АДАПТАЦИИ РЕПТИЛИЙ К ПУСТЫНЕ
Широко распространенные в аридной зоне рептилии представляют весьма интересный пример адаптаций организма, включающих как элементы подчинения факторам внешней среды (conforming organisms), так и элементы регуляции (regulating organisms), предохраняющие животное от чрезмерных сдвигов во внутренней среде.
В пустыне встречаются представители всех отрядов — змей, ящериц и черепах, питающихся как растительной, так и в большинстве животной пищей.У многих представителей рептилий, главным образом ящериц, обнаружены явления видовых температурных адаптаций, выражающихся в различных температурах, при которых наступает холодовое окоченение. Для ящериц характерно то, что активность наступает лишь при определенной температуре тела, которая поддерживается на постоянном уровне в течение всего времени, пока животное активно (колебания 0,56 — до 0,005°С). Такое своеобразное поддержание температуры тела достигается реакциями поведения, четко выраженной предпочитаемой температурой. Температура тела пустынных 280
рептилий в состоянии активности в значительной мере поддерживается и солнечным облучением. При этом некоторые виды способны изменять свою окраску и тем самым регулировать поглощение и отражение лучей различной части спектра.
Поскольку поверхность почвы в пустыне может в дневные часы нагреваться свыше 70° С, рептилии в это время суток укрываются в норах или на ветках растений (Кашкаров и Коровин, 1936). Многие виды пустынных ящериц—агамы, кругоголовки зарываются в песок, глубокие слои которого имеют более низкую температуру, нежели поверхность, и таким образом изолируются от прямого воздействия солнечных лучей.
Однако реакции поведения дополняются у пустынных рептилий мощным механизмом поддержания температуры путем испарения воды с поверхности верхних дыхательных путей — полипноэ. Испарение с поверхности тела, как и кожные железы у рептилий отсутствуют.
Термическое полипноэ лучше всего изучено у ящериц. При этом обнаружено, что между интенсивностью полипноэ, средней температурой активности и летальной температурой существуют определенные прямые и обратные зависимости (табл. 26).
Таблица 26
Реакция на нагревание у трех видов ящериц
(по Dawson a. Templeton, 1964)
| Виды | Температура | |||
| Средняя активности | Летальная | Наступления полипноэ | Возрастания частоты пульса на 1% | |
| Сцинк (Eumeces obso- letus) | 33 | 43 | Отсутст- | 2 |
| Ящерица (Crotaphytus collaris) | 38 | 46,5 | вует 42,5— 43,0 | 4.5 |
| Игуана (Dipsosaurus | 39-42 | |||
| dorsalis).................................... | 48—50 | 43 | 7 | |
Очень важное значение в поддержании постоянной температуры тела имеет так называемый париетальный глаз. При удалении его температура тела ряда видов ящериц повышалась при нагревании как в природной обстановке, так и в лаборатории (Stehlin a. Eakiri, 1958). Такие данные были получены В. Н. Долининым (1968) при заклеивании париетального глаза пластинкой станиоля. Эти факты указывают на рефлекторный характер поддержания температуры тела у пустынных ящериц, но изучены совершенно недостаточно.
Изменение окраски ящериц под влиянием солнечного облучения является одним из важных механизмов поддержания температуры тела. Отражаются от поверхности тела преимущественно инфракрасные (тепловые) лучи.
У европейских видов отражение составляет около 20% магниевого (MgO) стандарта. Однако исследование ряда видов с различным характером обитания указывает на исключительную роль отражения лучей у пустынных видов. Так, отражающая способность для лучей от 320 до 1100ммк составляет для тропической ящерицы- игуаны (Iguana iguana) 6,2%, а для пустынной (Phery- nosotna plathyshinos)—35%. Резкое уменьшение поглощения солнечного тепла наступает у ящериц благодаря побелению поверхности кожи спины, наступающему одновременно с повышением температуры тела. У некоторых видов наступает и побеление кожи живота, что, вероятно, предохраняет животное от отраженного излучения поверхности субстрата.Тканевые адаптации у рептилий в пустыне выражаются в большей термоустойчивости мышечных белков, активности адезинтрифосфатазы (Ушаков, 1963; Schmidt-Nielsen a. Dawson, 1964).
Повышение температуры тела приводит у рептилий к возрастанию обмена веществ, учащению ритма сердечных сокращений. Реакция на температуру среды зависит от дневного и ночного образа жизни животного. Так, летальная температура у дневных видов приходится на 49—50° С, а у ночных — 43—44° С (Saint-Girons et Saint-Girons, 1956). Как правило, температурный предел устойчивости к нагреванию коррелирует с уровнем температуры тела во время активности животных. У пустынных видов ящериц рода Sceleropus температура активности 35° С, а летальная 43° С. У воротниковой яще-
рицы (Crothaphytus collaris) соответствующие температуры равны 38—39° С и 46—47° С. Пустынная игуана активна при температуре тела около 42° С, а ее летальная температура близка к 50° С (Cowles a. Bogert. 1944) (рис. 64).
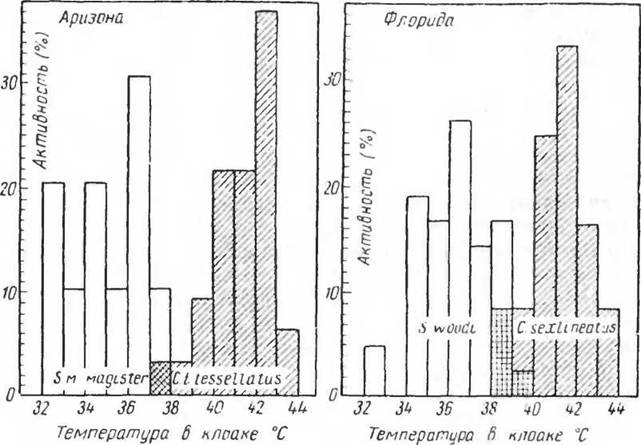
Рис. 64. Температура тела во время активности у ящериц Chemidophorus и Sceloporus magister из Аризоны и Sceloporus inagister из Флориды
(по Bogert, 1949)
Летальные температуры у рептилий обусловлены нарушениями деятельности нервной системы и аноксией в результате нарушения связывания гемоглобином кислорода при высокой температуре среды (Dill, 1938).
Весьма важно для понимания адаптаций рептилий в пустыне изучение потерь воды путем испарения. Как общее правило, потеря воды рептилиями при температуре тела, свойственной гомойотермним организмам (млекопитающим и птицам), составляет от 1/5 до 1/7. Примерно около 1/7 составляет и уровень обмена при высокой температуре тела (Dawson a. Bartholomew, 1958). Однако для пустынных рептилий соотношения несколько иные. Воротниковая ящерица испаряет в 5 раз меньше воды по сравнению с белой мышью, но только в 2 раза меньше по сравнению с кенгуровой крысой (Dipodomis merriami} (Schmidt-Nielsen, 1950).
Пребывание в норах — особенность экономии воды рептилиями. Более низкая температура среды, а следовательно, и более низкий обмен, более высокая относительная влажность, а следовательно, и более низкая испаряющая сила воздуха создают здесь для рептилий наиболее благоприятные микроклиматические условия.
Однако низкий уровень обмена (теплопродукции) создает благоприятные условия для существования рептилий и вне норы, так как в состоянии активности при одинаковой с пустынными млекопитающими температуре тела животному угрожает поступление тепла извне и наполовину меньшее образование тепла в теле. Этим можно объяснить богатую и разнообразную фауну рептилий в пустынях Старого и Нового Света.
Очень низки потери воды испарением через кожные покровы и при дыхании у черепах пустыни. У Gopherus agassizii потери воды оказались намного меньшими, чем у Terrapene Carolina, обитающего во влажной среде (Schmidt-Nielsen a. Bentley, 1966).
Особенно ярко эти зависимости проявляются при сравнении небольшого каймана (Caiman sclerops) с пустынными ящерицами (Iguana iguana и Sauromalus obe- sus). У ящериц испарение воды кожей составляло лишь 5% от этой величины у каймана (Bentley a. Schmidt- Nielsen, 1966). Данные такого рода совершенно изменяют укоренившееся представление о малой проницаемости кожи рептилий. Сравнительно высокие потери воды испарением, однако, хорошо компенсируются малыми потерями воды с мочой и калом. Моча рептилий содержит мочевую кислоту как основной конечный продукт азотистого обмена. Для пустынных рептилий этот тип азотистого обмена является весьма благоприятным, так как при этом теряется минимальное количество воды. Это достигается секрецией гиперосмотической — по сравнению с плазмой — мочи.
Многие рептилии пустыни к тому же обладают удивительной .способностью хорошо переносить увеличение концентрации хлоридов крови. Высокая концентрация летом наблюдается и в полевых условиях. Регуляция водного обмена у пустынных рептилий сходна с той, что имеет место у млекопитающих.
Еще по теме АДАПТАЦИИ РЕПТИЛИЙ К ПУСТЫНЕ:
- Ассоциативные временные связи у рептилий.
- Жаркий климат пустынь
- Особенности пищеварения и водного обмена пустынных млекопитающих
- Высокогорье и горно-пустынная местность
- 27. Профессиональная адаптация. Причины трудностей профессиональной адаптации
- Психофизиологическая адаптация
- Адаптация и компенсация.
- 7.Уровни адаптации
- 35. Адаптация глаза к изменению освещения, ее нарушения
- Психологические особенности адаптации осужденного