Условия культивирования продуцентов тилозина
Динамика накопления в культуральной жидкости антибиотика тилозина имеет типичный вид зависимости, характерный для биосинтеза вторичных метаболитов, т.е. максимум образования биомассы предшествует максимуму антибиотикообразования (рис.
4).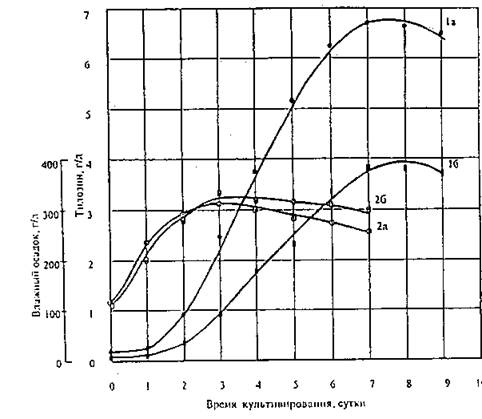
Рис. 4. Динамика накопления тилозина (1) и изменение концентрации влажного осадка (2) в процессе культивирования штаммов Str. fradiae 25 А (а) и Str. fradiae 165 (б) в лабораторных ферментерах объемом 1.7 л
В качестве источника азотного питания для комплексных сред может быть использована рыбная и мясная мука, мясной экстракт, смесь аминокислот, дрожжи, казеин, кукурузный экстракт; в качестве источника углерода — глюкоза, меласса, крахмал. В среде обычно присутствуют в небольших количествах соли, содержащие натрий, магний, алюминий, кальций, кобальт в виде сульфатов, фосфатов и хлоридов. Высокие концентрации ионов железа оказывают угнетающее действие на биосинтез тилозина. Оптимальная концентрация железа в среде 5-10 мкг/мл.
Для культивирования продуцента тилозина обычно используется ферментационная среда следующего состава; рыбная мука —2,0; меласса (по РВ) — 2,0; крахмал — 1,5; (NH4)2HPO4 — 0,02; NaCl — 0,2; MgSO4 • 7Н2О — 0,2; СаСО3 — 0,5; соевое масло — 3,0; pH среды до стерилизации 7,0-7,5.
Отмечается, что существенное влияние на биосинтез тилозина оказывает pH среды, его оптимальное значение, согласно данным литературы, составляет 6,5-7,0 или 7,4-7,6. К концу ферментации величина pH достигает для штамма Str. fradiae 25А — 7,5-7,8, для менее активных штаммов 8,0-8,3. Защелачивание культуральной жидкости, как правило, сопровождается лизисом культуры и падением концентрации антибиотика. Увеличение количества масла в среде, например, путем его дробного внесения и поддержания на заданном уровне, приводит к стабилизации уровня накопления тилозина, а также максимального уровня накопления биомассы и значения pH среды.
Культура лучше растет при температуре 25—32 °С, а синтезирует — при 26—30 °С. Для штамма Str. fradiae 165 показано, что увеличение температуры культивирования до 35 °С сопровождается увеличением количества образовавшейся биомассы и резким снижением антибиотической активности. В основном, на протяжении всего процесса поддерживают температуру 28—30 °С, но в литературе есть данные о преимуществе ферментаций, во время которых в первые 10 часов ферментации температуру увеличивали до 37 °С.
Процесс культивирования осуществляют при постоянном перемешивании, расходе воздуха 1 об./об. • мин. Синтез тилозина заканчивается на 6—9-е сутки.
Рекомендуется проводить процесс ферментации в колбах объемом 300 и 750 мл на качалках с числом оборотов 220-300 об./мин и в аппаратах объемом 1,7 л, 630 л, 63 м3 с объемами среды 1,
1 л, 300 л, 30 м3 соответственно, при следующих условиях: доза засева, как правило, 2,5-10 %, температура культивирования — 28 °С, расход воздуха — 0,5-1,5 об./об. • мин, число оборотов мешалки — 550-650 об./мин для аппаратов объемом 1,7 л и 150-170 об./мин для ферментеров объемом 630 л и 63 м3.
Для достижения высокого выхода тилозина необходимо обеспечить в ферментере высокую концентрацию растворенного кислорода, максимальное потребление кислорода наблюдается в начале ферментации. Есть данные, что динамика изменения концентрации растворенного кислорода в культуральной жидкости продуцента тилозина существенно зависит от изменения ее реологических свойств.
Как показано в работе, максимальная скорость синтеза тилозина, достигаемая на 3-4-е сутки ферментации при периодическом культивировании продуцентов, существенно зависит от интенсивности аэрации культуральной жидкости и для штаммов Str. fradiae 25А и Str. fradiae БС-1 может быть доведена до 1,5-2,0 г/л ■ сутки. Оценка интенсивности аэрации культуральной жидкости в ферментере может вестись по,количеству СО2 в отводимом из ферментера воздухе.
При этом для эффективного биосинтеза тилозина концентрация СО2 в отводимом воздухе к моменту максимальной скорости роста биомассы (1-2-е сутки ферментации) должна быть порядка или более 1 г СО2/л • час. Из данных рис. 5 следует, что максимальная интенсивность поглощения кислорода растущей культурой штамма-продуцента тилозина Str. fradiae БС-1, определяющая при прочих равных условиях максимальную скорость биосинтеза тилозина во время ферментации, наблюдается в первые двое суток культивирования штамма в период интенсивного роста биомассы. В дальнейшем интенсивность дыхания резко уменьшается до 20-30 % от максимального значения и сохраняется примерно на этом уровне до окончания периода интенсивного синтеза тилозина.Важно, однако, отметить, что при увеличении интенсивности аэрации культуральной жидкости концентрация тилозина в конце ферментации и максимальная скорость его синтеза стремятся к некоторому пределу, характерному для данного штамма-продуцента и выбранного состава питательной среды. В рассматриваемом случае предельное значение концентрации тилозина в конце ферментации равно 8 г/л, а максимальное значение скорости синтеза тилозина 2,0 г/л • сутки (рис. 6).
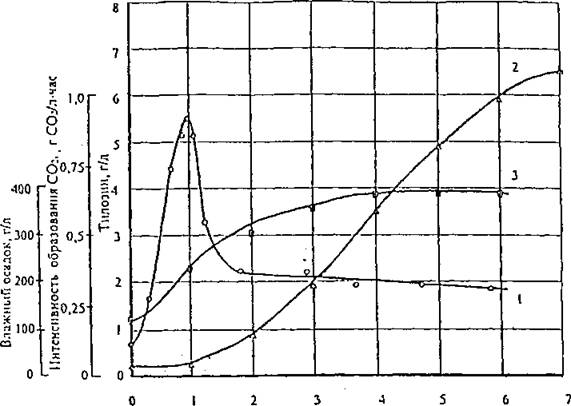
Рис. 5. Зависимость от времени интенсивности образования СО2 (I), изменения концентрации тилозина (2) и влажного осадка (3) в культуральной жидкости в процессе культивирования штамма Str. fradiae БС-1 в лабораторных
ферментерах объемом 1,7 л. Расход воздуха 1,0 об/об • мин, число оборотов мешалки < 600 об/мин
С целью интенсификации процесса биосинтеза тилозина и других антибиотиков предложено разбавление культуральной жидкости в процессе ферментации водой.
Имеются указания на то, что выход тилозина может быть увеличен при добавлении в среду фосфорнокислого магния, пропилового спирта, фумаровой кислоты, бромида аммония, конденсатов формальдегида. Показано, что влияние циклического АМФ на процесс биосинтеза тилозина зависит от состава среды и физиологического состояния культуры.
На примере тилозина в условиях хемостата отчетливо показан эффект репрессирования синтеза антибиотика глюкозой.
Синтез тилозина быстро прекращается после добавления в культуральную жидкость на 120 ч ферментации глюкозы, причем это продолжается до тех пор, пока добавленная глюкоза не усваивается полностью. Остается открытым вопрос, обусловлено ли действие глюкозы катаболитной репрессией или ингибированием. Предложено в составе ферментационной среды глюкозу заменять глицерином, что обеспечивает в некоторых случаях увеличение скорости синтеза тилозина на 20 %. Это связано с тем, что при стационарных условиях глицерин вызывает меньшую репрессию синтеза тилозина, чем глюкоза.В некоторых работах указывается, что иногда азотная репрессия у штамма Str. fradiae не сходна с углеродной репрессией. Например, значение скорости синтеза тилозина возрастает с увеличением в среде концентрации глутамата натрия. Увеличение удельной скорости поглощения глутамата натрия может стимулировать синтез тилозина путем увеличения количества предшественников, необходимых для синтеза лактонового кольца; На основании полученных данных была разработана среда с низким содержанием глюкозы и повышенным содержанием глутамата натрия. Зависимость начала синтеза тилозина от перехода культуры в идиофазу на этой среде отсутствует, антибиотик образуется в момент быстрого роста культуры.
Проводились работы по созданию отечественных продуцентов тилозина, в частности, Str. fradiae 25В (ВКПМ-S 1112), устойчивых к катаболитной репрессии и ингибированию неорганическим фосфором.
Кроме тилозина штамм Str. fradiae способен образовывать тилозиноподобные вещества (макро- цин, десмикозин, реломицин, лактеномицин). Предложены методики определения состава этого антибиотического комплекса. Имеются сведения о том, что макроцин является непосредственным предшественником тилозина, лактеномицин и десмикозин могут образовываться на различных стадиях превращения тилактона. Реломицин, появляющийся в конце процесса ферментации, является продуктом восстановления тилозина.
Подробно изучены генетические и биохимические аспекты механизма образования тилозина и родственных ему соединений, принимающих участие в этом процессе.
Предложена схема ко-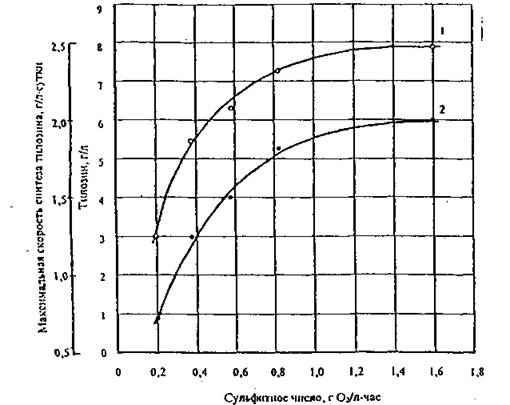
.Рис. в. Зависимость от конечной концентрации (1) и максимальной скорости синтеа (2) тилозина в культуральной жидкости от интенсивности аэрации при культивировании штамма Str. fradiae БС-1 в колбах объемом 300 мл. Продолжительность ферментации — 7 суток
нечного этапа биосинтеза тилозина. Проведены работы по установлению локализации генов, ответственных за процесс биосинтеза тилозина. Авторами работы выделен соответствующий фрагмент ДНК.
Изучено влияние ионов аммония, неорганического фосфата и аминокислот на процесс биосинтеза тилозина. При увеличении уровня фосфата наблюдается уменьшение скорости синтеза тилозина.
Увеличение степени поглощения фосфора уменьшает количество образовавшегося тилозина и увеличивает количество реломицина. В процессе периодического культивирования наблюдается трехкомпонентная система (тилозин, реломицин и макроцин), а в ферментациях, проведенных в хемостате, — двухкомпонентная (тилозин и реломицин).
Изучен процесс биосинтеза тилозина в условиях культивирования на синтетической среде с подпиткой. С целью поддержания скорости синтеза тилозина на высоком уровне были использованы три скорости долива среды. При оптимальной скорости подпитки общий объем синтезируемого антибиотика превысил контрольный, что происходило не за счет увеличения удельной скорости биосинтеза, а за счет дополнительного (по сравнению с контрольным) прироста биомассы.
С помощью циклических подпитывающих добавок возможна стимуляция активности ферментов, участвующих в биосинтезе тилозина, и увеличение периода активного синтеза этого антибиотика на синтетической среде.
Подобран количественный состав основных питательных компонентов комплексной ферментационной среды, вносимых в начале и по ходу процесса культивирования продуцентов тилозина, исключающий снижение концентрации антибиотика к концу ферментации и позволяющий, при необходимости, продлить период активного синтеза до 10—12 суток.
Скорость синтеза тилозина при продлении срока ферементации постепенно уменьшается, однако максимальная концентрация антибиотика в культуральной жидкости в конце ферементации возрастает. Целесообразность такого продления ферментации зависит от складывающегося на производстве соотношения между различными составляющими себестоимости продукта.Для продуцента тилозина Str. fradiae показано, что ряд аминокислот в свободном или связанном (в пептидах и белках) виде может служить альтернативным источником ацетил-КоА. Так, например, L-треонин служит источником коферментов, необходимых для биосинтеза тилозина и жирных кислот. Валин также является источником предшественников жирных кислот для биосинтеза макролидов, мощным индуктором синтеза валиндегидрогеназы в мицелии Str.fradiae.
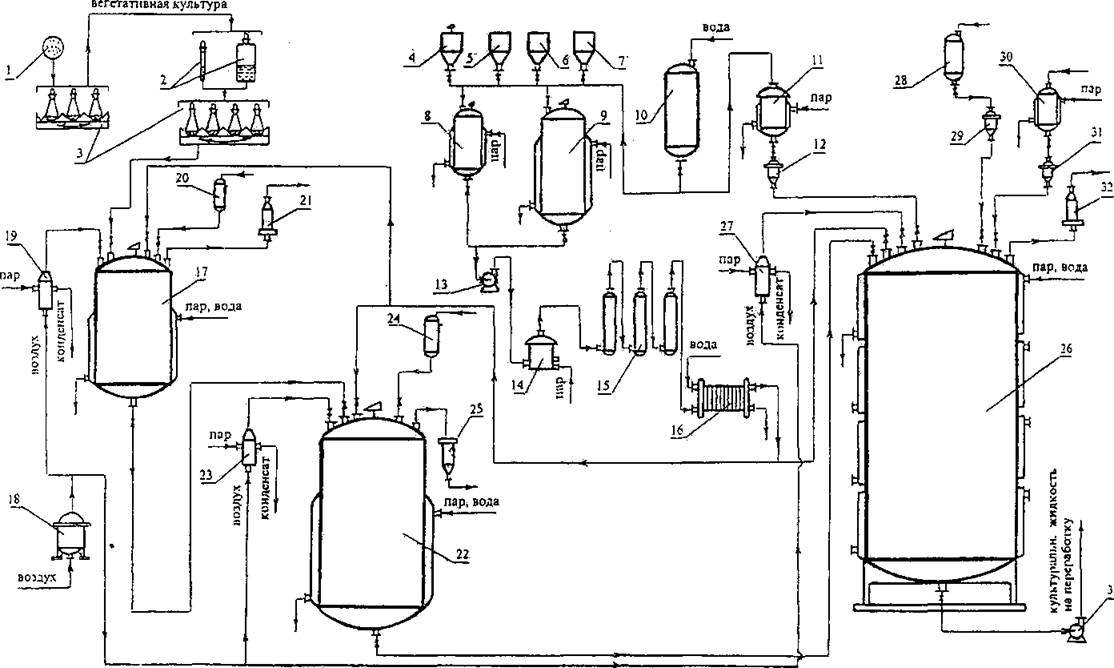
Рис. 7. Принципиальная технологическая схема производства тилозина для стадий подготовки посевного материала и культивирования штаммов-продуцентов Str. fradiae. 1 — чашки Петри с колониями штамма-продуцента, 2 — пробирки и матрасы с вегетативной культурой, 3 — качалка с колбами для проверки активности колоний и размножения посевного материала, 4,5,6,7 — сборники компонентов питательной среды и титрующих растворов, 8,9 — смесители для приготовления сред,
10,11 — сборник и стерилизатор воды, 12, 29, 31 — мерники, 13, 33 — насосы, 14 — нагреватель-стерилизатор, 15 — выдерживателъ, 16 — теплообменник-охладитель, 17 — инокулятор, 18 — головной фильтр, 19, 23, 27 — индивидуальные фильтры очистки воздуха, 21, 25, 32 — фильтры очистки отработавшего воздуха,
20, 24, 28 — сборники пеногасителя, 22 — посевной аппарат, 26 — ферментер. 30 — стерилизатор масла
6.
Еще по теме Условия культивирования продуцентов тилозина:
- Технологическая схема стадий подготовки посевного материала и культивирования продуцента тилозина
- Влияние условий культивирования продуцентов на биосинтез аргинина
- Продуценты тилозина
- Хранение продуцентов тилозина и проведение поддерживающей селекции штаммов
- Приготовление и хранение посевного материала продуцентов тилозина
- Основы технологии культивирования микроорганизмов-продуцентов микробных липидов
- Условия культивирования липидообразующих дрожжей
- 4.1. Взаимодействие ЭК - ГМК в условиях совместного культивирования
- Питательные потребности грибов рода Malassezia и условия для их культивирования
- Влияние условий культивирования на активность ключевых ферментов биосинтеза аргинина
- 2. Товарные формы тилозина
- 1. Применение, механизм действия тилозина
- Ветеринарный антибиотик тилозин
- Физико-химические свойства, методы определения концентрации тилозина
- Исследование ростовых свойств продуцентов гомелина, лепидоцида и битоксибациллина
- Биосинтез аргинина мутантными штаммами-продуцентами Brevibacterium flavum