Глава 2. Объекты и методы исследований
Микрооорганизмы и среды выращивания
В работе использовали Micrococcus luteus NCIMB 33267, Escherichia colt HMS174(DE3),
Mycobacterium tuberculosis H37Rv, Компетентные клетки E.coli готовили по стандартной методике
(Jansscn & Irvin, 2001, Molecular cloning).
Для выделения плазмидной ДНК использовали наборPromega Wizard Plus SVMinipreps (Promega). Для выделения РНК М luteus использовался набор
фирмы Qiagen RNeasy Mini Kit (Qiagen). Для выращивания E.coli и M.lufeus использовали богатую
среду BroThE (LabM),. M. luteus выращивали также на бедной, минимальной среде, содержащей
лактат и аммоний (ЛММ) (Mukamolova et al., 1998). Культуру М. tuberculosis выращивали на
жидкой среде Мидельбрука (Yeremeev et al,, 2000). Все культуры выращивали при 37 °С ( кроме
MJuteus- при 30 °С) в условиях аэрации.
ОТ-ПЦР(обратно-транскрипционнъш ПЦР, RT-PCR)
Для реакции обратной транскрипции использовали кит AMV Reverse Transcriptase (Promega), все
процедуры проводили в строгом соответствии с инструкциями, прилагаемыми к набору,
Праймеры для обратной транскрипции и амплификации кДНК (20-30 пМ на пробу):
Rv0367(F)-5'-CCAACGTCAGCGTCGCCAAC-3'
Rvl009 (F>- З'-ССОТСССССТОТТОСААА-З1
Rvl384 (F) - 5'-CGGTTGTGCATCCTTTGCC-3'
Rv2389 (F) - S'-TATGACACCGGGTTTGCTTACO'
Rv2450 (F) - 5'-CTCATCGCCGCCGCGATTO-3*
UNI (R) - 5 '-CGGTGTTGACCGACCC AGTT-3'
Rpf-(R) 5'-TTCATGTCCCAGGTGCCGTT-3' (F) 5'-GAGGACTCGCCATGGACACC-3'
Амплификация: 48С — 45мин.
32 94С-2мин,
94С-30се .
52C-30cej? 30 раз.
72С-1мит.
72С-7мин,
Выделение рекомбипантного белка Rpf-HisTag Micrococcus luteus (и его аналогов из
MJuberculosis) из штамма E*coli HMS174(DE3)t содержащего плазмиду Rp.-HisTag/pET19b
Трансформированные клетки выращивали в среде Broth E в присутствии ампициллина (100 мкг в мл) до OD60D = 0,7-0,9.
Затем к культуре добавляли изопропил JS-D-тиогалактопиранозид до конечной концентрации 0,5 мМ и инкубировали культуру 3 часа при 37 °С. Клетки разрушали ультразвуком на ледяной бане 10 раз по 15 секунд.* ■ 4
Хроматография на Ni2*chelatian Sepharose 6В column
Все этапы проводили при +4 С. Полученный лизат клеток ЕхоН центрифугировали на холоду при 13000g 25 минут, супернатант разводили в 4 раза буфером для нанесения (5 тМ имидазола, 0.5 М NaCl, 20 тМ Tris-HCl рН 8,0, б М мочевины) и наносили на аффинную колонку Iminodiacetic acid immobilized on Sepharose (Sigma), предварительно обработанной NiSOj. После нанесения супернатанта колонку промывали 20. объемами буфера с 50 мМ имвдазолом. Затем колонку промывали 5 объемами буфера, содержащего 100 мМ имидазола и элюировалй 3-мя oбъeмa^fи буфера, содержащего 500 мМ имидазола. Диализовали белки против 2-5 литров 5 мМ Tris-HCl рН 7,4,20 часов при 4° С.
Оценку концентрации рекомбинантных белков проводили по измерению поглощения Адяони У диализованных образцов против буфера, в котором происходил диализ. Рсчетные коэффициенты экстинщш: 1 сои Агво Rpf M. luleus = ОД мг/мл белка
33 1 eon A2S0 Rpftl ~ 0,56 мг/мл белка
1 eon Агяо Rpft2 = 0,43 мг/мл белка
1 eon А2Д0 Rpft3 = 0,31 мг/мл белка
1 eon A2soRpft4 = 1,13 мг/мл белка
1 eon A2S0 Rpfti - 0,91 мг/мл белка
Хроматография белков на DEAE-Sepharose
Использовали DEAE-Sepharose Fast Flow DFF-lOO(Sigma). Последовательно промывали колонку водой, затем 10 мМ трис-HCl рН 7.4. После нанесения образца промывали колонку 10 мМ тркс-НС1 рН 7.4 + 100 мМ NaCI. Элюировали Змл 10 мМ трис-НС1,рН 7,4 + 330 мМ NaCl-
SDS-электрофорез белков в иолиакрилам идиом геле (по Лэммли)
- Электрофорез проводили в приборе BioRad (в гелях содержалось 12% акриламида) на холоду при 30 тА на 1 пластину. 5хЭлеюродный буфер (0,25 М трис, 2 М Glycine, 0,5 М SDS рН 8S8). 8хБуфер для разделяющего геля (3 М трис-HCl рН 8,8 и при 25 С). 4хБуфер для концентрирующего геля (0,5 М трис-HCl рН 6,8 при 25°С):
Градиентным не денатурирующий электрофорез белков в полиакриламидном геле
Данный вид электрофореза проводили по методике J.M.Walker, Gradient SDS Polyacrylamide Gel Electrophoresis of Proteins, (Basic Protein and Peptide Protocols, Methods in Molecular Biology, v.32, ed.
J.M.Walker, Humana Press Inc., 1994 ). Градиентный гель содержал от 5 до 30 % акриламида.Приготовление Коллоидного Кумасси G-2S0:
Данный метод окраски позволяет детектировать от 0,2 мкг белка в полосе).
34
Растворы для этой окраски содерожали: V/
Г
1. 5% раствор Кумасси G-250
2. 2. 0,29 М Н3Р04 {2,85% р-р)
3. 25% раствор (NH4)2S04в °>29 м растворе Н3Р04
4. Раствор Кумасси G-250 в НЭР04:
2 мл 5% раствора Кумасси G-250 добавляли к 100 мл р-ра 15%(NH4)2S04 в 0,29 М растворе Н3РО4*
5. Непосредственно перед окрашиванием геля добавляли 10 мл метанола к 40 мл перемешанного раствора Кумасси G-250 в Н3РО4 (см. п.4).
Окрашивание производили при комнатной температуре в течение 20 часов.
Иммуноблоттинг рекомбинантных белков
Электрофорез белков проводили в двух одинаковых пластинах 12% SDS ПААГ, одну из которых затем окрашивали коллоидным Кумасси G-250, а вторую использовали для собственно электроблоттинга в приборе BIORAD в буфере ТГЭС (трис-3,2 г/л, глицин-14,4 г/л, SDS - 1 г/л, этанол - 200 мл, диет, воды до 1 л). Блоттинг проходил в течение 16 часов при 50 мА при 4°С на мембрану Millipore Nitrocellulose HAHY с размером пор 0,45 мкм,
Неспецифическое связывание с мембраной предотвращали проводя инкубацию мембраны в буфере TBS+3%BSA при 37°С в течение часа при перемешивании (50 rpm), после чего мембрану промывали 3 раза по 10 мл TBS при 37°С Буфер TBS: 20 мМ трис-HCI рН 8.0, 150 mM NaCI (20 ml 1 М трис-HCI рН 8.0; 8.7 g NaCI; Н^0 до 1 л); BSA - Бычий сывороточный альбумин.
Первичные AT (поликлональные кроличьи AT против Rpf:100 мкл препарата AT "A3" в 8 мл TBS) инкубировали с мембраной в течение 1-2 часов при 37°С при перемешивании (50 грт), после чего мембрану промывали 3 раза по 5 мин, 10 мл TBS+Tween 80 (0,05%) при 37ПС при перемешивании (50 грт).
35 Вторичные AT (AntiRabbit AT conjugate with Alcalic Phosphatase (Sigma)) использовали в
разведении 1:5000 {2 мкл AntiRabbit AT conjugate / 10 мл TBS+Tween 80 (0,05%)); AT
инкубировали в течение 40 мин.
при 37°С при перемешивании.Мембрану промывали 3 раза по 5 мин. 10 мл TBS+Tween 80 (0,05%) при 37°С, затем
аналогично - TBS, после чего окрашивали 3-5 мин SIGMA FAST BCIP/NBT Alcalinc Phosphatase
Substrate (Sigma) при комнатной температуре.
Анализ степени полимеризации рекомбшшнтиых белков при помощи гпутаралъдегидных
сшивок
Для сшивки готовили пробы объемом 100 мкл. Конечная концентрация белка в пробах составляла 40 мкг/мл. Использовали два варианта метода: 1) Концентрация глутарового альдегида остается неизменной, а время экспозиции его с белком меняется; 2) Концентрация глутарового альдегида в пробах изменяется, а время экспозиции постоянно.
В обоих случаях реакцию останавливали при помощи 10-и кратного избытка боргидрида натрия, инкубация с которым протекала 20 мин. Избыток боргидрида удаляли при помощи 2М NaH2P04, концентрация которого доводится до 7?5 объемных %. В варианте 1 во всех пробах концентрация глутарового альдегида составляла 1%. Экспозиция с белком до остановки
реакции составляет: 30с; 1 мин; 2 мин; 3 мин; 4 мин.
В варианте 2 Концентрация глутарового альдегида в пробах составляет: 0,1 %; 0,25 %; 0,5 %; 1%; 2 %. Во всех пробах экспозиция с белком составляет 2 мин.
В обоих опытах неспецифические связи диссоциировали, добавляя SDS до конечной концентрации 1 %. Далее пробы диализировали в течение суток против 0,2 % SDS при температуре 40-60 °С. После чего пробы подвергали SDS-форезу по обычной методике для установления молекулярной массы полученных агрегатов.
36 Животные
В качестве подопытных животных были использованы мыши обоего пола линии C57BL/6JCit (B6)
из питомника ЦНИИ Туберкулеза РАМН, массой 22-24 г.
Пролиферативный тест
Для постановки пролиферативного теста мышей иммунизировали подкожно в подушечку задней
лапы 10 мкг белка в неполном адъюванте Фрейнда (НАФ) и через 2 недели исследовали in vitro
*
пролиферацию клеток подколенных лимфоузлов в присутствии белков семейства Rpf, Суспензию клеток лимфоузлов (2x10 клеток/лунку) культивировали в 96-луночном планшете (фирма Costar, США) в присутствии 10 мкг/мл Rpf-белков при 370(^в атмосфере, содержащей 5% СОз в течение 72 ч.
Уровень пролиферативной активности лимфоцитов оценивали по включению 3Н-тимидина, Для этого за 18 ч до окончания инкубации в лунки добавляли но 0,5 uCi 3Н-тимидина. По окончании инкубации клетки с помощью харвестера переносились на фильтры и с помощью сщштиляционного счетчика определяли уровень включения радиоактивной метки в ДНК.Иммунизация Мышей иммунизировали 10 мкг белка в НАФ (неполный адъювант Фройнда), разведенном в 2 раза физиологическим раствором, под кожу спины 3-кратно с 2-недельным интервалом. Через 10 суток после последней инъекции у подопытных животных из ретроорбитального синуса брали кровь для определения в сыворотке количества специфических антител класса TgG с помощь иммуноферментного теста (ELISA).
Определение количества антител (ELISA)
Количество антител к белкам Rpf в сыворотках животных определяли при помощи иммуноферментного анализа. Использовали планшеты фирмы Costar, в течение ночи в каждой лунке инкубировали 10 мкг соответствующего антигена в 0,1 М карбонатном буфере рН 9,6. Неспецифическое связывание предотвращали, инкубируя планшет с 1 % БСА в 0,05 М ТРИС-НС Рв
37
течение 0,5 ч при 37 °С. После отмывки проводили инкубацию с сыворотками, содержащими
антитела против соответствующих антигенов в течение 2 ч при 37 °С. После отмывки планшеты инкубировали в течение 2 ч при 37 СС с Alkaline phosphatase-conjugated rat anti-mouse TgGl
т f ^
monoclonal antibodies (Pharmingen, San-Diego, Ca). Окрашивание проявляли в соответствии с рекомендациями фирмы Pharmingen,
Количество Rpf в супернатантах M.luteus определяли методом прямого ИФА но стандартной схеме, Суисрнатанты, содержащие Rpf .инкубировали в течение ночи в планшете фирмы Costar. Неспецифическое связывание предотвращали инкубируя планшет с 1% БСА в 0705 М ТРИС-HCI в течение 0,5 ч при 37 °С. После отмывки проводили инкубацию с кроличьей сывороткой, содержащей поликлональные антитела против Rpf в разведении 1:20000,. После отмывки в качестве вторичных антител использовали AntiRabbit AT conjugate with Alcalic Phosphatase (Sigma) использовали в разведении 1:5000.
Окрашивание проявляли при помощи SIGMA FAST pNPP Alcaline Thosphatase Substrate (Sigma). Количество Rpf определяли при помощи калибровки, построенной на основании измерения известных количеств рекомбинантного Rpf, растворенного в среде культивированиям iuteus.Заражение мышей
Через б над, после последней иммунизации мышей внутривенно заражали М. tuberculosis H37Rv в летальной дозе 2х107КОЕ (Yeremeev ct al.t 2003).
Протективный эффект Rpf белков оценивали по количеству жизнеспособных колониеобразующих микобактсрий на среде Мидельбрука (число КОЕ - колоний образующих единиц), в легких зараженных мышей, а также по длительности выживания иммунизированных и контрольных мышей.
8.I.L
Гсш|1>-ин5гак?рш^ №i№if№K3#in» тткулт&пш э^р^-ирчшл!. & кшт^шоЙ излишке «оои*; ююоировашзыя is $&эдюрс рМ'11.20 с
7
Рис, 4. ЗОК-гшсктрофорсграмма,, характеризующая выделяемые нами рекомбитшттые белки семейства Rpf, 1- маркеры молекулярное массы. 2 -Rpf 15, 3 -Rpft4,4- Rp£t3. 5- Rpf42, 6 -RpftU7-Rpf,
Все вдделяемые белки были охарактеризованы при помощи SDS-эЯЕктрофорйза в ПААГ ц иммурюблогтаига; для последнего нпюльзошлиеь иоликпо1шлг>ныь' антитела, полученные против Rpf, Степень очистки препаратов рекомбянантны* бегжпй превышала 95% (one пиналась сканированием SDS-mcEaTioiboperpnMOvE).
44 ЗЛ.2. Изучение свойств Rpf Micrococcus luteus
Так как белок Rpf был открыт в нашей лаборатории незадолго до начала работы над данной диссертацией, сведений о его физико-химических свойствах практически не было. Для того, чтобы установить в каком состоянии (мономолекулярном, полимерном) пребывает выделенный рекомбинантный белок и сильно ли он отличается от нативного белка М. luteus из супериатанта бактериальной культуры соответствующие препараты белков были подвергнуты HPLC гель-фильтрации на колонках BloSep 2000 и TSK 2000,
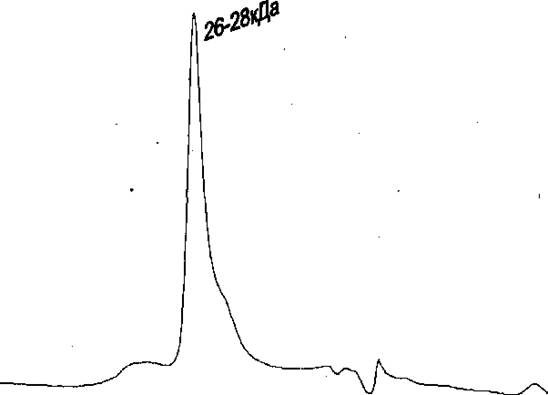
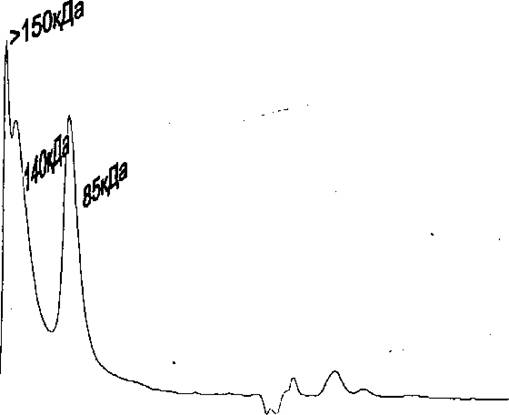
Рекомбинантный Rpf В случае рекомбинаитного белка, основная его масса была представлена в виде мономера (Рис. 5А), по при хранении как при + 4С, так и при - 20С, в течение, короткого времени наблюдалось возникновение крупных агрегатов, часть из которых представлена формами с молекулярной массой 140 и более кДа, а часть агрегатом с массой 85 кДа (Рис. 5В), Л
\UrnV
chl
10 12 14 lit IS 20 22 2-1 2f> 24 30 32 34 36
45
в
 |
г, !47mV
А
| П |
chl 1
| vS |
т---------- г
| с\ Р |
| \1 |
ID 11 12 1J 14 15 16 17 ион
офиль элюции рекомбинантного Rpf сразу после выделения, при HPLC гель-фильтрации на колонке BioSep 2000. В - Профиль элюции рекомбинантного Rpf через 48 часов после выделения {хранился при - 20 °С), при гель-фильтрации на колонке TSK-gel 2000, (указаны мол. массы, соответствующие пикам),
Поведение Rpf rtpu хранении Препарат рекомбинантного Rpf хранили при - 20 °С в 50 % глицерине и при + 4 °С без глицерина. Было обнаружено, что при хранении в нестерильных условиях белок нестабилен во времени и подвергается протео логической деградации- Скорость такой деградации варьировала
для различных выделенных препаратов белка (см. ниже). При хранении белка при -20 ftC он
У \ I
оставался стабильным (согласно SDS-форезу), Филтрация свежеполученного белка через
бактериальный фильтр 0.22 мкм приводила к существенной стабилизации белка при хранении при
+4 СС.
Если стерильный рскомбинантнын белок хранился при любой температуре в течение
нескольких недель, его нельзя было зафиксировать при пропускании через HPLC гель-
46 фильтрационные колонки, но в то же время, он хорошо детектировался SDS-электрофорезом,
правда при этом в препарате, хранившемся при + 4 С выявлялись дополнительные пептиды
(несколько полос на электрофореграмме в районе от 10 до 20 кДа), которые, очевидно»
соответствовали фрагментам белка, что свидетельствовало о том, что часть его подвергалась
деградации (Рис. 11). Этот феномен можно объяснить образованием крупных агрегатов белка,
настолько больших, что они даже не могут пройти через гель-фильтрационную колонку. Эту
гипотезу подтверждает и тот факт, что порядка 15.% белка (по поглощению при к = 280 нм)
задерживается при фильтрации через бактериальные фильтры (размер пор 0.22 (im).
Для того чтобы изучить, что представляют собой агрегаты Rpf, был проведен эксперимент по
ковалептному сшиванию белка глутаровым альдегидом. Глутаровый альдегид, являющийся
бифункциональным реагентом, способен образовывать межмолекулярные ковалентные связи, и
применяется для ковалентного сшивания контактирующих белковых молекул. После сшивания
препаратов белка Rpf разными концентрациями глутаровым альдегидом в течении 2 минут, они
были фракционированы в денатурирующих условиях (SDS-электрофорез), которые гарантировали
разъединение нековалентпо сшитых белковых молекул. При этом после обработки глутаровым
альдегидом в препаратах Rpf появлялась белковая полоса с удвоенным молекулярным весом по
сравнению с контролем (не обработанный препарат Rpf), что может свидетельствовать о
ковалентном "сшивании" глутаровым альдегидом рядом расположенных индивидуальных молекул
белка, образующих димер (Рис. 6), Также на гелях видны полосы на границе концентрируюшего и
разделяющего гелей, которые, скорее всего, соответствуют крупным агрегатам, неспособным
войти в разделяющий гель. Видимо, димерная форма характерна для Rpf , что в свою очередь
может свидетельствовать о сродстве мономеров Rpf друг к другу. В ходе работ описанных ниже
было обнаружено наличие димеров и у нативного белка, выделенного из культуральной жидкости
М luteus,
47
U% 0,25% 0,5% 1% 2% 0% Маркеры
, мта-шь эле1егрофорсз прешр&т& рско глутаровым алъисгидом. 'Треки I -5 - увеличивающаяся альдешда (0.1-2%); трек №6 - контрольный препарат
после сшмванн;
«тред юнг указана полоса,
Ир&дьщущи® мс&шжншшя* прошд&вдщшся ообшествд с ешоим>э брп'пшааши коллшши,
ггроптаонс!ририщли шличик Rpi' шз mw>?.px.naciw юетОк MJni&m 2002). Скорее кеет, параду с пулом бел&&; находящимся в культуралыюй жидкости, существует пул Rpf при&феплешюга к юиехочшж стеике микроорганизма* и* эддажноя обр&т^сщий на новерхнснл'и клетки своеобразную выстилку. Возможнее обнаруженная склонность Rof к
47
48
именно этим. В этом смысле, по предположению М. В. Телкова, поверхностный слой, состоящий из Rpf, может напоминать S-белки бактерий (Bahl et al, 1997).
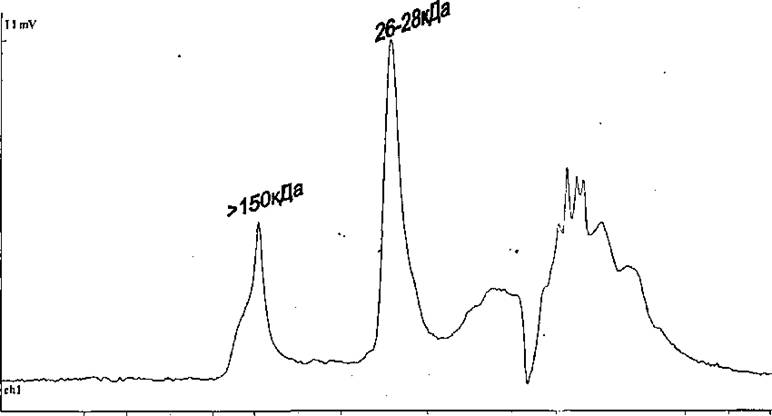
После того, как было установлено, что рекомбинантный белок так склонен к агрегации, была сделана попытка выработать процедуру, которая позволила бы увеличить долю мономерной формы Rpf в препаратах агрегированного в результате хранения белка. В результате подбора условий было установлено, что обработка препарата белка бМ мочевиной в присутствии 50 мМ дитиотриэтола (ДТТ), (агента, обратимо восстанавливающего S-S связи в белках), в течение 2-х часов, и последующий диализ приводят к переходу части агрегатов в мономерную форму (Рис. 7).
| I 3 4 5 6 7 8 9 . 1» II 12 13 14 15 16 17 шщ
Рис.7, Профиль элюции рекомбинантного Rpf, подвергнутого обработке ДТТ и мочевины, при HPLC гель-фильтрации на колонке TSK-gel 2000, (указаны мол. массы, соответствующие пикам).
Так^ке было обнаружено, что мономерная форма белка нестабильна и подвергалась деградации при хранении при положительных температурах. Рис. 8 иллюстрирует распад Rpf при + 4 °С, зафиксированный с помощью электрофореза в полиакриламидном геле в присутствии SDS, Еслц белок длительное время хранился в условиях, способствующих его распаду, он переставал детектиро ваться эле ктрофор ети ч ески .
49
.V—**~«*
,«,-»"-»
68кДа
42кДа
25кДа

|

!$*-• ^^
14кДа 12кДа
Рис, 8. З'ПЯ-Х'Юктроф^рсгр^лтд ^лнхпр'ирующая распад Rpf На треке №1 препарат белка. за^торожентши nps-i -■ 20 'С в 50% глицерине сразу после зшделеЕ-шя, на треке №2 тог же бс;юк, оставленный па недсято при Н- 4 "С Оба препарата не дстактировались при гель ПР1.С фильтрации iiti колонке Bioscp 20OO Трек }Ю - маркеры молекулярной массы,
Для разных ирегтрктоп скорость деградации КрГ варыфинала, что возможно связано с оео5сн постам и фоллинга рекомоанантного бепкд в ктгадом конкретном Езреиарате. На рис, Н-предсъявлеи профиль хпоег-щ раесшишегося мономера Rpf. EiaiisKmcc ярко характеризующий деградацию Склка. следует ота*г 1шъя что и случае с этим препаратом деградация пройдошна очень £i>ieipo: i* течение днух 'шсои при комшгшон температуре.
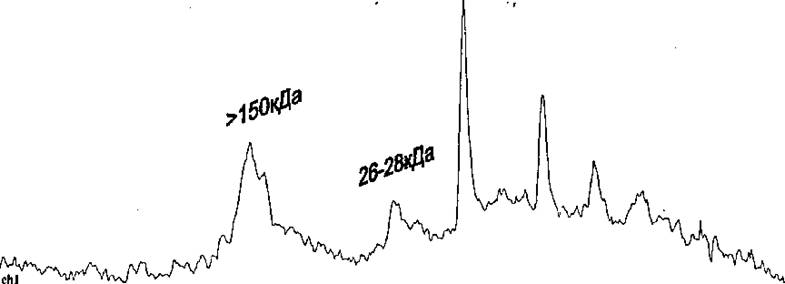
Как ви;пго ю рис, 9 некоторая часть эдономерной форм!.! агрегируют р форму с мол, массой бола: 150 кда. а остюшая масса белка распадается на мелкие фрагменты, различной молекулярной массы*
 |
| 50 |
| ].2mV |
| «/# |
10 И 12 T3 14 l'fl 16 17 rihii
г
к) Рис. 9. Профиль элюции изолированной мономериой формы Rpf (изначальная мол. масса 26-
28 кДа), хранившейся при комнатной температуре в течение 2-3 часов, HPLC гель-фильтраиия на
колонке TSK-gcl 2000, (указаны мол. массы, соответствующие максимумам).
Совместное с нашими коллегами из Института теоретической и экспериментальной физики
РАН изучение препаратов Rpf при помощи атомного силового микроскопа также
продемонстрировало наличие крупных агрегатов в препаратах белка (Рис, 10). Атомно-силовая
микроскопия позволяет изучать биологические объекты в условиях близких к нативным и при этом
обладает чрезвычайно высоким разрешением (менее чем 1 им) (Schneider et al. 1998). При
использовании этого метода получается трехмерная картина объектов, нанесенных на
специальную подложку, при этом можно измерить объем этих объектов. Использование
специального математического аппарата (Schneider et al. 1998), позволяет рассчитать
молекулярную массу белковых объектов в зависимости от их объема. Как видно из рис 10 в
■
препаратах как полноразмерного белка Rpf так и особенно его части (консервативный домен)
5!
!грисутст8у[от Eiiperaibf. соответствующие д^шерам или -гримерам Rpf и полимерам ком серва-плшсго домена содержащим от 15 до 80 молекул.
| А Б |
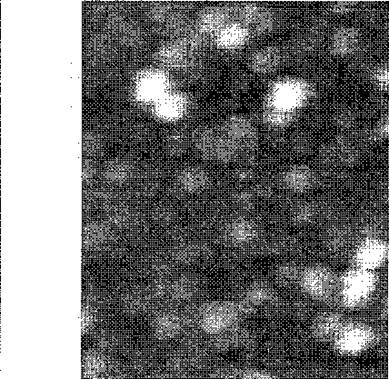
РиЫО. Атомно-сплов&я! микроскопия. А- препарат рекомбипаптного ~Rpf+ более яркие сиепше п%тт\ согласно расчетам пм&ют мол. массу порядка 70 кДа. более бле/шые - порядка 30. В -препарат рекг^^пштюй трункиронашюи формы }'£ (оке irpcc при сапный j? lixoH. п очшшшын фрагмент молекулы Rpf, содер^г,.Щ1ш вишеерштнигзый додан (см. 1jt-jc2)) Оодее яркие светлые пятнп соо^егствузотмол массе нередка 3S0 кДа, Счэлсе Плодные- порядка 170 кДн.
вых беякй н&тшного, После выяснения того, сколь неоднозначно и пршотлшю пт&я&иш рштбшштюю Kpt было реошю сравнить его с секрегируемым &ошм, выделенным из культуроьнои ж&щкоогпMfcroccKcus lutetob
Дня этот использовали бшилег очные суиерштшгш растущих культур М bfeus разных шкшшсгш я пшщериш их гень-фшш] ранни на ирешрпшшой колоните Bio&ep 2000 (HN,€) , j^актируй Rpf в злюируемых фракциях щт помогши им^уноф^рмшшюга анализа (ИФА) с поликлональнымн кроличьими антителами против Rpf. (Рис 11)
Рис. 11. Профили jjnomin бесклеточных супернагантои культур М. luteus разной плотности, выращенных на диали.юванной Broth Е. ОТ)ыю — А — ] .5, В — 2.7. С — 4.5. В чонах, обозначенных подчеркиванием. при помощи ИФА, был выявлен Rpf. НРТ.С гел?,-филр,тратщя на препаративной колонке BioSep 2000. D- данные иммуноблоттинга. подтверждающие наличие Rpi во фракциях, дающих положительную реакцию в ELISA (максимально интенсивные полосы соответствуют фракциям с 22 по 24 минуту, крайний правый трек - контрольный препарат рекомбинантного Rpf).
На рисунке представлены типичные результаты нескольких экспериментов. Как было выяснено максимумы, детектируемые при 280 им в области 21-25 мин, совпадали с максимумами FXTSA (не
показано), что свидетельствует о гом, что они принадлежат Rpf,
Анализ данных, полученных в результате этого 'эксперимента, приводит к ныводу о том, что п суперпатаптах Rpf содержится в основном в двух, формах: мономерной и димерной. Причем соотношение этих форм меняется на разных стадиях роста культуры; в ранней логарифмической фазе преобладает мономерная форма белка, в середине логарифмической фазы большая часть
55 белка пребывает в димерной форме, которая позже опять сменяется мономером. Некоторое
отклонение молекулярных масс белка от рассчитанных в соответствии со структурой гена его
кодирующего, по нашему мнению может быть следствием песферической формы молекул Rpf
Так, по предположениям наших британских коллег из лаборатории М. Янга, сделанным на
основании анализа первичной последовательности Rpf, весьма вероятно, что белок имеет
гантелеобразную форму. Это обусловлено тем, что белок содержит два домена (консервативный
домен Rpf и LysM-домен), соединенные между собой перемычкой.
Таким образом, на основании результатов изложенных в этом разделе можно сделать вывод о
том, что рекомбинантный Rpf склонен к образованию крупных агрегатов, в то же время нативный
белок способен образовывать димеры и находиться в среде как в мономерной, так и в димерной
форме. Haprfy с этим, было обнаружено, что Rpf в нестерильных условиях подвергается быстрой
лротеолитической - - " " ~~^~ ~-~ ~—-------- ■— - деградации.
56
Еще по теме Глава 2. Объекты и методы исследований:
- ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ ГЛАВА 3 Объекты и методы исследования
- Глава 2. Характеристика объектов и методов исследования
- ГЛАВА 2. Объекты, материалы и методы исследования
- Глава 2 Объект, методы и условия проведения исследований
- Объект, методы и средства исследования.
- Объекты и методы исследования Объекты исследования
- Объекты и методы исследования
- 2.1. Объект, методы и средства исследования.
- Объект и методы исследования
- Объект и методы исследования. Выбор пространства информативных признаков
- Тема №1. Патологическая анатомия: объекты и методы исследования. Аутопсия. Патология клетки как интегративное понятие.
- Объекты исследования
- Глава 2. Материалы и методы исследования
- Цели, задачи и объекты экспериментальных исследований
- Объект исследования
- Глава 8 Методы исследования в клинической неврологии и нейрохирургии
- 2.1 Объекты исследования