5. ДИНАМИКА ЧИСЛЕННОСТИ ПОВДИКД В СОСНОВЬК КУЛЬТУРАХ СТЕПНОЙ зоны и ФАКТОРЫ'ЕЕ ОПРЕДЕЛЯЮЩИЕ
Как уде говорилось вше, сосновые культуры Каменского лесхоза, где проходили исследования, но своей лесоводстве иной характеристике, условиям местопроизрастания и экологической обстановке типичны для культур степной зоны» В нашей страна наибольшая частота вспышек массового размножения и наибольшая вредо - косность обыкновенного соснового пилильщика Проявляются В СОСНОВЫХ культурах степной и лесостепной зон европейской части СССР» Поэтому выявление закономерностей динамики численности пилильщика для этих условий представляет особенно ванную для лесного хозяйства задачу.
Последняя вспышка массового размножения обыкновенною соснового пилильщика в Каменском лесхозе наблюдалась в 1980 г. По данным детального картирования поврежденности сосновьк культур Уляшкинского массива можно проследить подъём и спад численности пилильщика и перемещение его очагов в 1980-1988 гг., то есть в год вспышки (1930 г.) и пославспышечные годы (Рис. 21, 22 ).
Как видно из Рис» 21, в 1980 г. высокая численность пилильщика, сопровождающаяся сильной и средней степенью повреж - де ния сосновых культур, наблюдалась практически по всему массиву на площади около 3 тыс.га. В наибольшей степени повреждались пилильщиком культуры в южной части лесничества (кв. 51-72).
в 1981 г. культуры сосны вновь подверглись значительному объеданию, но центр очагов переместился в северо-восточную часть (кв« 89, 40). в сильной и средней степени повреждались культуры на площади около 200 га. В 1980 г. в этих участках имела место средняя степень повреждения хвои. На остальной территории повреждение хвои было слабым или его вообще не было.
IIS
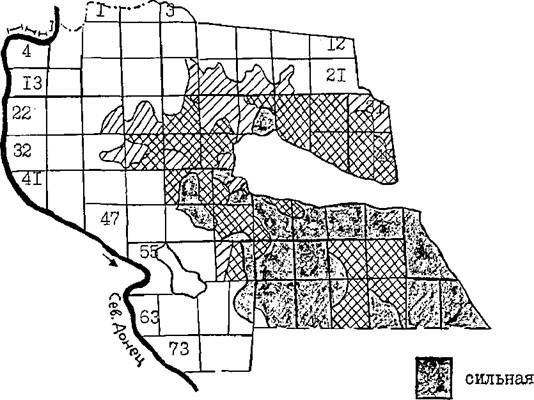
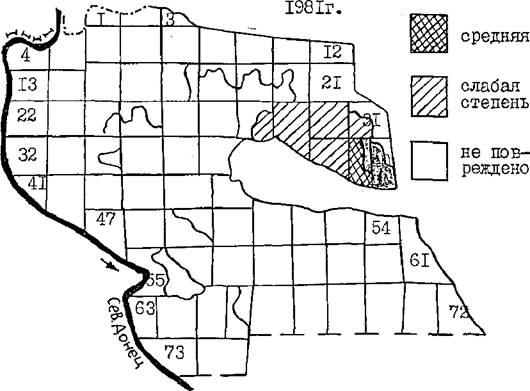
Рис. 21 . Повревденность сосновых культур лесничества пилильщиком в 1980 и 1981 гг.
lift
1982г.
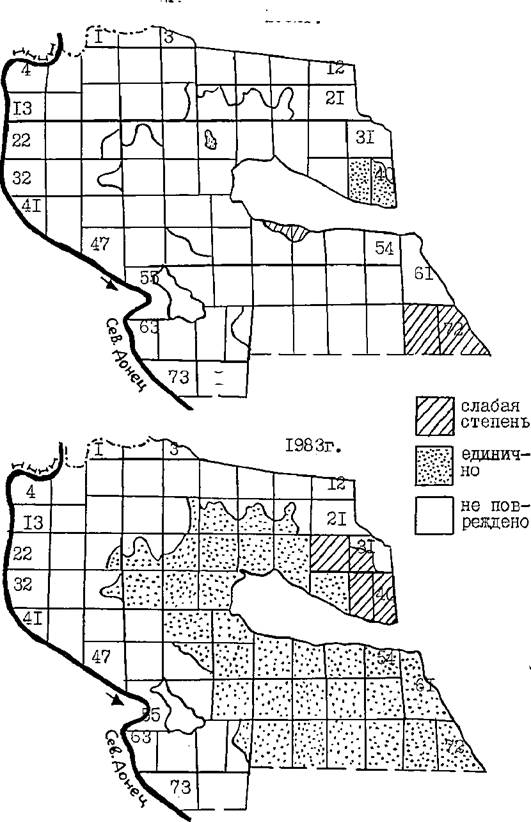
Рис. 22 . Поврежденность сосновых культур лесничества пилильщиком в 1982 и 1983 гго
В 1982 наблюдалась наименьшая за годы исследований численность активной части популяции пилильщика. В кварталах 39 и 40, где выход популяции пилильщика из диапаузы был наибольшим, повреждение хвои было единичным из-за высокой смертности ЯИЦ. Очень незначительное повреждение хвои отмечалось в юго-восточной части лесничества (кв. 71,72), где в 1981 г. почти вся популяция пилильщика находилась в диапаузе.
В 1983 г. по всей территории лесничества отмечались лишь отдельные сосны с единичными колониями личинок, а в кварталах 39 и 40, где постоянно присутствует запас живьк коконов пилильщика, а тагасе в кварталах 30 и ЗХ наблюдалось слабое повреждение хвои.
При сопоставлении данных рисунков 21 и 22 можно видеть,что в постоянных местообитаниях пилильщика имеет место ежегодное перемещение активной части популяции, наблюдаются как бы небольшие волны подъёма численности в тех или иных участках даже в межвспышечные годы. Обращает на себя внимание также наличие участков с более или менее постоянной жизнедеятельностью активной части популяции пилильщика. Например, насаждения в кварталах 39 и 40 (участок № I). По овоей экологической обстановке и лесо- водственной характеристике (возраст - 20 лет, бонитет П, тип условий местопроизрастания - Ь>, посадка полосами шириной 10 м, межполосное пространство 20 м, подрост редкий, подлесок отсутствует) этот участок культур почти не отличается от других участков (см. стр. 22 ), лишь, может быть, относительно большей удаленностью от поймы реки Северский Донец.
Данные о степени повреждения культур в 1931-І933 гг. интересно сопоставить с данными об изменении зкологичесной плотности активной части популяции пилильщика в эти годы (табл. 33).
Таблица 33
Плотность активной части популяции пилильщика в сосновых культурах Уляшкинского лесничества в 1931 - Х933 гг.
| № участка квартал | Годы | Экологическая плотность особей, шт/100 г хвои | 1------------------- ! Степень ! поврекде- ! ния 1 | ||
| яиц | • I ♦ I | личинок 1-го ВОЗр. | |||
| 1931 | 66 £5 | 36,79 | средняя | ||
| 39 | 1932 | 29,97 | 0,62 | единичн* | |
| 1983 | 43,8$ | 33,48 | слабая | ||
| 2 | 1931 | 14,29 | 2,50 | слабая | |
| 30 | |||||
| 1932 | 12,04 | 4,60 | слабая | ||
| 71 | 1933 | 7,92 | 2,97 | слабая | |
Как видно из табл* 33, при средней степени повреждения
(участок Я I, 1931 г.) экологическая плотность личинок в 3 и более раза превышала их наибольшую плотность в участках со слабым повреждением. Следует отметить также, что при внешне одинаковой (слабой) степени повреждения экологическая плотность личинок (вредящей фазы) сильно варьировала (от 2,5 до 13,48 шт/100г хвои), изменяясь более, чем в 5 раз.
Эта непропорциональность начальной экологической плотности пилильщика степени повреждения хвои объясняется разным набором и различным влиянием факторов смертности пилильщика, действующих в тех или иных участках и нивелирующих конечную плотность личинок старших возрастов и связанную с ней степень объедания хвои. Наибольшая экологическая плотность пилильщика в меквспышечные годы, как видно из табл. 33, достигала около семи десятков яиц и около четырёх десятков личинок первого возраста на 100 г хвои (то есть примерно на ветвь с диаметром около 1,2 см у её основания), что сопровождалось максимальной в этот период средней степенью повреждения хвои.
На примере тех не четырех участков стационарна наблюдений прослежено изменение экологической плотности популяции пилильщика с осени 1930 г по осень 1983 г (табл. 3$). Поколения пилильщика обозначены следующим образом: генерация 1980 г - г, 80, генерация 1981 г - г, 81 и т.д. £ расчётах использованы данные 680 пробных площадок размером 0,125 м^ и 756 модельных ветвей.
Таблица 34
Изменение экологической плотности пилильщика в сосновых культурах Уляшкинского лесничества в 1931 - 1983 гг.
| № j наб-; лю- ; де- ; ний ; | Даты наблюде ний . І | Фаза Развития, • генерация | {Экологическая плотность ;популяции по участкам ; Сшт/ІОО г хвои4) | |||
| Iі г | 2 ! | | S ; * t - | 4 | |||
| І і | 2 І 3 | 1 4 ! | 5 І | 6 і | 7 | |
| X | X, 1980 г | коконы г, 80 | 24,62 | 7,IX 22,29 | - | |
| 2 | ІУ, 1981 Г | коконы г, 80 | 14,23 | 3,20 XI,42 | - | |
| 3 | конец УП, | |||||
| 1981 г . | коконы г, 80 | 4,92 | 0,53 | 1,86 | ||
| 4 | нач. УШ, | коконы г, 80 | 1,53 | 0,03 | 0 ,65 | м |
| 1981 Г | имаго г, 80 | 3,34 | 0,50 | 0,71 | - | |
| 5 | нач» УШ, | коконы г, 80 | 1,58 | 0,03 | - | |
| Х93І г | яйца г, 81 | 66,25 14,29 | - | - | ||
| 6 | серед. УШ, | коконы г, 80 | 1,58 | 0 ,03 | — | |
| І98Х г . | личинки 1-го возр. | |||||
| Г,81 | 36,79 | 2,50 | — | - | ||
| 7 | нач. X, | - | ||||
| 1931 Г | коконы г, 80, ЗХ | 29,35 | 0,68 | — | ||
| 3 | ІУ, 1932 г | коконы г, 80, 81 | 22,85 | - | - | - |
| 9 | нач. УШ, | |||||
| 1932 г | коконы г, ЗО, ЗХ | 13,18 | - | - | - | |
ИВ
продолжение таблицы 34
| I | I 2 | 1 | 3 1 | і 4 1 | 5 1 | і 6 | 1 7 |
| 10 | серед. УШ, | коконы | г, 80, 81 | 7,09 | |||
| 1932 г | имаго | г, 80, 81 | 6,09 | - | - | - | |
| II | серед* УШ, | коконы | г, 80, 81 | 7,09 | - | ||
| 1932 г . | яйца | г, 82 •' | 29,97 | - | «и | 12,04 | |
| 12 | нач. JX, | коконы | г, 80, 81 | 7,09 | HI | на | |
| 1982 г. | личинки | 1-го воэр. | |||||
| г, 82 | 0,62 | - | *н | 4,60 | |||
| 13 | серед. X, | коконы | г, 80, 81, | - | - | ||
| 1982 г . | 82 | 7,25 | - | - | 2,91 | ||
| 14 | ІУ, 1983 г | коконы | г, 80, 81, | ||||
| - | 02 | 5,18 | — | - | 1,41 | ||
| 15 | конец УП, | коконы | г, 80, 81, | - | |||
| 1983 г | 82 | 3,97 | — | - | 0,44 | ||
| 16 | нач* УШ, | коконы | г, 80, 81, | ||||
| 1983 г | 82 | 1,56 | - | - | 0,14 | ||
| имаго | г, 80, 81, | ||||||
| 82 | 2,41 | — | •»* | 0,30 | |||
| 17 | нач* УШ, | коконы | Г, 80, 81, | ||||
| 1933 г | 82 / | 1,56 | - | - | 0,14 | ||
| яйца | Г, 83 | 48,84 | - | - | 7,92 | ||
| 18 | серед* УШ, | коконы | г, 80,81, | ||||
| 1983 г | 82 | 1,56 | - | - | 0,14 | ||
| личинки | 1-го возр. | -■ | |||||
| г, 83 | 13,48 | - | - | 2,97 | |||
| 19 | нач. X, | ||||||
| 1933 г | коконы | г, 80,81, | - | ||||
| 82,83 | 8,60 | «и | нг | 1,30 |
На основании данных табл. 34»вычислены коэффициенты баланса популяции обыкновенного соснового пилильщика по периодам его развития (табл. 35). Коэффициенты определялись как частное от деления экологической плотности популяции в конце этапа на плотность популяции в начале этапа. Данные для определения коэффициентов баланса популяции за период лёта взяты из таблиц вживания (см* табл. 16, глава 3)*
IB
Коэффициенты баланса популяции ( К ) обыкновенного соснового пилильщика га периоды
Таблица 35
| пп | г ----------- ' і-------------------------- • • 1 1 Лоои w | Г К Г і № і іуч- ; їства і- • * ! ! | Коэффициент баланса популяции ( К ) | |||
| j периоды « I а і | стадии | |||||
| 1930 ї 1931 • | 1932 І 1983 » | |||||
| I | І 2 | 3.... | І 4 ! | 5 ! 6 | 7 ! 8 | |
| I | Зимовка и BQ- | эонимфа | І | 0,20 0,45 0,55 | ||
| сенне-летняя | В КОКО- | 2 | 0.07 | |||
| диапауза | не | |||||
| 3 | 0,06 | а* | ||||
| 4 | 0,15 | |||||
| 2 | Лёт | имаго | І | 0,26 | 0,07 0,26 | |
| 2 | 0,36 | - | ||||
| 3 | 0 | - | ||||
| 4 | - | 0,30 | ||||
| 3 | Яйцекладка | ЯЙЦО | І | - 19,84 | 4,92 20,27 | |
| 2 | - 28,58 | - | ||||
| 3 | ** «а | - | ||||
| 4 | - | - 26,40 | ||||
| * | Развитие яиц. | яйцо, | І | 0,42 | 0,01 0,14 | |
| питание личи- | личинка | 2 | 0,04 | а* — | ||
| нок | 4 | |||||
| 3 | — — | “ — | ||||
| 4 | - | 0,24 0,15 | ||||
| 5 | Год (от осени | яйцо ,ли- | І | 1,19 0,25 1,19 | ||
| до осени) | чинка, | 2 | 0 *10 “ ** | |||
| эонимфа, | ~~ | |||||
| пронимфа | 3 | ■ | ||||
| куколка, | 4 | 0,45 | ||||
имаго
Данные табл. 35 целесообразно дополнить обобщенными результатами изучения роли факторов смертности (табл. 36). Сопоставление данных табл. 35 и 36 показывает какие факторы в наибольшей степени влияли на баланс популяции обыкновенного соснового пи - лильщика в меквспышечные годы (табл. 36).
Данные табл. 35 иллюстрирует рис. 23 , на котором в количественном и временном условных масштабах показано изменение плотности популяции пилильщика в годы исследований.
Как показывает табл. 35, в меквспышечные годы популяция значительно убывала за период зимовки и весенне-летней диапаузы зонный в коконах в подстилке. Коэффициент баланса популяции за этот период показывает, что в различных участках и в разные годы убыль популяции значительно отличалась. В участке № I (кв. 39), где наблюдения были наиболее длительными, убыль в этот период была наименьшей (К 0,2) и коэффициенты баланса популяции
увеличивались от года к году по мере удаления от конца вспышки (от К = 0,20 до К = 0,55). Как было показано в главе 3 и табл. 36, убыль популяции в этот период происходила, в основном, под влиянием паразитов (в среднем от них гибло 45,9% коконов).
На втором месте по значению стояжболезни (22,2%), затем - хищники (6,6%).
Анализ данных показал, что убыль популяции в период диапаузы эонимф в подстилке в какой-то мере прямо пропорциональна степени повреждения сосны в предыдущий год. Так, смертность пилильщика на фазе кокона в участке № I (кв. 39) (при среднем повреждении в 1980 г.) была намного меньше, чем в участка Л 3 (кв. 51), где в 1980 г. было сильное повреждение хвои. Пораженность коконов болезнями в первом случае равнялась 18,20%, а во втором - 40,68%, Гибель коконов в участке 1S 3 под действием комплекса факторов в 1981 г. была столь велика, что популяция практи-
Таблица 86
Роль факторов смертности обыкновенного соснового пилильщика на разных фазах и в разные периоды его развития в I98I-I983 гг
| ......... ............... • Фазы и стадии ! развития пилиль- ! щика ! * I • 1 | Повтор ность наблюде ний | j Смертность от отдельных групп факторов, | |||
| J г ! Паразиты ! • 4 ! ! ! ! і і • ♦ | ХИЩНИКИ И { "Кпттяяни случ.факто-і Волезни ры во время і ' перемещений; • t * | І Высыха- І НИ6Г і яиц t • 1 • | (Неизвести., {ЭНД0Г6Н. {факторы ІРезистентн, {дерева. • | ||
| Яйцо | 6 | 52.31 | 0 0 | 16.34 | L.64 |
| 35,86-77,11 | 4,12-25 | ,76 ІіОХ-2,44 | |||
| Личинки 1-6 возраста, | 6 | 0 | 27.60 8,10 | 0 | 17.48 |
| свободные эонимфы | 14,52-45,16 3,78-12 | •0 | 6,22-28,40 | ||
| Эонимфы, пронимфы, | 6 | .45,95 | 6,56 22.16 | 0 | 0.60 |
| куколки в коконах | 20,42-72,85 | 4,10-10,59 11,25-40 | ,68 | 0,08-1,93 | |
| Имаго | 6 | 0 | 79.XI 0 | 0 | 0 |
181
64,0-100,0
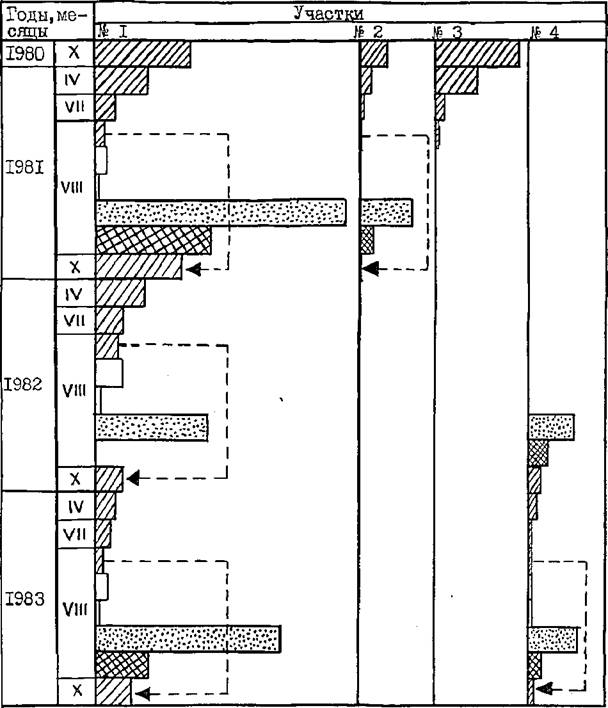
Рис. 23
Изменение экологической плотности пилильщика за 1980-1983 гг. в 4-х участках Уляшкинского лесничества Каменского лесхоза (в I см 10 особ./ЮОг хвои)
Фазы и периоды развития

лйцо личинка
кокон
о
имаго
чески вымерла и в последующие годы повреждений сосны здесь не отмечалось.
Резкое увеличение плотности популяции, как и следовало ожидать, происходило после яйцекладки летней генерации пилильщика за счет высокой плодовитости самок, но его частично уравновешивала относительно высокая смертность имаго ( К СО,36). Так, например, благодаря последнему обстоятельству, в участке № I в 1982 г. плотность популяции возросла только в 4,9 раза, тогда как средняя плодовитость самок в этот год равнялась III ,0^5,0 яиц
Убыль популяции на фазе яйца и во время питания личинок в межвспышечные годы была достаточно высока во всех случаях. Наиболее низкие коэффициенты баланса популяции ( К = 0,01 и К = 0,04) указывают на высокую смертность особей, которая была наибольшей в этих случаях на фазе яйца, в основном, под влиянием яйцеедов (участки JS I и № 2).
Коэффициенты баланса популяции пилильщика, характеризующие изменение его плотности в целом за один год (от осени одного года до осени следующего), свидетельствуют о том, что его популяция находилась в состоянии депрессии: незначительные увеличения плотности популяции в некоторых участках в одни годы (К=1эр9) чередовались со спадами в другие (В>0,1; 0,25; 0,45). Рост плотности популяции пилильщика происходил в таких участках, где наблюдался активный лёт летнего поколения и осенью увеличивался запас зимующих коконов за счёт закоконировавшихся эонимф. Таким образом, летние поколения пилильщика в межвспышечные годы восполняют постоянную убыль длительно диапауз ирующих эонимф, поддерживая определенный уровень численности вида в его местообитаниях. В связи с этим,случаи полного вымирания популяции пилильщика возможны только в редких случаях (см.табл.35),хотя убыль популяции на отдельных этапах жизненного цикла бывает очень велика
( К = 0,01; 0)04$ 0)06 и т.п.).
Постоянный запас диапаузирующих эонимф пилильщика имеет очень большое биологическое значение в обеспечении сохранности вида в его местообитаниях- Так, доля диапаузирующих эонимф пилильщика при летних учетах в различных участках по годам колебалась от 6 до 54% и составляла в среднем 35%. Плотность их на I м^ подстилки варьировала от 0,2 до 45,5 шт/м^ и в среднем равнялась 11,3 шт/м2- При этом плотность недиапаузирующих особей в межвспышечные годы составляла в среднем 14,3 шт/м^, то есть незначительно превышала среднюю плотность диапаузирующих.
При анализе и прогнозировании динамики численности очагов хвоегрызущих насекомых большое значение придается, так называемым, количественным и качественным показателям состояния популяций. К числу важнейших из них относят абсолютную и экологичесвдгго плотность популяции, соотношение полов, размеры и вес особей, плодовитость, пораженность особей болезнями и энтомофагами, коэффициент размножения, поврежденность зелёной массы деревьев вредителем и проч. (Ильинский и др., 1965, Воронцов, 1932 и др.)
В табл. 37 сведаны некоторые показатели состояния популяции обыкновенного соснового пилильщика в межвспышечный период.
Таблица 37
Некоторые популяционные показатели обыкновенного соснового пилильщика в годы наблюдений
| № і пп і * | Показатели | 3—5 j--------- j участков і I93O | Годы Ї 1931 ! . | 1932 ' 1983 | ||
| і ! | 2 | 1 з! | 4 | І 5 і | 6 | І 7 |
| і | Средний процент | I | 50 | 50 | 5 | 20 |
| повреждения куль- | 2 | 40 | хо | 0 | 10 | |
| тур сосны | 3 | 30 | 0 | 0 | 0 | |
| 4 | 70 | 0 | 10 | 5 | ||
| в среднем по всему лесничеству | 60 | 5 | 2 | 6 | ||
| I | 1 2 | I | 3 | ! 4 ! | 5 | І 6 ! 7 |
| 2 | Абсолютная | J "' ' ‘ ‘ ілетний' | І | не учит. | 31,6 | 84,6 25,5 |
| плотность | {запас • | 2 | —U. | 8,7 | не учитывал. | |
| коконов в | 1 t | 3 | -П- | 3.4 | 0 0 | |
| подстилке, | ( | |||||
| шт*/м^ | 1 « 1 | 4 | не учитывалось 2,8 | |||
| j осенний {запас 4 1 • 1 t • 1 ♦ | І | 158,0 | 188,3 | 46,5 55,2 | ||
| 2 | 45,6 | 4,4 | не учитывал. | |||
| 3 | 143,0 | 4,2 | 0 0 | |||
| 4 | не учитывалось | И,7 8,3 | ||||
| 3 | Доля диапаузирующих | І | не учит. | 32,1 | 53,8 39.3 | |
| эонимф, %% | 2 | -П- | 5,7 | не учитывал. | ||
| 3 | —П_ | 47,8 | - ~ | |||
| 4 | -її- | не учитывалось 31,8 | ||||
| 4 | Соотношение | подов | І | 1:2,3 | 1:1,9 | ІЙ,0 1:1,8 |
| ( o’ 5 ? | ) | 2 | 1:1,9 | 1:1,5 | не учитывал. | |
| 3 | 1:0,7 | - | - | |||
| 4 | не учитывалось | 1:2,1 1:1,5 | ||||
| 5 | Экологичен- | Г ’ 1 ! яйцо 1 | І | не учит* | 66,25 | 29,97 48,84 |
| кая плот- - ность попу- | 1 І I | 2 | — Н— | 14,29 | не учитывал. | |
| і | 3 | _пм | 0 | 0 0 | ||
| ляции, | і | - . - | ||||
| шт ./100 г | і t | 4 | не учиш | .12,04 7,92 | ||
| хвои | 1 і | |||||
| І І личинка і | І | не учит. | 36,79 | 0,62 ІЗ ,48 | ||
| і { | 2 | —Я— | 2,50 | не учитывал. | ||
| і і | 3 | -Я- | 0 | 0 0 | ||
| і і | 4 | не учит | . 4,60 2,97 | |||
I!
Плодовитость, шт .яиц
для
всех
участ
ков
129,9± III,0^ 132,0± не учит. 4,9' 5,0' 12,7’
Количество пораженных ентомофагами и болезнями, %%
------------------------ Р
3 Выживаемости %%
38,08 77,11 66,09
57,59 на учитывал.
яиц I не учит.
2
3 ’
4 на учит. 39,12 35,86
коко
нов
яиц
1
2
3
4
79,89 54,10 43,31 92,40 не учитывалось 93,80
не учитывалось 84,54
не учит
55.5 2,1 27,6
17.5 не учитывал.
| ! ! | ї . | 3 | - - _ | |
| 4 | не учит. 38,2 37,4 | ||
| і личи- | І | не учит. | 75,5 25,8 52,2 |
| ! нок 1 | 2 | 26,0 не учитывал. | |
| І і | 3 | — - г | |
| і і | 4 | не учит. 63,3 37,4 | |
| I і коко- І нов ! | 1 2 | 20,0 7,5 | 44,9 54,8 не учитывалось |
| і і | 3 | 6,1 | г |
| і і | 4 | на учитывалось 15,1 | |
имаго
1
2
3
4
на учит
26,0 7,6 25,7
36,0 не учитывал.
— п_
не учитывалось 30,0
| X 1 2 | 1 | 1 3 1 4 I | 5 ! | 6 1 7 |
| 9 Коэффициент | 1 !по ко- 1 - ;нонам, | I 1,19 | 0,25 | 1,19 |
| баланса попу | 2 0,10 | |||
| ляции | !ош осе- | О | ||
| 1НИ до ;осени | 3 - | - | ||
| 4 | - | 0,45 | ||
| Как видно | из данных | табл. 37, межвспышечные | годы характери- | |
зовались незначительной степенью поврежденности хвои, в среднем не превышающей 6%. Плотность коконов в подстилке при этом достигала от нескольких штук до почти 200 шт, на I м^ подстилки, доля диапаузирующих особей колебалась в отдельных участках по годам.приблизительно от в до 54%, а экологическая плотность яиц и личинок первого возраста варьировала от 0 до 66 шт/100 г хвои.
Соотношение полов пилильщика, определенное по коконам, в различных участках по годам наблюдений такие сильно варьировало. Как уже говорилось ранее (см. табл. 7, раздел 2.3), оно зависело от степени повреждения хвои в участках в предыдущий год. Так, например, в участке № 3, где в 1930 г. было сильное повреждение, уже осенью 1930 г. преобладали самцы, тогда как во всех других участках, повреждавшихся в средней и слабой степени, соотношение полов всегда было в пользу самок. Регуляция соотношения полов может происходить и в период питания вследствие нехватки корма для личинок 6-го возраста (Рывкин, 1963), и в период лёта имаго. Как показали наблюдения западногерманских ученых, при высоких плотностях популяции пилильщика происходит снижение аттрактивно- сти самок, поэтому растёт доля неоплодотворённых самок,и за счёт откладки ими инфертильных яиц увеличивается количество самцов в последующих поколениях пилильщика ( schonherr et al., 1979). Обобщая данные по соотношению полов в период I93I-I933 гг, можно
видеть, что в большинстве случаев количество самок в популяции пилильщика в эти годы в 1,5-2,3 раза превышало количество самцов» Такое соотношение полов для хвоегрызущих вредителей из отряда чешуекрылых характерно,скорее,для периода нарастания численности популяции, чем для периода её депрессии.
Такне нетрадиционны данные о плодовитости пилильщика в эти годы. Как уже говорилось ранее (см. Раздел 2*3), плодовитость пилильщика в годы наблюдений варьировала незначительно и достоверно не различалась по годам. Она была близка к средней плодовитости вида и не обнаруживала тенденций к снижению в первый послевспьшечный год. Эта особенность, вероятно, связана с участием в ежегодном лёте самых разных поколений пилильщика, вылетающих из длительно диапаузирующих коконов и выкармливавшихся в разные по условиям годы. Поэтому нам представляется невозможным использование данных о плодовитости пилильщика для суждения о снижении или нарастании численности популяции в его очагах*
Интересны данные о пораженности покоящихся фаз развития пилильщика (яиц и коконов) энтомофагами и болезнями в межвспы- шечные годы. Во все годы наблюдений эта пораженность была, в основном, высокой и на снижалась для яиц ниже 35%, а для коконов - ниже 43%. По этим данным также трудно судить об общих тен денциях изменения пораженности по мере течения лет после вспышки: в различных участках есть случаи и её снижения (участок № I для коконов), и её повышения (тот же участок, для яиц) с течением лет. То же можно сказать, анализируя данные о выживаемости пилильщика на разных фазах развития, которая варьировала исключительно широко и ненаправленно. Можно сделать вывод о том, что роль знтомофагов и болезней в динамике численности пилильщика, безусловно, во многих случаях очень высока. Однако проявление эффнкта действия знтомофагов и болезней в меквспышечные годы
имеет чаще всего не фоновый, а местный характер*
Коэффициент баланса популяции за год (от осени до осени), который в данном случае уместно рассматривать вместо общепринятого коэффициента размножения, указывает на относительно ста - бильную, хотя и невысокую, численность пилильщика в некоторых участках (К=1,19) или на её снижение ( К < I).
Из данных табл* 37 можно сделать важный вывод о пространственной изменчивости качественных и количественных параметров популяции обыкновенного соснового пилилыцика в пределах ком - пактного и небольшого лесного массива в один и тот же временный период..Она связана с мозаичностью экологических условий, имеющих место даже в относительно однородных биотопах. Зто создает дополнительные трудности при использовании качественных и количественных показателей состояния популяции для суждения о на - правлении движения численности пилильщика и прогнозировании вспышек его массового размножения.
Частоту вспышек обыкновенного соснового пилилыцика в Ростовской области можно охарактеризовать изменением площади очагов за последние 29 лет по области в целом и Каменскому лесхозу в частности. На рис* 24 показана относительная площадь очагов пилильщика по Ростовской области и Каменскому лесхозу в процентах от наибольшей за годы наблюдений. Последняя составила 44,1 тыс.га по области и 12,3 тыс. га по лесхозу. По рис. 24 хорошо заметны годы высокой численности пилилыцика. Общий рост площади очагов по области и по лесхозу с годами связан с увеличением площади сосновых культур на их территориях. Из рис. 24 можно видеть также , что периодичность проявления вспышек массового размножения пилильщика в Ростовской области составляет 3-6 лет. Примерно такую же продолжительность межвспышечного периода в 60-ые годы отмечают в Белоруссии Б.В.Рывкин (1967)
iso
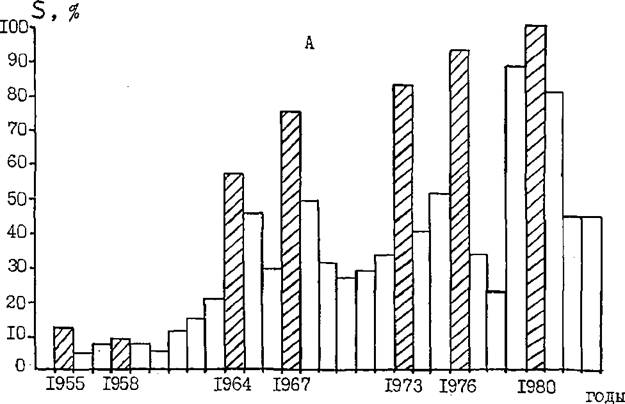
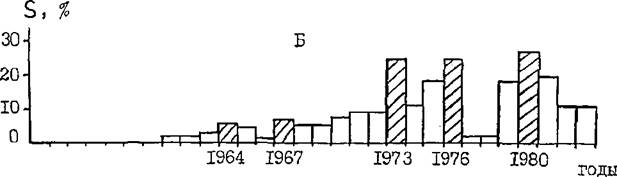
Вис, 24 , Динамика площади очагов обыкновенного соснового пилильщика (5) по Ростовской области (А) и Каменскому лесхозу (Б) с 1955 по 1983 гг. в % от наибольшей по области
и на Украине И«Д.Авраменко (1969). Имеются данные о кратковременности и бодее частой периодичности вспышек массового размножения и рыжего соснового пилильщика по сравнению с другими хвое- грызущими насекомыми из отряда чешуекрылых (Воронцов, Каюкина, 1961; Воронцов, 1932).
Возникновению частых вспышек массового размножения обыкновенного соснового пилильщика в условиях Ростовской области способствуют несколько обстоятельств, среди которых важнейшими являются: искусственное происхождение СОСНЯКОВ, почти ПОСТОЯННЫЙ дефицит влажности воздуха и почвы, а также сложившаяся практика посадки чистых культур сосны неширокими полосами (10-20 м); при этом создаются хорошо освещенные и прогреваемые насаждения с протяженными опушками. Названные обстоятельства являются причиной пониженной биологической устойчивости культур, в которых создается благоприятная экологическая обстановка для жизнедеятельности популяций пилильщика.
Продолжительность подъёма и спада численности, если судить по данным о площади очагов (рис. 24 ), составляет 1-3 года.Как показали наблюдения многих исследователей в различных точках ареала обыкновенного соснового пилильщика, где он даёт вспышки массового размножения, резкий подъём численности, сопровождающийся сильным повреждением насаждений, обычно наблюдается в течение только одного года с обязательным проявлением бивольтин- ного сезонного цикла и с наибольшей повреждаемостью кормовой породы личинками второй, летней генерации (Рывкин, 1963, 1966, 1967; Dusaussoy, Geri, 1971; Шаров, 1930; Eichhorn ,1932
и др.). Наши наблюдения и данные надзора по Каменскому лесхозу также подтверждают это положение. В обе последние вспыпки массового размножения, в 1976 и 1980 гг, обыкновенный сосновый пилиль щик развивался по бивольтинному типу и культуры заметно повреж-
132
дались дважды в течение лишь одного года.
Сопоставляя литературные данные об особенностях Развития вспышек массового размножения обыкновенного соснового пилильщика с результатами наших наблюдений за динамикой его популяции в меквспышечные годы, можно сделать вывод о том, что тип динамики численности пилильщика и её закономерности значительно отличаются от таковьк у хорошо известных представителей группы хвое- и листогрызущих насекомых из отряда чешуекрылых- Так, обычно у последних выделяются два предшествующие эруптивной фазе периода - начальная фаза вспышки и фаза нарастания численности, для которых характерно постепенное (из года в год) увеличение размера и веса особей, плодовитости самок, плотности популяции, а также низкая пораженность вида энтомофагами и болезнями (Ильине кий и др-, 1965)- Совершенно иначе развивается вспышка массового размножения у обыкновенного соснового пилильщика. Резкий подъём его численности, по нашему мнению, связан прежде всего с реактивацией основной части диапаузирующих особей в биотопе, плотность которых и определяет уровень подъёма численности первой и второй генерации пилильщика в год вспыики. Зимующий запас коконов в меквспышечные годы складывается из длительно диапаузирующих коконов и только что закоконировавшихся особей летнего поколения пилильщика. Варьирование плотности зимующего запаса коконов пилильщика в меквспышечные годы показано в табл* 37*
Из табл. 37 видно, что на третий год после вспышки плотность ко конов в отдельных участках достигала нескольких десятков на 1м2
В возможности возникновения вспышки массового размножения обыкновенного соснового пилильщика на основе реактивации диапа- узирующих особей можно убедиться по приведенному ниже теоретическому расчету, для которого использовались данные о выживаемости и плотности диапаузирующих коконов в годы исследования, а
S3
такие некоторые литературные данные об особенностях развития первого поколения пилильщика в годы вспышек. Многими иселедога- тєляіїи отмечается повышенная жизнеспособность популяции пилильщика первой генерации, что и учитывалось нами при выборе значений коэффициентов выживаемости для различных фаз и стадий развития пилильщика первой генерации. Плодовитость имаго пилильщика при весеннем лёте также установлена по литературным данным, а для имаго летней генерации - по фактическим данным ,• как средняя за годы исследований. Исходный осенний запао коконов взят по материалам наших учётов 1933 г. в участке '£ X, где их плотность была наиболее постоянна: 55,2 шт/ь£ или 8,6 шт. коконов на ЮОг хвои. Для удобства расчет представлен в виде таблицы 38.
Таблица 38
Расчет вероятной конечной плотности личинок старших возрастов обыкновенного соснового пилильщика при
бивольтинном сезонном цикле
| ! Показатели расчета ( х ) ! | "Г • I і | Единица {Значения учёта j z | ||
| I | ! 2 | I | з Г | 4. |
| I | Исходная ПЛОТНОСТЬ КОКОНОВ В'подстилке (осенний запас) в t -том ГОДУ | шт./м^ | 55,2 | |
| шт ./100 г хвои | 8,6 | |||
| 2 | Выживаемость зимующих коконов | доли ед. | 0,5 | |
| 3 | Плотность ЖИВЫХ коконов весной | шт./ХОО г | ||
| t + I года | хвои | 4,3 | ||
| 4 | Доля диапаузирующих коконов | % | 20 | |
| 5 | Плотность вылетевших весной имаго | шт./IOO г ХВОИ | 3,4 | |
| 6 | Соотношение ПОПОВ (o’ : 0 ) | ед. | 1:2 | |
продолжение таблицы 38
| I | 1 2 і | 3 ] | ! 4 |
| 7 | Плотность вылетевших весной самок | шт ./100 г хвои | 2,3 |
| 8 | Выживаемость имаго во время весеннею лёта | доли ед. | 0,3 |
| 9 | Плотность самок, отложивших яйца | шт./100 Г хвои | 0,69 |
| 10 | Плодовитость самок во время весеннего лёта | шт .яиц | 100 |
| II | Плотность яиц первой генерации | шт.яиц/ЮОг хвои | 69,0 |
| 12 | Выживаемость яиц первой генерации | доли ед. | 0,7 |
| ІЗ | Плотность личинок первой генерации первого возраста | шт./100 г хвои | 48,3 |
| 14 | Выживаемость личинок первой генерации | доли ед. | 0,8 |
| 15 | Плотность закоконировавшихся эонимф первой генерации | шт ./100 г хвои | 38,6 |
| 16 | Выживаемость коконов первой генерации в крона | доли ед* | 0,8 |
| 17 | Плотность ЖИВЫХ коконов первой генерации | шт ./100 г хвои | 30,9 |
| 18 | Соотношение ПОЛОВ ( о* : $ ) | ед. | 1:2 |
| 19 | Плотность самок в период летнего лёта | шт ./100 г хвои | 20,6 |
| 20 | Выживаемость имаго в период летне го лёта | шт ./100 г хвои | 0,3 |
| 21 | Плотность самок, отложивших яйца | шт ./100 г хвои | 6,2 |
| 22 Плотітость самок в период летнего | шт. яиц | 120 | |
продолжение таблицы 38
| I | 1 2 | ! 3 | Ї 4 |
| 23 | Плотность яиц второй генерации | шт ./100 г хвои | 741,9 |
| 24 | Выживаемость яиц второй генерации | доли ед. | 0,3 |
| 25 | Плотность личинок второй генера- | вт ./100 г | 222,6 |
| ции первого возраста | ХВОИ | ||
| 26 | Выживаемость личинок второй генерации | доли ед. | 0,5 |
| 27 | Конечная плотность личинок второй генерации старших возрастов в t + X году | шт./100 г хвои | 111,3 |
їаким образом, при условии вькода из диапаузы 80% особей пилилыцика от осеннего запаса коконов а при их фактически установленной за годы исследований зимней смертности равной 0,5 и соотношении полов 1:2, вылетевшие самки могут обеспечить наличие в кроне в конце вегетационного периода 111,3 личинок старших возрастов второй генерации на 100 г хвои. Предположительно это будет соответствовать сильному повреждению хвои с учетом частичного её предварительного повреждения личинками первой генерации (см. табл. 33). Поэтому при указанном выше фактическом осеннем запасе коконов на третий межвспышечный год и при перечисленных выше прочих условиях вспышка массового размножения обыкновенного соснового пилильщика вполне монет реализоваться уже в 1934 г.
Как видно из наблюдений в Ростовской области, бивольтинный цикл пилильщика в годы вспышек сменяется на хорошо выраженный моновольтинный цикл с одной летней волной лёта в межвспышечные годы. Зто возможно либо в результате постепенной смены соотношения форм с различными наследственно закрепленными фенологическими циклами в популяциях пилильщика в послевспышечные годы, либо в результате индукции диапаузы у подавляющей доли особей в
популяциях пилильщика под влиянием изменяющихся условий среды и эндогенных факторов. Как уже говорилось вше, более вероятна, по нашему мнению, смена моновольтинного жизненного цикла пилильщика на моновольтинный под.влиянием индукции диапаузы.
Экспериментально диапауза обыкновенного соснового пилильщика нами не изучалась. Как уже упоминалось ранее, исследованиями 0.8йхюрна ( Eichhorn , 1979, 1933), А.А«Шарова и А.Ф.Сафонки- на (1932) доказана тесная связь диапаузы пилильщика с фото - периодом и температурным режимом среды. Этим объясняется почти полное отсутствие диапаузы у развивающихся в первую половину лета при длинном дне особей весеннего поколения и наличие диапаузы Развивающихся во вторую половину лета при укороченном световом дне особей летнего поколения. А.А.Шаровыы и А.Ф.Сафонкиным выявлена диапауза обыкновенного соснового пилильщика и на стадии про нимфы. Ими отмечено, что не все зимующие пронимфы окукливаются весной, часть из них остается в коконах и окукливается только перед летним лётом. В годы исследований (I98I-I933 гг) при осенних учетах и анализе коконов пронимфы на были обнаружены, они появлялись, как это уже указывалось в разделе 2.5, за две недели до летнего лёта, с середины июля.
Механизм реактивации диапаузирующих особей пилильщика рассмотрен А.А.Шаровыы и А.Ф.Сафонкиным дифференцировано по отношению к особям весеннего и летнего поколений. При этом для каждого из них в лабораторных условиях экспериментально установлены оптимальные фотопериод и температура, при которых происходит выход особей из диапаузы. Тем не менее, представляется не убедительным сведение условий реактивации пилильщика из затяжной диапаузы только к определенным температуре и фотопериоду. Совершенно очевидно, что смена моновольтинного цикла развития на бивольтин - ный, совершающаяся благодаря массовому весеннему выходу пилильщи
ка из диапаузы, в природе происходит под воздействием более сложного комплекса условий внешней среды. Среди них должны быть обязательно какие-либо сильно варьирующие из года в год факторы. Температура среды, безусловно, относится к таким факторам, тогда как фотопериод для данного района неизменен из года в год. Пос- колы^ прекращение затяжной диапаузы у обыкновенного соснового пилильщика имеет определенную, хотя и не строго выраженную, периодичность, о которой можно судить по вспышкам массового разы ножения пилильщика (см. рис. 24 ), можно предположить, что оно связано с какими-либо периодически изменяющимися факторами внешней среды. Сради многих факторов среды в местообитаниях обыкновенного соснового пилильщика в Ростовской обл. наибольшей изменчивостью обладают погодные условия. Их принято называть модифицирующими факторами динамики численности насекомых. Погодные фак торы воздействуют как непосрэдственно на организм насекомого, так и косвенно на систему его биоценотических связей (Викторов, 1967; Воронцов, 1978 и др.). Кроме погодных факторов в местообитаниях обыкновенного соснового пилильщика из года в год меняется состояние кормовой породы, её резистентность к повреждениям, поврежденность и кормовые качества хвои. Именно поэтому имеет смысл установление связи возникновения и прекращения затяжной диапаузы обыкновенного соснового пилильщика прежде всего с погодой и состоянием кормовой породы.
Некоторые исследователи (Рывкин, 1963, 1967; Авраменко, 1969; Завада, 1976) отмечают тесную связь вспышек массового разы ножения обыкновенного соснового пилильщика с погодными условиями предыдущих лет. Одним из хороших критериев специфики погодных условий может служить гидротермический коэффициент Г.Т.Селянино- ва (сумма осадков, деленная на сумму температур, уменьшенную в 10 раз), характеризующий засушливость тех или иных периодов.
J33
Так, И.ЩїАвраменко (1969) считает, что массовому размножению пилилыцика предшествуют два относительно засушливую года, когда гидротермический коэффициент за вегетационный период меньше 1,0* Н. Оавада (1976) сделал вывод о том, что потенциальная угроза массового размножения пилильщика создаётся при условии, если гидротермический коэффициент (ГТК) августа-сентября предыдущего года близок к 0,5,
На примере последних 29 лет прослежена связь ГТК с годами вспышек массового размножения пилильщика в Ростовской области (рис. 25 ). По рис. 25 видно, что годам с наибольшей площадью очагов (1958, 1964, 1967, І9*ЇВ, 1976, 1980) предшествуют Годы с ІТК за вегетационный период близким к 0,5 (от 0,30 до 0,83).
Для характеристики роли метеорологической обстановки в возникновении вспышки массового размножения пилильщика можно использовать и другие показатели засушливости погоды: коэффициент водности за вегетационный период, определяемый как процент коли - чества осадков от среднего многолетнего, относительный дефицит влажности воздуха по БЖФпёрову, отражающей изменения температуры и влажности воздуха по отклонениям, в процентах, суммы среднемесячных дефицитов влажности воздуха на ІЗ ч. дня за вегетационный период от средних многолетних значений (табл. 39).
Все зти показатели характеризуют годы, предшествующие подьё мам численности пилильщика, как сухие и жаркие. Так, трём последним вспышкам размножения пилильщика предшествовали годы с количеством осадков за вегетационный период на 7,5 - 45,6% меньше нормы и с относительным дефицитом влажности воздуха на 40,4 - 66,7% вше нормы.
Рассматривая периодичность вспышек массового размножения пилильщика и повторяемость сухих и жарких вегетационных периодов в Ростовской области (рис. 24, 25 ), можно заметить, что послед
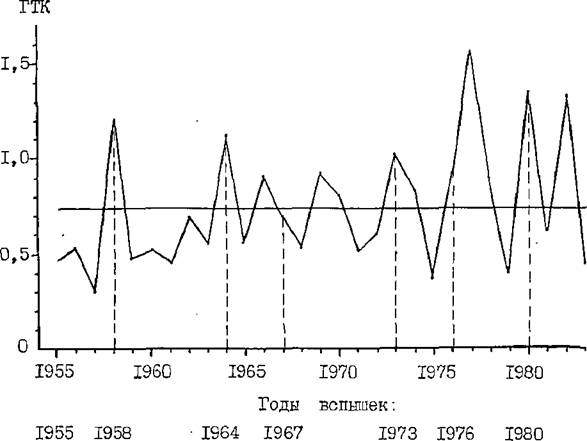
Рис. 25 . Значения гидротермического коэффициента Селянинова (ГТК) за 1955-1983 гг. в Каменском районе Ростовской области
W)
ние наблюдаются значительно чаще» Следовательно, не только они определяют периодичность подъёма численности пилильщика.
Таблица 39
Изменение некоторых метеорологических показателей за 1970-1933 гг в районе Каменского лесхоза (по данным Каменской метеостанции)
Мете оролэтические 5 і Me те орол огиче окне
показатели ; I показатели
-— --------------------------- ■---- г — -------------------------- гг--------
| Годы | ’Коэффициент {водности за ’вегетационный период, | ;Относительн.! {дефицит ! {влажности ’ {воздуха по J {Флёрову, { і; $ і | * 1 {Коэффициент { {водности за { ;вегетацион-f Іннй период,; І І | Относит, дефицит влажности воздуха по Флёрову, | |
| 1970 | 104,7 | 96,3 | 1977 | 209,0 | 80,7 |
| 1971 | 68,2 | 105,5 | 1978 | 110,6 | 80,7 |
| 1972 | 92,5 | 140,4 | 1979 | 56,4 | 130^ |
| 1973 | 127,9 | 67,0 | 1980 | 166,7 | 71,6 |
| 1974 | 102,8 | 89,0 | І98Г | 89,6 | 115,6 |
| 1975 | 54,4 | 145,9 | 1982 | 170,4 | 75,2 |
| }976 | 116,5 | 81,7 | 1983 | 72,0 | 119,2 |
Другим циклическим процессом з местообитаниях обыкновенного соснового пилильщика в Ростовской области является периодическое повреждение хвои сосны в годы вспышек и восстановление хвои в межвспышечные годы (см. главу 4). Период восстановления хвои длится, как показали наши наблюдения, большей частью три, иногда даже два года. Поскольку периодичность вспышек массового размножения пилильщика в Ростовской области равна 3-6 годам, можно предположить, что она в значительной мере определяется влиянием на популяцию пилильщика самой кормовой породы.
Влияние кормовой породы на популяцию палильщика многогранно. Во-первых і имеет значение общая резистентность сосны к поврежде-
ш
ниям. От неё зависит выживаемость питающихся на хвое насекомых, в особенности личинок первых возрастов. Кроме того, большое значение имеет биохимический состав хвои. Он меняется у сосны на протяжении вегетационного периода (Ижевский, 1967; Васильева, 1973), и в связи с нарушением метаболизма дерева при его дефолиации, и после восстановления хвои (Гире, 1982; Плешанов,1932).
У обыкновенного соснового пилильщика личинки первой генерации питаются преимущественно старой, а личинки второй генерации- - и старой, и молодой хвоей текущего года. По наблюдениям Стадницкого, проводившего выкармливание личинок рыжего соснового пилильщика на хвое разного возраста, старая хвоя сосны содержит большее количество пластических веществ (сахаров, крахмала,геми- целлюлозы). При выкармливании личинок пилильщика на старой хвое диапауза его была наименьшей (около 8%), а выживаемость - высокой (около 85$). При выкармливании личинок на молодой хвое их гибель достигала 40$, а вес коконов был значительно меньшим,чем при питаний старой хвоей (Стадницкий, 1969).
Меняются пищевые свойства хвои у деревьев во время и после их дефолиации. По мере увеличения степени дефолиации-содержание углеводов в оставшейся хвое снижается. Отрастающая хвоя на поврежденных деревьях также обеднена углеводами (Гире, 1982), что значительно снижает жизнеспособность питающихся ею насекомых - филлофагов (Плешанов, 1982). Поэтому питание личинок летних поколений в год вспышки и в межвспышечные годы на ранее повреждавшихся деревьях и питание восстановившейся хвоёй снижает их жизнеспособность . и, быть может,является сигналом о неблагополучии условий среды; Зто сказывается на увеличении доли длительно диа- паузирующих особей в популяции. Можно предположить, что недостаток корма, крторый испытывает летнее поколение пилильщика в год вспышки, и неблагоприятные пищевые качества хвои определяют
протяженность затяжной диапаузы пилильщика, соразмерную с наиболее вероятным периодом полного восстановления хвои в поврежденных насаждениях* Это приспособление могло выработаться у пилилыцика в процессе коэволюции с его кормовой породой путем отбора определенных экологических типов вида, наиболее приспособленных к условиям конкретною региона*
Таким образом, массовый весенний выход пилилыцика из диапаузы происходит и вспышка реализуется не раньше, чем спустя два-три года после окончания последней вспышки, во время которых восстанавливается масса и пищевая ценность хвои поврежденных сосен. Кроме того, необходимым условием является жаркая и сухая погода вегетационного периода предшествующего вспышке года, которая воздействует на популяцию пилильщика и резистентность его кормовой породы.
Важным по своему значению фактором, воздействующим на уровень численности обыкновенного соснового пилильщика, является также комплекс знтомофагов, который почти постоянно снижает плотность популяции пилилыцика на всех фазах его развития, в особенности на фазе яйца и стадии эонимфы в коконе. Меньшее значение имеют болезни пилильщика. По данным А.АДарова (1982),игра ют роль не только плотность популяции знтомофагов и их состав,но и сопряженность сезонного цикла паразитов с сезонным циклом хозяина,которая различается в годы вспышек и межвспышечные годы. Как видно из полученных в процессе исследования данных,естественные враги и болезни пилильщика в межвспышечные годы в отдельных участках существенно снижали уровень его численности, а иногда являлись основной причиной гибели пилильщика на том или ином этапе жизненного цикла. Благодаря большой неравномерности их распределения даже в пределах относительно однородных местообитаний пилильщика, имела место различная их эффективность в
один и ют ко отрезок времени. Это увеличивало изменчивость степени сопротивления среды в местообитаниях пилилыцика и способствовало увеличению варьирования уровня плотности его популяций.
В свою очередь Различная плотность популяции пилилыцика в пределах относительно однородных изолированных массивов сосновых куль тур, наблюдавшаяся в те или иные годы, обеспечижла неравномерность последующей степени их повреждения дане при общем подъёме численности пилильщика (см. рис. 21, 22 ). Таким образом, увеличивалась возможность совместного длительного сосуществования всех членов биогеоценоза в степных посадках сосны: и кормовой породы, и обыкновенного соснового пилильщика, и его естественных врагов.
Приведенные выше особенности динамики численности обыкно - венного соснового пилильщика в Ростовской области долины быть приняты во внимание при организации надзора в его очагах и прогнозировании вспышек массового размножения.
ш
Еще по теме 5. ДИНАМИКА ЧИСЛЕННОСТИ ПОВДИКД В СОСНОВЬК КУЛЬТУРАХ СТЕПНОЙ зоны и ФАКТОРЫ'ЕЕ ОПРЕДЕЛЯЮЩИЕ:
- 6.1. Характер распределения пилильщика в сосновьк культурах и методы его учёта
- Изучение влияния пролина (ростового фактора) на рост культуры Br. flavum 194(Рго ) и динамику биосинтеза аргинина в ферментере
- Динамика численности населения (N) РФ с 2000 по 2004гг.
- Харлашина, Анна Владимировна. Экология и лесохозяйственное значение обыкновенного соснового пилильщика (Diprion ріпі L.) в сосняках степной зоны Диссертация на соискание учёной степени кандидата биологических наук. Москва, 1984, 1984
- Спрос и факторы, его определяющие
- Факторы, определяющие сенсибилизацию
- Факторы, влияющие на организационную культуру компаний.
- Вопрос 3. Экономические границы отрасли и факторы, их определяющие
- 13.Факторы, определяющие эффективность психокоррекции.
- Эластичность предложения, факторы, ее определяющие.
- Факторы, определяющие чувствительность и специфичность
- Факторы, определяющие прогноз у больных с инфарктом миокарда
- Факторы, определяющие исход и значение местного малокровия
- Генетические факторы, определяющие эффективность химиотерапии