3.2.2.Половозрелые крысы.
Как указывалось ранее, для выявления влияния барбитуратов на клетки ядер основных отделов МТ головного мозга в эксперименте были также использованы половозрелые животные, которые получали фенобарбитон в дозе 30 мг/кг.
Эксперимент с воздействием фенобарбитона в указанной дозе длился 7, 15, 30 и 60 суток. Результаты, полученные в эксперименте, затем сравнивались с показателями аналогичных ядер отделов МТ головного мозга крыс контрольных групп и с результатами, полученными в эксперименте с неполовозрелыми крысами.При микроскопии препаратов указанной экспериментальной группы (IIФ1) на 7 сутки эксперимента, в отличие от контроля, у большинства нейронов всех ядер обоих групп МТ головного мозга выявлены такие же изменения обратимого характера, как и в группах неполовозрелых животных: увеличение размеров тела и ядра клеток, околоядерный хроматолиз. При этом тела клеток округляются, контуры их нечеткие, ядра в некоторых нейронах принимают краевое расположение, просветлены, ядрышки расположены эксцентрично. В отдельных клетках отмечаются признаки острого набухания – вакуолизация цитоплазмы, перицеллюлярный отек. Со стороны макроглии отмечается усиление пролиферативных процессов (рис. 3.30-3.31).
Морфометрически полученные данные показали, что в данной экспериментальной группе животных по сравнению с контрольной на 7 сутки в базо-латеральном отделе МТ головного мозга половозрелых крыс происходило увеличение площади сечения тел клеток с 75,71 ± 0,1 мкм2 до 87,26 ± 0,09 мкм2 (t = 2,48; Р < 0,05), уменьшение количества нейронов с 1526,82 ± 4 до 1451,61 ± 3,25 (t = 3,17; Р < 0,05); увеличение количества общей глии с 1657,26 ± 2,39 до 2351,94 ± 2,95 (t = 3,24; Р < 0,05), сателлитной глии с 809,14 ± 3,11 до 858,06 ± 2,39 (t = 5,64; Р < 0,05) и интерглиального коэффициента с 0,34 ± 0,06 до 0,45 ± 0,04 (t = 4,06; Р < 0,05). Эти изменения носили статистически достоверный характер.
Изменения остальных показателей не являлись достоверными. В кортико-медиальном отделе МТ головного мозга на 7 сутки эксперимента происходило увеличение площади тела клеток с 63,6 ± 0,11 мкм2 до 88,59 ± 0,11 мкм2 (t = 4,187; Р < 0,05) и их ядер с 4,05 ± 0,03 мкм2 до 4,76 ± 0,03 мкм2 (t = 2,52; Р < 0,05). Как видно, изменения цифровых показателей носили статистически достоверный характер. По остальным показателям изменения не достигали уровня достоверности.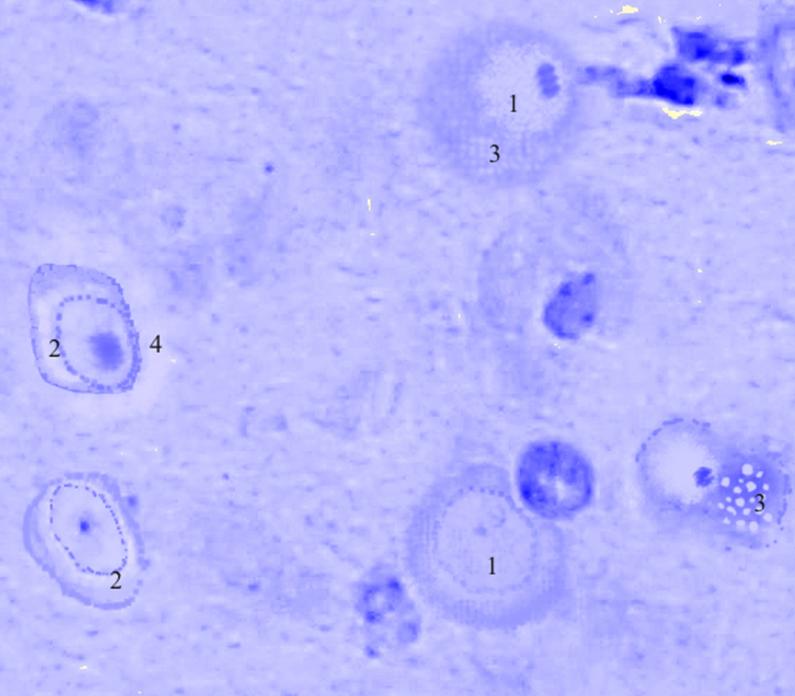
Рис. 3.30. Микрофотография. Кортикальное ядро кортико-медиального отдела миндалевидного тела головного мозга половозрелой крысы на 7 сутки эксперимента с фенобарбитоном в дозе 30 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Округлая форма клеток.
2. Околоядерный хроматолиз.
3. Вакуоляризация цитоплазмы.
4. Перицеллюлярный отек.
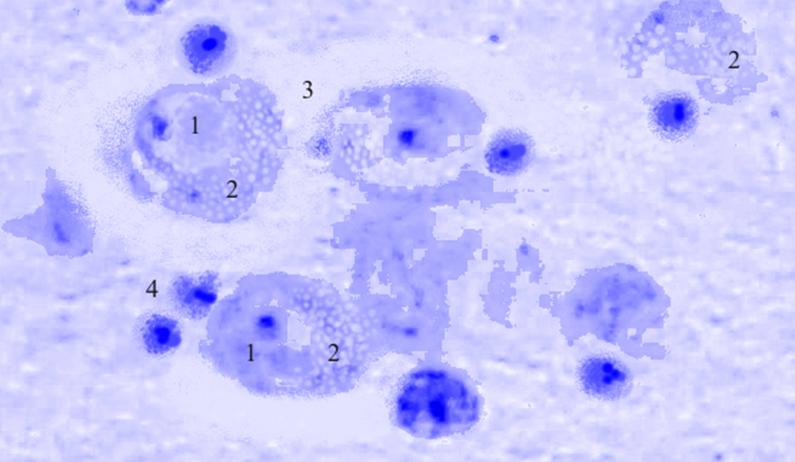
Рис. 3.31. Микрофотография. Латеральное ядро базо-латерального отдела миндалевидного тела головного мозга половозрелой крысы на 7 сутки эксперимента с фенобарбитоном в дозе 30 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Увеличение площади клетки.
2. Вакуоляризация цитоплазмы.
3. Перицеллюлярный отек.
4. Пролиферация глии.
Вышеприведенные данные свидетельствуют, что на 7-ые сутки эксперимента статистически достоверные изменения цифровых показателей нейронов ядер основных отделов МТ головного мозга половозрелых крыс наблюдались лишь в значениях площадей клеток в обоих указанных отделах, а ядер клеток – только в кортико-медиальном отделе. Количество нейронов статистически достоверно уменьшалось только в базо-латеральном отделе.
Показатели, характеризующие количество клеток глии в ядрах указанных отделов МТ статистически достоверно изменялись только в базо-латеральном отделе.
Следует отметить, что на 7-е сутки наиболее выраженная реакция нейронов наблюдалась в кортико-медиальном отделе МТ, а реактивность глии – в базо-латеральном отделе.В средние сроки эксперимента (15 суток) вышеперечисленные неспецифические изменения нервных клеток изучаемых ядер сохранились. В этот период наблюдались также гиперхромные клетки при наличии клеток с гидропическими изменениями (набухшие, имеющие вакуоли в цитоплазме), с явлениями хроматолиза, величина которого увеличилась и распространилась на большую часть клеточного тела. Однако появились единичные нейроны, в которых преобладали уже дистрофические изменения в виде сморщивания. Такие клетки уменьшены в размерах, имели вытянутые контуры тела. Иногда на значительном расстоянии прокрашивались отростки нервных клеток. Нередко вокруг нейронов группировались (до 5 и более) клетки глии (рис. 3.32-3.33). Полученные при микроскопии визуальные данные подтверждались данными морфометрии. Так, в базо-латеральном отделе МТ статистически достоверный характер носили изменения плотности общей глии с 2361,13 ± 3,09 до 3086,02 ± 2,11 (t = 7,24; Р < 0,05), сателлитной глии с 829,09 ± 3,39 до 1338,71 ± 2,01 (t = 5,1; Р < 0,05), а также перинейронального индекса с 0,46 ± 0,07 до 0,6 ± 0,03 (t = 2,112; Р < 0,05). Изменения других цифровых показателей не достигали достоверных величин. В ядрах кортико-медиального отдела МТ на 15-е сутки наиболее существенные изменения отмечались по следующим показателям: увеличивалась площадь клеток с 67,26 ± 0,08 до 97,22 ± 0,05 (t =
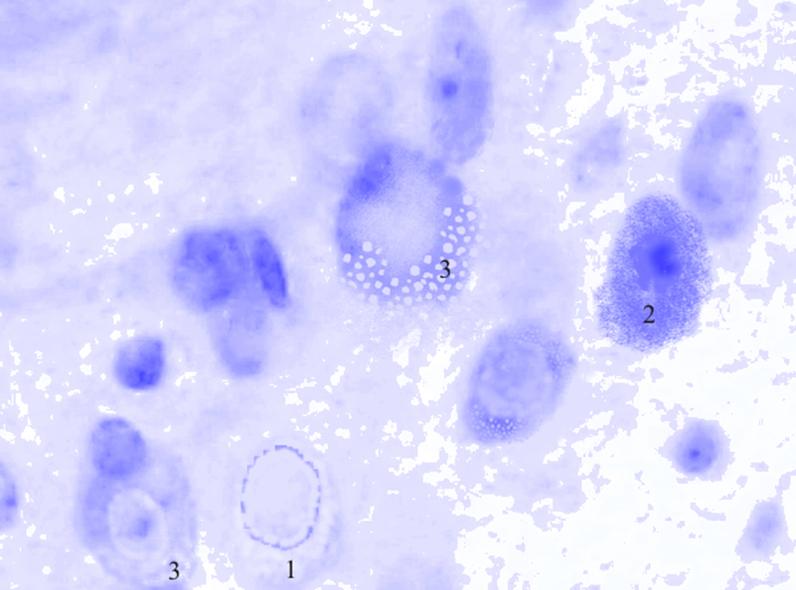
Рис. 3.32. Микрофотография. Латеральное ядро базо-латерального отдела миндалевидного тела головного мозга половозрелой крысы на 15 сутки эксперимента с фенобарбитоном в дозе 30 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Тотальный хроматолиз.
2. Гиперхроматоз.
3. Вакуоляризация цитоплазмы.
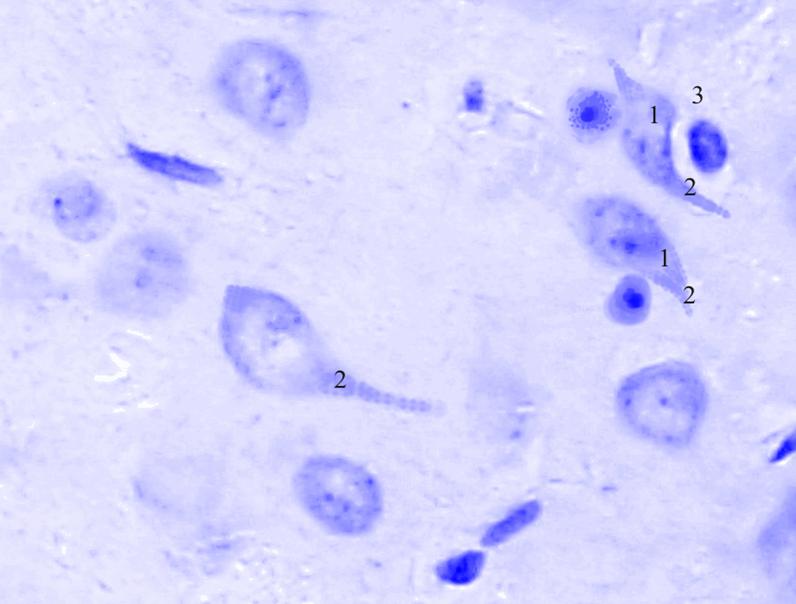
Рис.
3.33. Микрофотография. Центральное ядро кортико-медиального отдела миндалевидного тела головного мозга половозрелой крысы на 15 сутки эксперимента с фенобарбитоном в дозе 30 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.1. Сморщивание.
2. Утолщение отростков.
3. Сателлитоз.
3,0; Р < 0,05) и их ядер с 4,76 ± 0,03 до 6,93 ± 0,02 (t = 2,17; Р < 0,05), а также ядерно-цитоплазматическое отношение с 0,08 ± 0,004 до 0,1 ± 0,003 (t = 2,707; Р < 0,05). Со стороны глии отмечалось увеличение количества сателлитной глии с 848,82 ± 2,2 до 1190,86 ± 2,06 (t = 3,42; Р < 0,05), перинейронального индекса с 0,49 ± 0,07 до 0,59 ± 0,03 (t = 2,244; Р < 0,05) и интерглиального коэффициента с 0,35 ± 0,05 до 0,48 ± 0,03 (t = 2,127; Р < 0,05). По остальным показателям статистически достоверная разница не обнаружена. Таким образом, на 15-е сутки статистически достоверные изменения изучаемых показателей нейронов происходили в ядрах кортико-медиального отдела МТ головного мозга половозрелых крыс, а существенные изменения показателей глии наблюдались в ядрах обоих отделов МТ. Из вышеприведенного следует, что, видимо, наиболее ранимыми были ядра кортико-медиального отдела МТ головного мозга крыс половозрелого возраста, где появлялись статистически достоверные изменения цифровых показателей и нейронов, и глии.
Вышеперечисленные морфологические изменения в ядрах основных отделов МТ головного мозга крыс данной возрастной группы особенно ярко были выражены в более длительные сроки эксперимента, то есть на 30 сутки воздействия фенобарбитона. В ряде нервных клеток изучаемых ядер МТ изменения носили обратимый характер: размеров ядра и ядрышка, их эктопия, хроматолиз околоядерный и тотальный, гиперхроматоз, набухание, наличие мелких и крупных вакуолей в цитоплазме, перицеллюлярный отек. В то же время накапливались и дистрофически измененные клетки: сморщенные клетки – уменьшенные в размерах, с четкими контурами и вытянутой веретенообразной формой. Ядра таких нейронов часто палочковидные, плохо или совсем неразличимы.
На отдельных препаратах обнаруживались явления нейронофагии, клетки-тени. Вокруг нервных клеток нередко отмечалось увеличение количества макроглии (рис. 3.34-3.35).На 30 сутки эксперимента в ядрах базо-латерального отдела МТ головного мозга половозрелых крыс наблюдалось увеличение площади ядер нервных клеток с 3,64 ± 0,02 до 5,04 ± 0,05 (t = 3,67; Р < 0,05), уменьшение
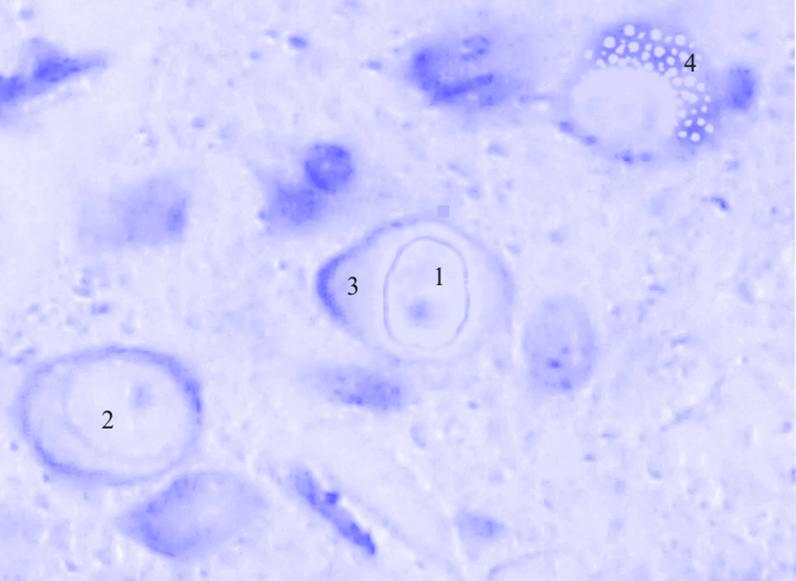
Рис. 3.34. Микрофотография. Базальное ядро базо-латерального отдела миндалевидного тела головного мозга половозрелой крысы на 30 сутки эксперимента с фенобарбитоном в дозе 30 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Увеличение площади клетки и ядра.
2. Краевое расположение ядра и ядрышка.
3. Тотальный хроматолиз.
4. Вакуоляризация цитоплазмы.
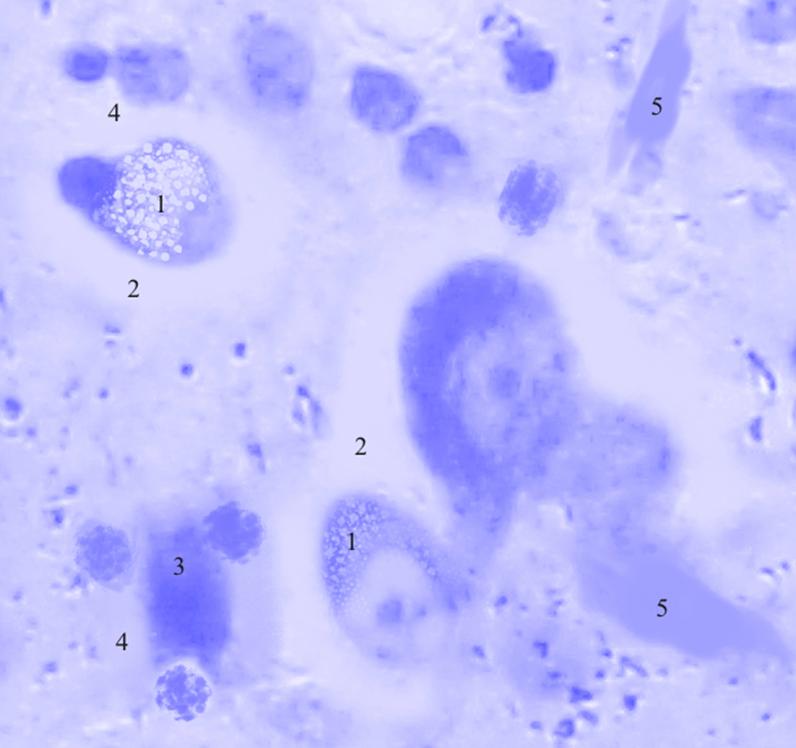
Рис. 3.35. Микрофотография. Центральное ядро кортико-медиального отдела миндалевидного тела головного мозга половозрелой крысы на 30 сутки эксперимента с фенобарбитоном в дозе 30 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Вакуолизация цитоплазмы.
2. Перицеллюлярный отек.
3. Сморщивание нейронов.
4. Сателлитоз.
5. Клетки-тени.
плотности нейронов с 1561,24 ± 4,83 до 1053,75 ± 6,53 (t = 2,896; Р < 0,05); со стороны глии – увеличение плотности общей глии с 2236,67 ± 3,64 до 3833,33 ± 2,86 (t = 2,45; Р < 0,05) и интерглиального коэффициента с 0,35 ± 0,07 до 0,54 ± 0,09 (t = 2,538; Р < 0,05). Указанные изменения являлись статистически достоверными. По всем остальным морфометрическим показателям изменения носили недостоверный характер. В кортико-медиальном отделе МТ отмечено увеличение площади клеток с 63,6 ± 0,1 до 95,01 ± 0,1 (t = 5,128; Р < 0,05) и их ядер с 4,23 ± 0,04 до 5,1 ± 0,05 (t = 1,959; Р < 0,05), уменьшение ядерно-цитоплазматического отношения с 0,08 ± 0,006 до 0,06 ± 0,009 (t = 2,513; Р < 0,05), увеличение плотности общей глии с 2361,29 ± 4,84 до 2902,02 ± 4,05 (t = 2,715; Р < 0,05), сателлитной глии с 909,68 ± 3,33 до 1458,33 ± 5,67 (t = 3,933; Р < 0,05) и перинейронального индекса с 0,41 ± 0,05 до 0,51 ± 0,09 (t = 2,339; Р < 0,05).
Как видно, по всем вышеперечисленным показателям изменения носят статистически достоверный характер (изменения других изучаемых показателей статистически недостоверны). Таким образом, на 30 сутки эксперимента статистически достоверные изменения цифровых показателей нейронов характерны для ядер обоих отделов МТ головного мозга крыс половозрелого возраста. Существенные изменения показателей глии наблюдались в ядрах обоих отделов МТ. При этом наиболее ранимыми представлялись ядра кортико-медиального отдела, где статистически достоверных изменений цифровых показателей было выявлено больше.При увеличении сроков эксперимента до 60 суток в нервной ткани изучаемых ядер МТ у половозрелых животных наблюдались признаки и репаративного процесса. Морфология ряда нейронов нормализовалась: исчезали признаки набухания, отмечалось новообразование нисслевского вещества (прежде всего в перинуклеарной зоне), восстанавливались ядро и ядрышко. Со стороны глии отмечались пролиферация и активация макроглии, перинейрональный сателлитоз (рис. 3.36). На 60-е сутки в базо-латеральном отделе МТ происходило увеличение площади сечения нейронов с 72,9 ± 0,08 до 79,4 ± 0,06 (t = 5,652; Р < 0,05) и их ядер с 3,62 ± 0,02 до 4,12 ± 0,01 (t = 3,754; Р
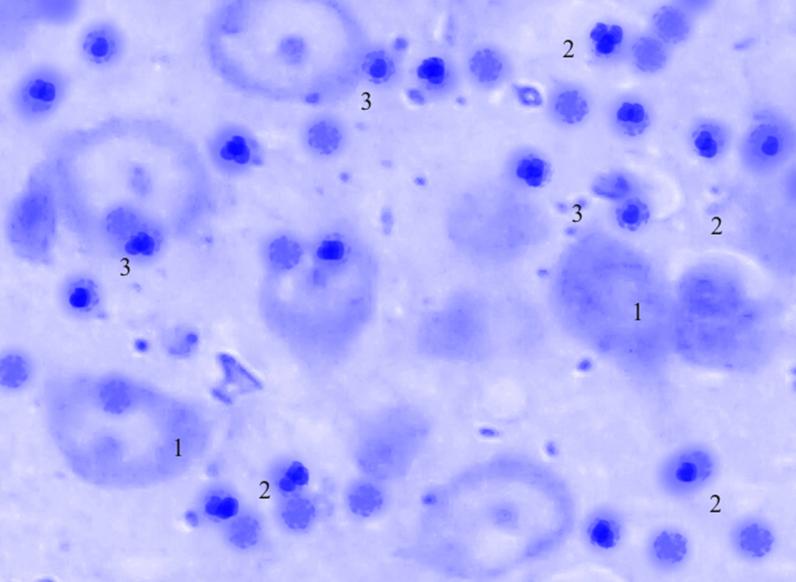
Рис. 3.36. Микрофотография. Базальное ядро базо-латерального отдела миндалевидного тела головного мозга половозрелой крысы на 60 сутки эксперимента с фенобарбитоном в дозе 30 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Гиперхроматоз.
2. Активация и пролиферация глии.
3 Сателлитоз.
< 0,05), количества общей глии с 1894,92 ± 3,85 до 2107,54 ± 6,34 (t = 4,865; Р < 0,05), количества сателлитной глии с 687,6 ± 3,09 до 954,72 ± 4,1 (t = 2,976; Р < 0,05), перинейронального индекса с 0,47 ± 0,08 до 0,68 ± 0,05 (t = 1,187; Р < 0,05) и интерглиального коэффициента с 0,34 ± 0,06 до 0,47 ± 0,03 (t = 2,298; Р < 0,05). Изменения носили достоверный характер (по остальным изучаемым показателям изменения статистически недостоверны). В кортико-медиальном отделе МТ наблюдалось существенное уменьшение количества нейронов с 1410,14 ± 2,89 до 1246,17 ± 3,75 (t = 3,153; Р < 0,05), увеличение плотности общей глии с 1658,99 ± 2,63 до 2064,83 ± 5,13 (t = 3,174; Р < 0,05), сателлитной глии с 668,2 ± 2,12 до 1004,18 ± 3,22 (t = 2,285; Р < 0,05), глиального индекса с 1,22 ± 0,08 до 1,45 ± 0,09 (t = 2,396; Р < 0,05) и перинейронального индекса с 0,48 ± 0,05 до 0,67 ± 0,02 (t = 2,407; Р < 0,05). По остальным морфометрическим показателям изменения не достигали достоверности.
Для более глубокого анализа обнаруженных нами в эксперименте морфологических изменений был использован корреляционный анализ. С его помощью выяснялось, как изменится взаимосвязь показателей, характеризующих состояние ядер основных отделов МТ головного мозга крыс половозрелого возраста при воздействии фенобарбитона в дозе 30 мг/кг. Анализ включал 2 изучаемых отдела МТ и 9 показателей клеток изучаемой структуры головного мозга.
При исследовании корреляционных связей между ядрами базо-латерального и кортико-медиального отделов МТ головного мозга половозрелых крыс при интоксикации фенобарбитоном в дозе 30 мг/кг длительностью 7 суток взаимосвязь средней силы (0,3 < r < 0,7) отмечалась лишь по показателю плотности общей глии. По остальным показателям наблюдали слабую взаимосвязь (r < 0,3). Взаимосвязей сильной степени между исследуемыми отделами МТ при указанных условиях эксперимента мы не наблюдали.
На 15 сутки эксперимента сильная корреляционная связей (0,7 < r < 1,0) между отделами МТ была обнаружена только по показателю площади сечения нейронов, а средней силы связи (0,3 < r < 0,7) были установлены по показателям плотности общей глии, перинейронального индекса и глиального индекса. По остальным показателям была выявлена слабая взаимосвязь (r < 0,3).
При эксперименте с длительностью влияния фенобарбитона в дозе 30 мг/кг 30 суток связи средней силы (0,3 < r < 0,7) были отмечены по показателям плотности нейронов, глиального индекса и интерглиального коэффициента. Остальные связи были зафиксированы как слабые. Сильных связей между исследуемыми параметрами, как и в предыдущие сроки эксперимента, выявлено не было.
На 60 сутки эксперимента сильная взаимосвязь (0,7 < r < 1,0) обнаруживалась между ядрами основных отделов МТ головного мозга по плотности общей глии, сателлитной глии и по показателю интерглиального коэффициента. Средней силы связи (0,3 < r < 0,7) были выявлены по показателям площади сечения нейронов, плотности нейронов и показателю глиального индекса. По остальным показателям установлена слабая взаимосвязь (r < 0,3).
Сравнивая корреляционные связи ядер обоих отделов МТ головного мозга половозрелых крыс контрольных и экспериментальных групп, можно отметить, что происходит изменение силы взаимосвязей, как по количеству, так и по качеству.
Из анализа корреляционных связей можно сделать следующие выводы:
1. воздействие фенобарбитона в дозе 30 мг/кг на ядра основных отделов МТ головного мозга половозрелых крыс изменяет характер и силу взаимосвязей между основными отделами МТ головного мозга;
2. по сравнению с контрольными группами во все сроки эксперимента изменяется характер и сила взаимосвязей между ядрами обоих отделов МТ в сторону ослабления в экспериментальных группах;
3. сильные взаимосвязи между отделами МТ головного мозга отмечались только при длительности эксперимента 60 суток;
4. средних связей было обнаружено больше, чем сильных, однако, меньше, чем слабых (15 и 30 суток);
5. слабые взаимосвязи между отделами МТ головного мозга сохранялись во все сроки эксперимента.
Для выявления влияния барбитуратов на нервную ткань ядер отделов МТ головного мозга половозрелых крыс, экспериментальные животные получали фенобарбитон и в дозе 70 мг/кг. Результаты исследования также анализировались и сравнивались с показателями изучаемых ядер МТ головного мозга крыс контрольных групп.
Получены следующие данные. На препаратах указанной экспериментальной группы (IIФ2/7) в отличие от контрольной на 7 сутки изменения имели те же черты, что и при интоксикации фенобарбитоном в меньшей дозе (30 мг/кг), но носили более выраженный характер. Это выражалось выявлением большего количества клеток с признаками неспецифических морфологических преобразований, а именно хроматолиза (особенно околоядерного) и гидропических изменений. Нередко определялись нейроны с явлениями острого набухания, такими как: увеличение размеров клеточного тела, округление клеток, увеличение и просветление ядра, которое иногда занимало эксцентрическое положение. В некоторых нервных клетках наблюдалось утолщение отростков, и они становились видимыми на значительном протяжении от тела клетки. Со стороны макроглии отмечалось усиление пролиферативных процессов и перераспределение ее вокруг нейронов, что также может свидетельствовать о повышении функциональных потребностей нейронов (рис. 3.37-3.38).
Полученные при микроскопии данные подтверждались результатами морфометрии и статистического анализа. Так, на 7 сутки эксперимента по сравнению с контролем в базо-латеральном отделе МТ увеличивались площади клеток с 75,71 ± 0,1 до 90,16 ± 0,07 (t = 4,235; Р < 0,05) и их ядер с 4,16 ± 0,02 до 4,74 ± 0,05 (t = 4,046; Р < 0,05). Со стороны глии было выявлено увеличение количества общей глии с 1657,26 ± 2,39 до 2381,82 ± 5,77 (t = 3,092;
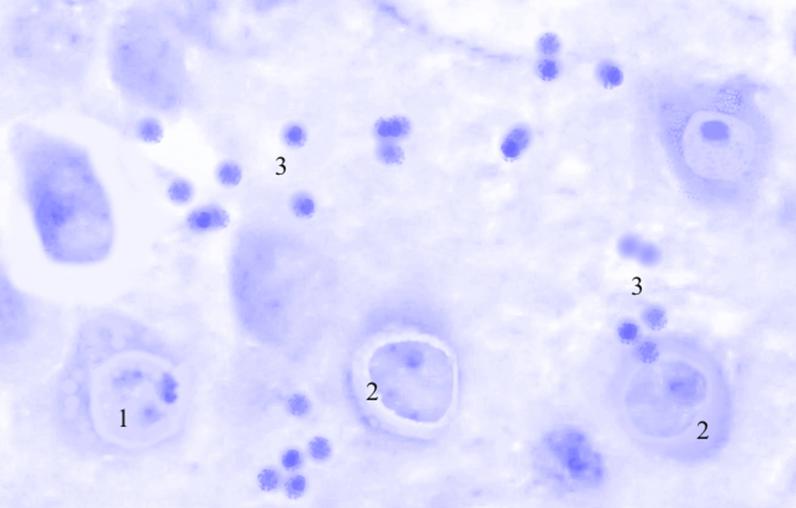
Рис. 3.37. Микрофотография. Кортикальное ядро кортико-медиального отдела миндалевидного тела головного мозга половозрелой крысы на 7 сутки эксперимента с фенобарбитоном в дозе 70 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Увеличение в размерах клеток и ядер.
2. Хроматолиз.
3. Пролиферация глии.
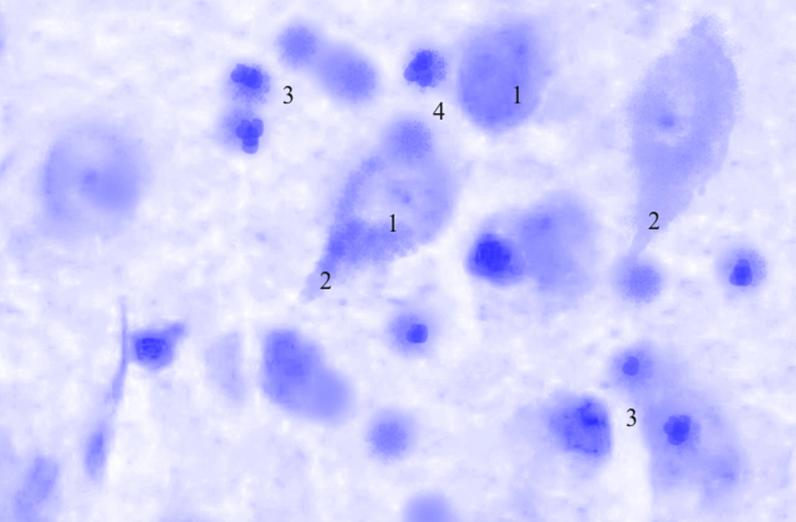
Рис. 3.38. Микрофотография. Латеральное ядро базо-латерального отдела миндалевидного тела головного мозга крысы половозрелого возраста на 7 сутки эксперимента с фенобарбитоном в дозе 70 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Гиперхроматоз.
2. Утолщение отростков.
3. Пролиферация глии.
4. Сателлитоз.
Р < 0,05) и глиального индекса с 1,55 ± 0,08 до 1,7 ± 0,09 (t = 1,579; Р < 0,05). Указанные изменения носили статистически достоверный характер. Остальные показатели, характеризующие нервные клетки и глию, не являлись достоверными. В кортико-медиальном отделе МТ головного мозга происходило увеличение площади сечения клеток с 63,6 ± 0,11 до 90,89 ± 0,38 (t = 4,66; Р < 0,05) и их ядер с 4,05 ± 0,01 до 4,89 ± 0,05 (t = 2,474; Р < 0,05), а также уменьшение плотности нейронов с 1526,82 ± 4 до 1289 ± 4,6 (t = 3,616; Р < 0,05). Со стороны глии мы наблюдали увеличение количества клеток макроглии с 790,32 ± 3,62 до 1363,63 ± 3,11 (t = 3,727; Р < 0,05) и перинейронального индекса с 0,69 ± 0,08 до 0,86 ± 0,03 (t = 2,838; Р < 0,05). По остальным показателям изменения не достигали статистической достоверности.
Как видно из вышеперечисленного, при исследовании нервной ткани ядер МТ при воздействии фенобарбитона в дозе 70 мг/кг длительностью 7 суток статистически достоверные изменения нейронов отмечались в обоих отделах МТ головного мозга, но более выражены в кортико-медиальном отделе. Выраженная реакция глии была также обнаружена в обоих отделах МТ. Таким образом, наиболее ранимыми были ядра кортико-медиального отдела МТ головного мозга крыс, где статистически значимых изменений было больше.
В средние сроки эксперимента (15 суток) вышеуказанные неспецифические морфологические изменения нейронов изучаемых ядер МТ сохранялись. Наряду с этим появлялись нервные клетки, в которых преобладали дистрофические изменения в виде сморщивания. Основными характеристиками таких изменений были уменьшение клеточного тела и гиперхроматоз. Клетки были уменьшены в размерах, их контуры становились более четкими, а боковые поверхности западали. Ядро изменяло форму до палочковидной, становилось плохо, а иногда совсем неразличимо. Глыбки тигроида сближались, сливались в компактные, темно окрашенные конгломераты. В конечной стадии этого процесса нервные клетки имели вид узких, темных, веретеновидных, палочковидных или угловатых образований. Нередко вокруг таких клеток группировались клети глии и наблюдались процессы нейронофагии (рис. 3.39-3.40). При этом на 15 сутки в базо-латеральном отделе МТ со стороны показателей, характеризующих нейроны, происходило увеличение площади сечения клеток с 64,95 ± 0,08 до 85,38 ± 0,06 (t = 7,22; Р < 0,05). А со стороны глии – увеличение количества сателлитной глии с 829,09 ± 3,39 до 983,87 ± 1,57 (t = 3,733; Р < 0,05), уменьшение глиального индекса с 1,38 ± 0,1 до 1,18 ± 0,07 (t = 2,549; Р < 0,05) и увеличение интерглиального коэффициента с 0,34 ± 0,06 до 0,41 ± 0,03 (t = 3,964; Р < 0,05). Как видно, изменения вышеуказанных параметров носили статистически достоверный характер. Остальные изучаемые цифровые показатели не отличались от контрольных, то есть практически были одинаковы. В кортико-медиальном отделе МТ на 15 сутки эксперимента наблюдались со стороны нервных клеток увеличение площади сечения нейронов с 67,26 ± 0,08 до 93,15 ± 0,07 (t = 6,086; Р < 0,05) и их ядер с 4,76 ± 0,03 до 5,08 ± 0,01 (t = 2,33; Р < 0,05), а также уменьшение количества нейронов с 1911,1 ± 4,83 до 1802,48 ± 2,49 (t = 4,335; Р < 0,05). Реакция глии выражалась следующими измененими: увеличение количества общей глии с 2437,85 ± 3,31 до 2807,85 ± 1,47 (t = 3,944; Р < 0,05) и сателлитной с 848,82 ± 2,2 до 1006,72 ± 1,43 (t = 2,457; Р < 0,05). Изменения лишь этих параметров носили достоверный характер, а другие – не достигали величины статистической достоверности. Следовательно, на 15 сутки эксперимента оба изучаемых отдела МТ головного мозга половозрелых крыс имели изменения, как со стороны нейронов, так и со стороны глии. Наиболее ранимым можно считать кортико-медиальный отдел, так как статистически значимых изменений показателей клеток в нем установлено больше.
Морфологические изменения ядер основных отделов МТ головного мозга половозрелых крыс были ярко выражены в экспериментальной группе (IIФ2/30) с влиянием фенобарбитона в большой дозе (70 мг/кг) в течение 30 суток. На ряду с сохранившимися у части нервных клеток признаками морфологических изменений (тотальный хроматолиз, набухание, наличие
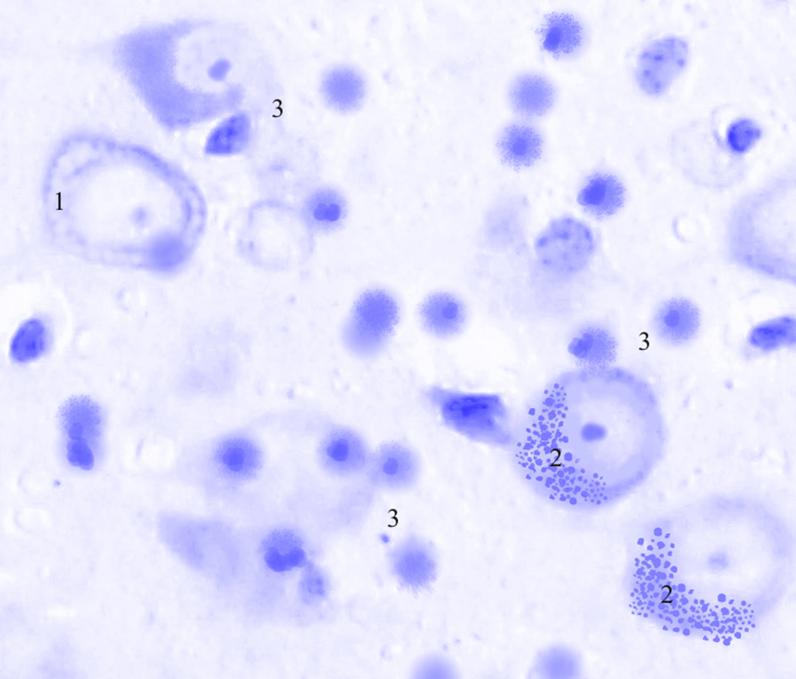
Рис. 3.39. Микрофотография. Центральное ядро кортико-медиального отдела миндалевидного тела головного мозга половозрелой крысы на 15 сутки эксперимента с фенобарбитоном в дозе 70 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Хроматолиз.
2. Агрегация глыбок тигроида.
3. Сателлитоз.
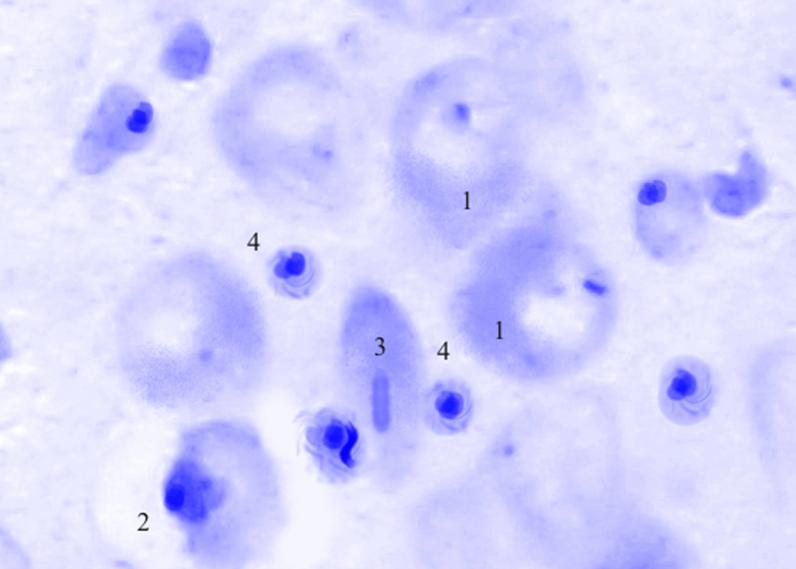
Рис. 3.40. Микрофотография. Базальное ядро базо-латерального отдела миндалевидного тела головного мозга половозрелой крысы на 15 сутки эксперимента с фенобарбитоном в дозе 70 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Гиперхроматоз.
2. Перицеллюлярный отек.
3. Сморщенные клетки.
4. Сателлитоз.
мелких и крупных вакуолей в цитоплазме, увеличение ядра, изменение его формы, эктопия, одновременное наличие гиперхроматоза) появились сморщенные клетки (уменьшенные размеры тел нервных клеток, утрата свойственной округлости, нейроны вытягиваются, суживаются и становятся угловатыми) с изменениями со стороны ядра и вещества Ниссля. Указанные изменения нейронов аналогичны таковым в неполовозрелой группе. Иногда обнаруживались участки активной нейронофагии и клетки-тени. В данных условиях эксперимента глия реагировала усиленной пролиферацией и сателлитозом (рис.3.41-3.42). Так, на 30 сутки в базо-латеральном отделе МТ головного мозга половозрелых крыс выявлялись увеличение площади сечения клеток с 73,54 ± 0,11 до 83,13 ± 0,11 (t = 4,22; Р < 0,05), уменьшение количества нейронов с 1561,24 ± 2,89 до 1166,67 ± 4,42 (t = 3,536; Р < 0,05); происходило также увеличение количества общей глии с 2236,67 ± 3,64 до 3083,33 ± 3,02 (t = 3,092; Р < 0,05), глиального индекса с 1,44 ±0,11 до 2,64 ± 0,14 (t = 2,33; Р < 0,05) и перинейронального индекса с 0,5 ± 0,08 до 0,93 ± 0,09 (t = 2,549; Р < 0,05), то есть указанные изменения имели статистически достоверный характер. По всем остальным морфометрическим показателям изменения не достигали статистической достоверности. В кортико-медиальном отделе МТ наблюдались увеличение площади сечения нейронов с 63,6 ± 0,1 до 82,64 ± 0,11 (t = 3,278; Р < 0,05). Реакция глии выражалась в увеличении количества сателлитной глии с 909,68 ± 3,33 до 1532,25 ± 5,59 (t = 2,524; Р < 0,05). Как видно, по всем выше приведенным показателям изменения носили статистически достоверный характер. Другие показатели статистически недостоверны. Следовательно, на 30 сутки статистически достоверные изменения, как со стороны нейронов, так и со стороны глии отмечались в обоих отделах МТ головного мозга половозрелых крыс, однако, наиболее ранимыми, в данный срок эксперимента, можно считать ядра базо-латерального отдела, так как в них наблюдалось больше статистических значимых изменений изучаемых параметров нейронов и глии.
При увеличении сроков эксперимента до 60 суток, также как и в
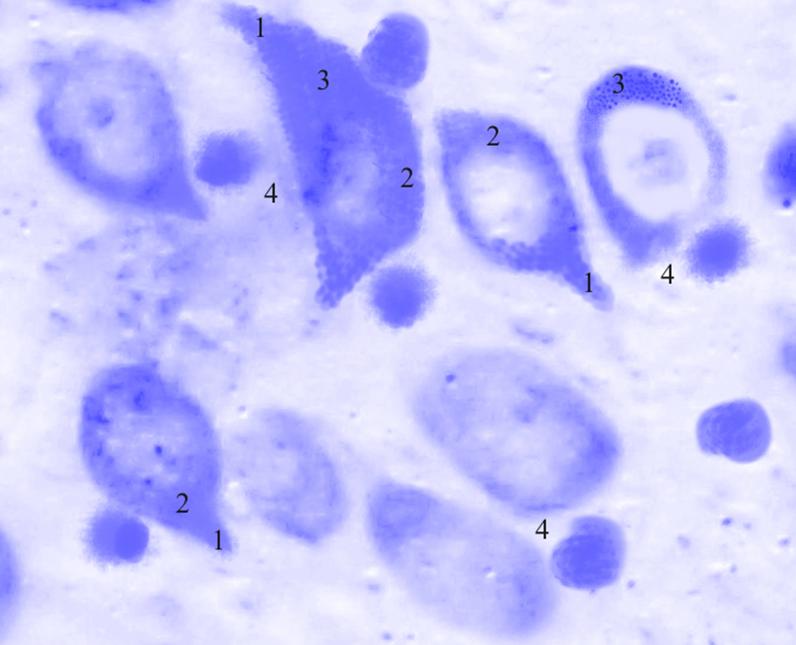
Рис. 3.41. Микрофотография. Кортикальное ядро кортико-медиального отдела миндалевидного тела головного мозга половозрелой крысы на 30 сутки эксперимента с фенобарбитоном в дозе 70 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Утолщение отростков.
2. Гиперхроматоз.
3. Агрегация глыбок тигроида.
4. Сателлитоз.
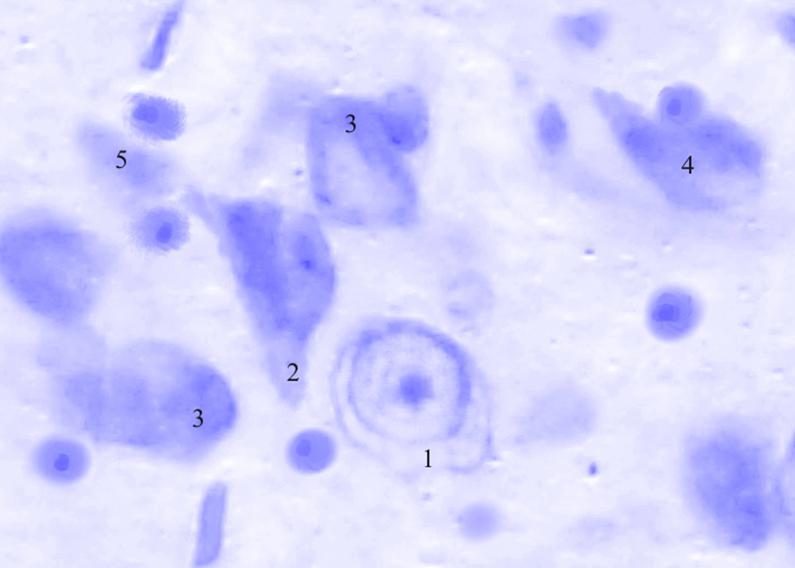
Рис. 3.42. Микрофотография. Базальное ядро базо-латерального отдела миндалевидного тела головного мозга половозрелой крысы на 30 сутки эксперимента с фенобарбитоном в дозе 70 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Хроматолиз.
2. Утолщение отростков.
3. Гиперхроматоз.
4. Сморщенные клетки.
5. Клетки-тени.
экспериментальной группе с интоксикацией фенобарбитоном в меньшей дозе (30 мг/кг), мы наблюдали признаки репаративного процесса, выражавшиеся в основном глиальной реакцией (пролиферация нейроглии, ее активация и усиленный сателлитоз) (рис. 3.43). Так, на 60 сутки в базо-латеральном отделе МТ наблюдалось увеличение площади сечения нейронов с 72,9 ± 0,08 до 87,8 ± 0,06 (t = 4,235; Р < 0,05) со стороны параметров, характеризующих нервные клетки, а со стороны параметров, характеризующих реакцию глии – увеличение количества общей глии с 1894,92 ± 3,85 до 2636,36 ± 1,73 (t = 4,483; Р < 0,05) и сателлитной глии с 687,6 ± 3,09 до 1455,55 ± 1,34 (t = 5,483; Р < 0,05). По другим параметрам изменения не имели статистической достоверности. В кортико-медиальном отделе МТ выявлено увеличение площади сечения ядер нейронов с 3,95 ± 0,02 до 4,67 ± 0,01 (t = 3,304; Р < 0,05), а также количества общей глии с 1658,99 ± 2,63 до 2318,18 ± 1,55 (t = 4,78; Р < 0,05) и сателлитной глии с 668,2 ± 2,12 до 927,3 ± 1,68 (t = 3,891; Р < 0,05). По этим параметрам изменения имели статистическую достоверность (по остальным параметрам изменения не являлись статистически достоверными). Реакция клеток, описанная при микроскопии, подтверждалась морфометрически и была более выражена со стороны нейронов в базо-латеральном отделе МТ. В тоже время в кортико-медиальном отделе наряду с существенными изменениями нейронов оставались выраженными глиальные изменения.
Таким образом, указанные данные могут свидетельствовать, что воздействие фенобарбитона в дозе 70 мг/кг вызывает у половозрелых крыс вначале (7 суток) неспецифические полиморфные морфологические изменения во всех ядрах обоих отделов МТ головного мозга, а с увеличением срока эксперимента эти изменения могут приобретать и дистрофический характер. Данные морфометрии показывают, что достоверно изменяются показатели нейронов и глии ядер обоих отделов МТ головного мозга крыс в той или иной степени. При этом реакция и нейронов, и глии на воздействие фенобарбитона выражена.
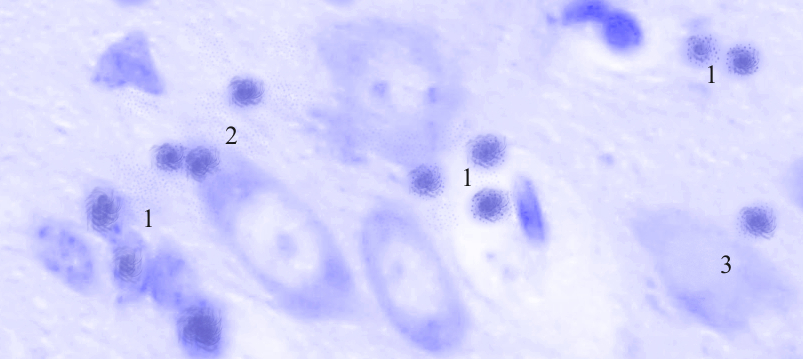
Рис. 3.43. Микрофотография. Медиальное ядро кортико-медиального отдела миндалевидного тела головного мозга крысы половозрелого возраста на 60 сутки эксперимента с фенобарбитоном в дозе 70 мг/кг. Окраска по Нисслю в модификации Викторова. Увеличение 1200.
1. Пролиферация глии.
2. Сателлитоз.
3. Клетки-тени.
Для более глубокого анализа обнаруженных в ходе эксперимента морфометрических преобразований МТ был проведен корреляционный анализ. С его помощью было выяснено, как влияет воздействие большой дозы фенобарбитона (70 мг/кг) на взаимосвязь показателей, характеризующих состояние ядер основных отделов МТ головного мозга половозрелых крыс. Анализ включал 2 отдела МТ и 9 показателей клеток ядер указанной структуры головного мозга.
На 7 сутки эксперимента сильная взаимосвязь (0,7 < r < 1,0) наблюдалась между исследуемыми отделами МТ по параметрам площади сечения тела нейронов, плотности нейронов, перинейронального индекса и глиального индекса. Средней силы взаимосвязи (0,3 < r < 0,7) обнаруживались по показателям плотности общей глии и интерглиального коэффициента. Слабая взаимосвязь (r < 0,3) обнаруживалась по остальным изучаемым показателям.
На 15 сутки эксперимента сильных взаимосвязей между параметрами исследуемых отделов МТ головного мозга крыс мы не выявили. Связи средней силы (0,3 < r < 0,7) отмечались по параметрам плотности общей и сателлитной глии, глиального индекса, перинейронального индекса, интерглиального коэффициента. По остальным показателям установлена слабая взаимосвязь (r < 0,3).
На 30 сутки эксперимента также сильных взаимосвязей между исследуемыми отделами МТ установлено не было. Средней силы связи (0,3 < r < 0,7) были выявлены по показателям плотности нейронов и ядерно-цитоплазматического отношения. По остальным показателям была выявлена слабая взаимосвязь (r < 0,3).
На 60 сутки эксперимента сильных взаимосвязей (0,7 < r < 1,0) между основными отделами МТ головного мозга половозрелых крыс установлено не было. Связи средней силы (0,3 < r < 0,7) были зафиксированы по показателям плотности нейронов, ядерно-цитоплазматического отношения и сателлитной глии. По остальным параметрам установлена слабая взаимосвязь (r < 0,3).
Сравнивая корреляционные связи контрольных и экспериментальных групп, необходимо отметить, что происходило изменение силы взаимосвязей, как по количеству, так и по качеству. При сопоставлении и сравнении коэффициентов корреляции по срокам эксперимента наибольшее число сильных связей наблюдалось при длительности эксперимента 7 суток, связей средней силы при длительности 15 суток, а другие сроки эксперимента изобиловали слабыми связями.
Таким образом, изучение корреляционных связей показало следующее:
1. характер и сила взаимосвязей между ядрами отделов МТ головного мозга половозрелых крыс существенно изменялись в ходе эксперимента под воздействием большой дозы фенобарбитона – 70 мг/кг;
2. по сравнению с контрольными группами во все сроки эксперимента изменяется характер и сила взаимосвязей между отделами в сторону ослабления в указанных экспериментальных группах;
3. сильные взаимосвязи между отделами МТ головного мозга отмечались в основном при длительности эксперимента 7 суток;
4. средних связей было обнаружено наибольшее количество на 15 сутки данного эксперимента;
5. преобладающими были взаимосвязи между отделами МТ слабой силы, особенно в средние и большие сроки эксперимента (30 и 60 суток).
Еще по теме 3.2.2.Половозрелые крысы.:
- 3.2.1. Неполовозрелые крысы.
- Операция вживления силиконового катетера в яремную вену крысы
- Сравнительное изучение времени жизни в плазме крови крысы пептидов TGENHR и TGeNHR-NH2
- Методика острого эксперимента по введению дисперсии фуллерена Сбо в изолированную петлю тонкой кишки крысы
- Глава 3. Исследование уровня окислительной модификации белков и молекул средней массы на модельной биологической системе желточных липопротеидов, продуктах пчеловодства каквеществах природного происхождения, обладающих антиоксидантным действием, и сыворотке крови экспериментальных животных (крысы)
- Глава 4. Разработка подходов комплексного использования модельной биологической системы желточных липопротеидов при одновременном добавлении продуктов пчеловодства как веществ природного происхождения, обладающих антиоксидантным действием, и сыворотки крови экспериментальных животных (крысы) при спонтанном и Бе2+-индуцированном окислении
- ГЕЛЬМИНТОЗЫ.
- Экспериментальная часть
- Изучение биораспределения фуллерена Сбо по органам и тканям в токсикологических экспериментах
- Результаты морфологического исследования через 3 месяца
- Блохи и эндемический сыпной тиф.
- ДЕРАТИЗАЦИЯ В ПОРТАХ
- Описание экспериментальных групп
- Wuchereria bancrofti Заболевание: вухерерироз.
- Trichinella spiralis Заболевание: трихинеллез.