Дизайн вырожденных праймеров
В случаях, когда исследователь не имеет информации о нуклеотидной последовательности фрагмента целевого транскрипта и располагает данными только о частичной аминокислотной последовательности белка, амплификация фрагментов целевого гена может быть осуществлена с использованием вырожденных праймеров.
Вследствие вырожденности генетического кода (табл. 2.1), одну аминокислотную последовательность может кодировать несколько нуклеотидных. Вырожденный праймер представляет собой смесь олигонуклеотидов, кодирующих одну и ту же аминокислотную последовательность и отличающихся по одной или нескольким нуклеотидным позициям.
Так, например, если целевой ген кодирует аминокислотную последовательность LFYGW, последовательность вырожденного праймера будет выглядеть следующим образом: (T/C)TN TT(T/C)
TA(T/C) GGN TGG.
Если известны только структуры гомологичных белков из других организмов, для выбора фрагментов аминокислотных последовательностей, пригодных для дизайна вырожденных праймеров, осуществляют их выравнивание (sequence alignments) и находят высококонсервативные блоки. Например, на рис. 2.4 показано множественное выравнивание последовательностей флуоресцентных белков различных коралловых полипов. Подчеркиванием обозначены консервативные участки последовательности, которые могут быть использованы для дизайна вырожденных праймеров, затемнены - высококонсервативные последовательности.
Таблица 2.1. Генетический код
| 1-е основание | 2-е основание | |||
| U (T) | C | A | G | |
| U (T) | UUU (Phe/F) UUC (Phe/F) UUA (Leu/L) UUG (Leu/L) | UCU (Ser/S) UCC (Ser/S) UCA (Ser/S) UCG (Ser/S) | UAU (Tyr/Y) UAC (Tyr/Y) UAA (Стопкодон) UAG (Стопкодон) | UGU (Cys/C) UGC (Cys/C) UGA (Стопкодон) UGG (Trp/W) |
| C | CUU (Leu/L) CUC (Leu/L) CUA (Leu/L) CUG (Leu/L) | CCU (Pro/P) CCC (Pro/P) CCA (Pro/P) CCG (Pro/P) | CAU (His/H) CAC (His/H) CAA (Gln/Q) CAG (Gln/Q) | CGU (Arg/R) CGC (Arg/R) CGA (Arg/R) CGG (Arg/R) |
| A | AUU (Ile/I) AUC (Ile/I) AUA (Ile/I) AUG (Met/M) | ACU (Thr/T) ACC (Thr/T) ACA (Thr/T) ACG (Thr/T) | AAU (Asn/N) AAC (Asn/N) AAA (Lys/K) AAG (Lys/K) | AGU (Ser/S) AGC (Ser/S) AGA (Arg/R) AGG (Arg/R) |
| G | GUU (Val/V) GUC (Val/V) GUA (Val/V) GUG (Val/V) | GCU (Ala/A) GCC (Ala/A) GCA (Ala/A) GCG (Ala/A) | GAU (Asp/D) GAC (Asp/D) GAA (Glu/E) GAG (Glu/E) | GGU (Gly/G) GGC (Gly/G) GGA (Gly/G) GGG (Gly/G) |
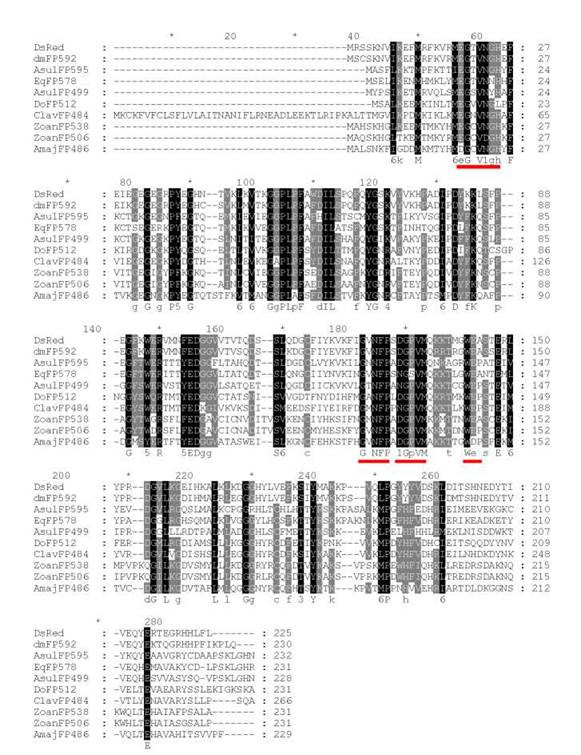
Рис. 2.4.
Множественное выравнивание последовательностей флуоресцентных белков различных коралловых полипов2.4.2. Амплификация 5'-концевого фрагмента кДНК, 5' RACE
Описанный ранее синтез кДНК со сменой матрицы позволяет получать первую цепь кДНК, содержащую известную последовательность адаптера на обоих концах молекулы. Это решает основную проблему, долгое время ограничивавшую возможности исследователей по идентификации 5'-концевых фрагментов целевых транскриптов, - отсутствие известной последовательности для отжига праймера на 5'- концах молекул мРНК.
Первая цепь кДНК, полученная с помощью метода синтеза со сменой матрицы, может быть использована в нескольких раундах step-out/ nested ПЦР с вырожденными или ген-специфическими праймерами, направленными в сторону 5'-конца целевого гена (рис. 2.5). Первичная структура продукта 5'-RACE дает информацию о расположении стартового кодона и длине 5'- концевой нетранслируемой области.
2.2.