Макарова). Погружение в воду (без погружения головы) вызывает очень небольшое снижение электрической активности скелетной мускулатуры. Погружение тела с головой вызывает резкое ограничение электрической активности, сопровождающееся, как уже указывалось выше, урежением пульса. Так как понижение тонуса возникает сразу же после погружения, нет оснований думать, что это следствие накопления СОг, как это имело место в исследованиях И. С. Репина (1959а), и недостатка кислорода, как в опытах К- П. Иванова (19596). Можно думать о существовании специального механизма, снижающего мышечный тонус у ныряющих животных (тюленей), обусловливающий сниженный кислородный запрос в период ныряния (рис. 61).
Исследование потребления кислорода во время ныряния (А. Ф. Давыдов и А. Р. Макарова) обнаружило повышение газообмена только в первую минуту после 10—15-минутного погружения в воду. Затем наступает довольно длительное (в пределах до 10—25 мин) снижение потребления кислорода ниже исходного уровня. В результате этого снижения суммарное потребление Ог за период восстановления может быть более низким, чем
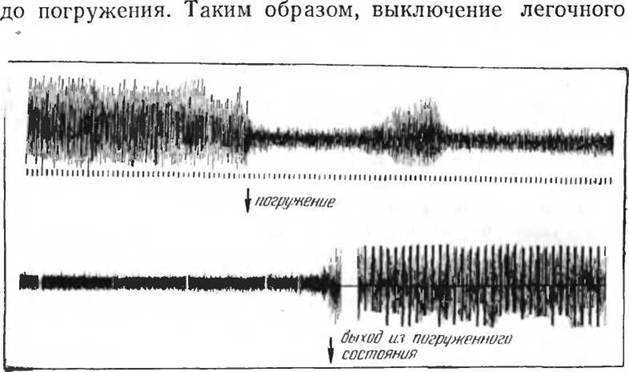
Рис. 61. Эчектромнограммы скелетных мышц тюленей при погружении в воду (по Давыдову и Макаровой), 1967)
дыхания, снижение мышечного тонуса и урежение пульса приводит к значительному снижению кислородного запроса у ныряющих тюленей. Правда, эти опыты поставлены в условиях насильственного погружения, в покое. Очень интересен вопрос, как развиваются эти явления при интенсивной мышечной работе — плавании в погоне за рыбой. Однако ответа на него пока нет.
Итак, основными физиологическими механизмами, определяющими продолжительность ныряния, являются, с одной стороны, ограничение кислородного запроса организма благодаря расслаблению скелетной мускулатуры, а с другой — наличие некоторых запасов кислорода в теле (в крови, мышцах и воздухоносных путях). Важным механизмом является нечувствительность дыхательного центра ныряющих животных к накоплению угольной кислоты в крови, благодаря чему возможна более полная утилизация кислорода крови.
Еще по теме ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ НЫРЯЮЩИХ ОРГАНИЗМОВ:
- КЛЕТОЧНЫЙ И ТКАНЕВОЙ УРОВНИ АДАПТАЦИЙ У ГОМОЙОТЕРМНИХ ОРГАНИЗМОВ
- АДАПТАЦИИ РАВНИННЫХ ОРГАНИЗМОВ В ГОРАХ И В ЭКСПЕРИМЕНТЕ
- ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ НЫРЯЮЩИХ ОРГАНИЗМОВ
- ПЕРЕДВИЖЕНИЕ НАЗЕМНЫХ ОРГАНИЗЛЮВ
- ОГЛАВЛЕНИЕ