цкюбю 30. бхпсяш цепоеяю
цЕПОЕЯБХПСЯШ ПЮЯОПНЯРПЮМЕМШ ЯПЕДХ ЛМНЦХУ БХДНБ ЛКЕЙНґОХРЮЧЫХУ Х ОРХЖ, ХЛЕММН ЩРХ БХПСЯШ МЮХАНКЕЕ ЯРПНЦН ЙКЮЯЯХґТХЖХПНБЮМШ [аЮПХМЯЙХИ х. т., 1982; . Matthews R., 1982; Brown F.,-1986].
нМХ НАЗЕДХМЕМШ Б НАЬХПМНЕ ЯЕЛЕИЯРБН Her-pesviridae,ЙНРНПНЕ -ОНДПЮГДЕКЪЕРЯЪ МЮ РПХ ОНДЯЕЛЕИЯРБЮ: ЮКЭґТЮ-ЦЕПОЕЯБХПСЯШ (Alphaherpesvirinae),БЙКЧВЮЧЫХЕ ПНДШ Simplexvirus(БХПСЯ ОПНЯРНЦН ЦЕПОЕЯЮ), Poikilovirus(БХПСЯ КНФМНЦН АЕЬЕМЯРБЮ) Х Varicellavirus(БХПСЯ БЮПХЖЕККШ-ГНЯРЕ- ПЮ); АЕРЮ-ЦЕПОЕЯБХПСЯШ (Bethaherpesvirinae),БЙКЧВЮЧЫЕЕ ПНДШ Cytomegalovirus(ЖХРНЛЕЦЮКНБХПСЯ ВЕКНБЕЙЮ) Х Murome- galovirus(ЖХРНЛЕЦЮКНБХПСЯ ЛШЬЕИ), Х ЦЮЛЛЮ-ЦЕПОЕЯБХПСЯШ (Gammaherpesvirinae),БЙКЧВЮЧЫЕЕ ПНДШ Lymphocryptovirus (БХПСЯ щОЬРЕИМЮ ≈ аЮПП), Thetacryprolymphovirus(БХПСЯ АНґКЕГМХ лЮПЕЙЮ) Х Rhadonovirus(БХПСЯ ЦЕПОЕЯЮ ЯЮИЛХПХ). б ЙЮФДНЛ ПНДЕ ХЛЕЕРЯЪ НДХМ ХКХ МЕЯЙНКЭЙН БХПСЯНБ. бХПСЯШ ЦЕПОЕЯЮ ХКХ ЯУНДМШЕ Я МХЛХ НАМЮПСФЕМШ РЮЙФЕ С МХГЬХУ ОНґГБНМНВМШУ≈ ОПЕЯЛШЙЮЧЫХУЯЪ, ГЕЛМНБНДМШУ Х ПША, ЯУНДМШЕ C МХЛХ БХПСЯШ БШЪБКЕМШ С ЛНККЧЯЙНБ Х ЦПХАНБ [Fenner F., 1976]. нДМЮЙН Б АНКЕЕ ОНГДМХУ ЙКЮЯЯХТХЙЮЖХЪУ ОНЯКЕДМХЕ БХПСЯШ МЕ СОНЛХМЮЧРЯЪ. рЮЙХЛ НАПЮГНЛ, НЯРЮЕРЯЪ МЕЪЯМШЛ, ОНПЮФЮЧР КХ ЦЕПОЕЯБХПСЯШ РНКЭЙН БШЯЬХУ ОНГБНМНВМШУ (РЕОґКНЙПНБМШУ) ХКХ ФЕ АНКЕЕ ЬХПНЙХИ ЙПСЦ ╚УНГЪЕБ╩ (МХГЬХЕ ОНГБНМНВМШЕ, АЕЯОНГБНМНВМШЕ).
бХПХНМШ ПЮГМШУ ОПЕДЯРЮБХРЕКЕИ ЦЕПОЕЯБХПСЯНБ ДНБНКЭМН ЯУНДМШЕ. нМХ ХЛЕЧР МЕОПЮБХКЭМСЧ ЯТЕПХВЕЯЙСЧ ТНПЛС, ДХЮґЛЕРП 120≈200 МЛ, ЯНЯРНЪР ХГ 4 ЯРПСЙРСПМШУ ЙНЛОНМЕМРНБ: ЩКЕЙРПНММН-ОКНРМНИ ЯЕПДЖЕБХМШ, ХЙНЯЮЩДПЮКЭМНЦН МСЙКЕНЙЮО- ЯХДЮ, ЩКЕЙРПНММН-ОКНРМНИ БМСРПЕММЕИ НАНКНВЙХ (tegument)Х БМЕЬМЕИ НАНКНВЙХ, ХКХ ЛЕЛАПЮМШ (envelope).яЕПДЖЕБХМЮ ЯНґЯРНХР ХГ дмй, ЮЯЯНЖХХПНБЮММНИ Я АЕКЙЮЛХ. йЮОЯХД ХЛЕЕР ДХЮґЛЕРП 100≈110 МЛ, ТНПЛС ХЙНЯЮЩДПЮ Х ЯНЯРНХР ХГ 162 ЙЮОЯНЛЕ- ПНБ, ХГ ЙНРНПШУ 150 ЪБКЪЧРЯЪ ЦЕЙЯЮЛЕПЮЛХ Х 12 ОЕМРЮЛЕПЮЛХ; НМХ ПЮГЛЕЫЮЧРЯЪ ОН 5 МЮ ЙЮФДНИ ТЮЯЕРЙЕ (edge).йЮОЯНЛЕПШ ХЛЕЧР БХД ОНКШУ РПСАНЙ ПЮГЛЕПНЛ 2,5?12,5 МЛ Я ЙЮМЮКНЛ ДХЮЛЕРПНЛ 4 МЛ.
бМСРПЕММЪЪ НАНКНВЙЮ ЯНЯРНХР ХГ ЯКНЪ АЕКґЙНБШУ ЦКНАСКЪПМШУ ЛНКЕЙСК, МЮПСФМЮЪ ЪБКЪЕРЯЪ ДБСЯКНИМНИ КХОХДМНИ ЛЕЛАПЮМНИ, Б ЙНРНПСЧ БЯРПНЕМШ АЕКЙНБШЕ БШЯРСОШ [Wildy P., 1986].цЕМНЛ ОПЕДЯРЮБКЪЕР ЯНАНИ КХМЕИМСЧ ДБСМХРЕБСЧ дмй, ЛНґКЕЙСКЪПМЮЪ ЛЮЯЯЮ ЙНРНПНИ ЙНКЕАКЕРЯЪ НР 86?IO6(БХПСЯ ЦЕПґОЕЯЮ ЯНЛЮ) ДН 145?106(ЖХРНЛЕЦЮКНБХПСЯ ВЕКНБЕЙЮ). яУЕЛЮРХґВЕЯЙХ НМ ЯНЯРНХР ХГ ДБСУ ЙНБЮКЕМРМН ЯНЕДХМЕММШУ ЯЕЦЛЕМРНБ ≈ ДКХММНЦН (L)Х ЙНПНРЙНЦН (S)≈ЯННРБЕРЯРБЕММН 82 Х 18% ЦЕґМНЛЮ (ПХЯ. 56). йЮФДШИ ЙНЛОНМЕМР Б ЯБНЧ НВЕПЕДЭ ЯНЯРНХР ХГ СМХЙЮКЭМШУ (UlХ Us)ОНЯКЕДНБЮРЕКЭМНЯРЕИ, ╚НАПЮЛКЕММШУ╩ ХМБЕПРХПНБЮММШЛХ ОНБРНПЮЛХ abХ э'Ю' ДКЪ ЙНЛОНМЕМРЮ LХ Ю'Я' Х ЯЮ ДКЪ ЙНЛОНМЕМРЮ S [Spectoret D. et al., 1982]. оНЯКЕґДНБЮРЕКЭМНЯРХ L(НЙНКН 500 МСЙКЕНРХДНБ) НАЕЯОЕВХБЮЧР ЖХПґЙСКЪЖХЧ дмй- пЮГМЮЪ ЯРШЙНБЙЮ L-Х S-ЯЕЦЛЕМРНБ Б ГЮБХЯХґЛНЯРХ НР' ХУ ЛЕЯРЮ ПЮЯОНКНФЕМХЪ Х НПХЕМРЮЖХХ НАСЯКНБКХБЮЕР НАПЮГНБЮМХЕ 4 ХГНЛЕПМШУ ТНПЛ ЦЕМНЛЮ ЦЕПОЕЯБХПСЯНБ.

пХЯ. 56. яРПСЙРСПЮ ЦЕМНЛЮ БХПСЯЮ ОПНЯРНЦН ЦЕПОЕЯЮ (ЯУЕЛЮ). аСЙБЮЛХ НАНґГМЮВЕМШ ОПЪЛШЕ Х ХМБЕПРХПНБЮММШЕ ОНБРНПШ, ЯРПЕКЙЮЛХ ОНЙЮГЮМЮ ХУ НПХґЕМРЮЖХЪ; 1 Х 2 ≈ ЙНБЮКЕМРМН ЯБЪГЮММШЕ ТПЮЦЛЕМРШ ЦЕМНЛЮ, ПЮГДЕКЕММШЕ ХМґБЕПРХПНБЮММШЛХ ОНБРНПЮЛХ. б ОНОСКЪЖХХ БХПСЯЮ ЯНДЕПФЮРЯЪ 4 БЮПХЮМРЮ ЮПЮМФХПНБЙХ ЦЕМНЛЮ Б ЩЙБХЛНКЪПМШУ ЙНМЖЕМРПЮЖХЪУ, ПЮГКХВЮЧЫХУЯЪ ОН НПХЕМРЮЖХХ 1-ЦН Х 2-ЦН ТПЮЦЛЕМРНБ.
Н
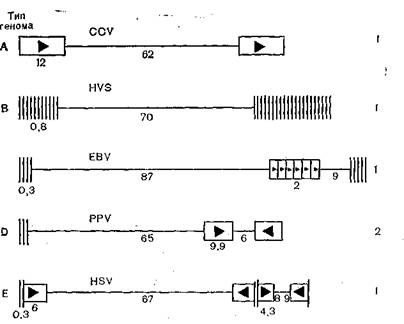
пХЯ. 57. юПЮМФХПНБЙЮ ОНЯКЕДНБЮРЕКЭМНЯРЕИ Б ЦЕМНЛЮУ 5 ЦЕПОЕЯБХПСЯНБ (ЯУЕґЛЮ). цНПХГНМРЮКЭМШЛХ КХМХЪЛХ НАНГМЮВЕМШ СМХЙЮКЭМШЕ ХКХ ОЯЕБДН СМХЙЮКЭґМШЕ НАКЮЯРХ; ОПЪЛНСЦНКЭМХЙЮЛХ НАНГМЮВЕМШ ОНБРНПЪЧЫХЕЯЪ ОНЯКЕДНБЮРЕКЭґМНЯРХ, РПЕСЦНКЭМХЙХ ОНЙЮГШБЮЧР, ОНБРНПЪЕРЯЪ КХ ОНЯКЕДНБЮРЕКЭМНЯРЭ Б ОПЪґЛНИ ХКХ НАПЮРМНИ НПХЕМРЮЖХХ; ЖХТПШ ЯОПЮБЮ ≈ ТПЮЦЛЕМРШ ЦЕМНЛЮ.
яНДЕПФЮМХЕ ц+ж С ПЮГМШУ ОПЕДЯРЮБХРЕКЕИ ЦЕПОЕЯБХПСЯНБ ЙНКЕАКЕРЯЪ Б ЬХПНЙХУ ОПЕДЕКЮУ; ПЮГКХВМШ БЕКХВХМЮ ЦЕМНЛЮ Х ЕЦН НЦЮМХГЮЖХЪ [Honess R., 1984] (ПХЯ.
57). с НДМНЦН Х РНЦН ФЕ БХПСЯЮ 1B ПЕГСКЭРЮРЕ ОЕПЕЯРПНЕЙ ЦЕМНЛШ ЛНЦСР БЮПЭХПНБЮРЭ ЙЮЙ КН ПЮГЛЕПЮЛ, РЮЙ Х ОН ЯРПСЙРСПЮЛ. рЮЙ, С ЦЕПОЕЯЮ ЯЮИЛХ- ПХ ЦЕМНЛ БЯРПЕВЮЕРЯЪ Б ДБСУ БЮПХЮМРЮУ: л, МЮ 70% ЯНЯРЮБКЕМґМШИ ХГ КЕЦЙНИ ЖЕОХ, Х м, ЯНЯРЮБКЕММШИ ХЯЙКЧВХРЕКЭМН ХГ РЪґФЕКНИ ЖЕОХ [Bornkamm G. et al., 1976]. с ПЮГМШУ ЬРЮЛЛНБ БХПСЯЮ щОЬРЕИМЮ ≈ аЮПП МЮАКЧДЮЧРЯЪ ПЮГКХВХЪ Б ЯРПНЕМХХ ЦЕМНЛЮ [Given D., Kieff E., 1978], Ю С ЖХРНЛЕЦЮКНБХПСЯЮ ПЮГґЛЕПШ ЦЕМНЛЮ ЛНЦСР ЙНКЕАЮРЭЯЪ НР 100?106ДН 155?106 [Kilk- patrick б., Huang E., 1977]. б ЯБЪГХ Я ОНДБХФМНЯРЭЧ ЦЕМНЛЮ БНГЛНФМШ ПЮГМННАПЮГМШЕ ЕЦН ОЕПЕЯРПНИЙХ ГЮ ЯВЕР ДСОКХЙЮЖХХ Х !ОЕПЕЛЕЫЕМХЪ НРДЕКЭМШУ ЦЕМНБ [Pogue-Geile й. et al,, 1985], гМЮВХРЕКЭМШ Х ЙЮВЕЯРБЕММШЕ ОЕПЕЯРПНИЙХ ЦЕМНБ ЦЕПОЕЯБХПСґЯНБ. рЮЙ, НАКЮЯРХ ЦЕМНБ gF, gCБХПСЯЮ ЦЕПОЕЯЮ ВЕКНБЕЙЮ ЯЕПН- БЮПЮ ХЛЕЧР ГМЮВХРЕКЭМСЧ ЦНЛНКНЦХЧ [Dowbenko D., Lasky L.,
1984] . б ЯНЯРЮБЕ ЦЕМНЛЮ БХПСЯЮ ОПНЯРНЦН ЦЕПОЕЯЮ РХОЮ / ХЛЕґЕРЯЪ РЮМДЕЛМН ОНБРНПЪЕЛШИ РПХОКЕР ццж, ЯСЫЕЯРБНБЮМХЕ ЙНґРНПНЦН УЮПЮЙРЕПМН ДКЪ ПХАНЯНЛМНИ 28S PHK ВЕКНБЕЙЮ.
б ЯНЯРЮБЕ БХПХНМНБ НАМЮПСФЕМН АНКЕЕ 30 АЕКЙНБ: 7 ЦКХЙНґОПНРЕХДНБ (g∙B, gC, gD, gE, gF, gGХ gX)МЮУНДЪРЯЪ МЮ ОНґБЕПУМНЯРХ Х БШГШБЮЧР НАПЮГНБЮМХЕ БХПСЯМЕИРПЮКХГСЧЫХУ ЮМґРХРЕК [Vaughan P. et al., 1985]; 6 АЕКЙНБ ХЛЕЧРЯЪ Б ЙЮОЯХДЕ, ЯПЕДХ МХУ юртЮГЮ Х ОПНРЕХМЙХМЮГЮ. лМНЦХЕ ДЕЯЪРЙХ АЕКЙНБ, Б РНЛ ВХЯКЕ РХЛХДХМЙХМЮГЮ, ЪБКЪЧРЯЪ МЕЯРПСЙРСПМШЛХ Х НАґПЮГСЧРЯЪ Б УНДЕ ПЕОПНДСЙЖХХ БХПСЯЮ. яПЕДХ ДПСЦХУ АЕКЙНБ ЯКЕДСЕР МЮГБЮРЭ дмй-ОНКХЛЕПЮГС Х АЕКНЙ, ЯБЪГЮММШИ Я дмй.
пЕОКХЙЮЖХЪ БХПСЯНБ ЦЕПОЕЯЮ ЪБКЪЕРЯЪ ЯКНФМШЛ Х ЛМНЦНґЯРСОЕМВЮРШЛ ОПНЖЕЯЯНЛ Х БЙКЧВЮЕР Б ЯЕАЪ ОПХЙПЕОКЕМХЕ БХґПХНМНБ Й ЙКЕРНВМШЛ ПЕЖЕОРНПЮЛ, ЩМДНЖХРНГ Х ЯКХЪМХЕ ЛЕЛАґПЮМ БХПХНМНБ Х ЙКЕРЙХ. б ПЕГСКЭРЮРЕ НАМЮФЮЕРЯЪ ЙЮОЯХД, ЙНґРНПШИ ОНОЮДЮЕР Б ЖХРНГНКЭ. гЮРЕЛ ЙНЛОКЕЙЯ дмй≈: АЕКНЙ ОПНМХЙЮЕР Б ЪДПН, ЙЮОЯХД ПЮГПСЬЮЕРЯЪ Х БХПХНММЮЪ дмй БШґУНДХР Б МСЙКЕНОКЮГЛС Х ГДЕЯЭ МЮВХМЮЕР ТСМЙЖХНМХПНБЮРЭ, РПЮМЯЙПХАХПСЪЯЭ ЙКЕРНВМНИ пмй-ОНКХЛЕПЮГНИ. пЮГКХВЮЧР ЯБЕПУПЮММЧЧ, ПЮММЧЧ Х ОНГДМЧЧ РПЮМЯЙПХОЖХЧ, ОПНЖЕЯЯХМЦ Лпмй, ЯХМРЕГ ЙНДХПСЕЛШУ ОПНДСЙРНБ Х ВЮЯРХВМШИ НАПЮРМШИ РПЮМЯОНПР ХУ Б ЪДПН, ПЕОКХЙЮЖХЧ дмй Х ТНПЛХПНБЮМХЕ ДНґВЕПМХУ ЛНКЕЙСК, НАПЮГНБЮМХЕ Б ЪДПЮУ ЙКЕРНЙ МЕГПЕКШУ ЙЮОЯХґДНБ, !ОНВЙНБЮМХЕ ХУ ВЕПЕГ ЪДЕПМСЧ ЛЕЛАПЮМС, ТНПЛХПНБЮМХЕ ГПЕКШУ ЙЮОЯХДНБ Х БМЕЬМЕИ НАНКНВЙХ Б ЖХЯРЕПМЮУ ЩМДНОКЮГґЛЮРХВЕЯЙНЦН ПЕРХЙСКСЛЮ, РПЮМЯОНПР Й ОНБЕПУМНЯРХ ВЕПЕГ ЛНґДХТХЖХПНБЮММШИ ЩМДНОКЮГЛЮРХВЕЯЙХИ ПЕРХЙСКСЛ Х БШУНД ХГ ЙКЕРЙХ (ПХЯ.
58).щЙЯОПЕЯЯХЪ БХПСЯМШУ ЦЕМНБ ОПНХЯУНДХР ОНЩРЮОМН Х ЯНОПНґБНФДЮЕРЯЪ БШЙКЧВЕМХЕЛ ЯХМРЕГЮ ЙКЕРНВМШУ ЛЮЙПНЛНКЕЙСК. яБЕПУПЮММХЕ Х ПЮММХЕ ЯХМРЕГШ ОПЕДЬЕЯРБСЧР ПЕОКХЙЮЖХХ дмй. мЮ ЩРНИ ЯРЮДХХ ЯХМРЕГХПСЧРЯЪ ПЮГМШЕ ТСМЙЖХНМЮКЭМШЕ (МЕЯРПСЙРСПМШЕ) АЕКЙХ, ЯПЕДХ МХУ РХЛХДХМЙХМЮГЮ Х ДПСЦХЕ АЕКЙХ ЯХМРЕГЮ дмй, Ю РЮЙФЕ ТЕПЛЕМРШ, ОНДЮБКЪЧЫХЕ ЙКЕРНВґМШЕ ЛЮЙПНЛНКЕЙСКЪПМШЕ ЯХМРЕГШ. щЙЯОПЕЯЯХЪ ПЮММХУ (ЮКЭТЮ) ЦЕМНБ ЯНОПНБНФДЮЕРЯЪ ЯХМРЕГНЛ 5 ЮКЭТЮ-АЕКЙНБ. гЮРЕЛ ЯКЕДСґЧР ЯХМРЕГШ 8 АЕРЮ-ОЕОРХДНБ, БЙКЧВЮЪ АНКЭЬНИ дмй-ЯБЪГШБЮ-
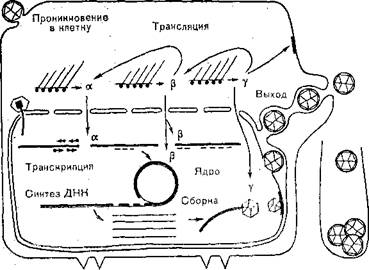
пХЯ. 58. пЕОКХЙЮРХБМШИ ЖХЙК БХПСЯЮ ЦЕПОЕЯЮ (ЯУЕЛЮ).

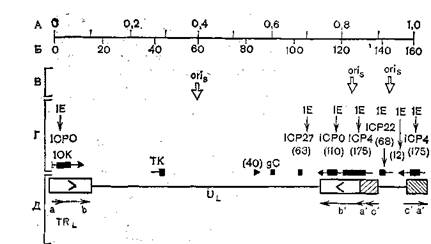
пХЯ. 59. пЮЯОНКНФЕМХЕ МЕЙНРНПШУ ЦЕМНБ МЮ ЦЕХНЛЕ БХПСЯЮ ЦЕПОЕЯЮ.
рЕПЛХМЮКЭМШЕ ОНБНПНРШ Х ХУ НПХЕМРЮЖХЪ ОНЙЮГЮМШ АСЙБЮЛХ Х ЯРПЕКЙЮЛХ; IE ≈ Ю-ЦЕ- МШ; TK,/нй≈п-ЦЕМШ; gC ≈ ОНГДМХЕ ЦЕМШ; ХУ НПХЕМРЮЖХЪ ОНЙЮГЮМЮ РЕЛМШЛХ ОПЪЛНґСЦНКЭМХЙЮЛХ. Х ЯРПЕКЙЮЛХ; Б ЯЙНАЙЮУ ≈ ЛНКЕЙСКЪПМЮЪ ЛЮЯЯЮ (?103); ю ≈ ЕДХМХЖШ ЙЮПРШ; а ≈ ОЮПШ (ЙА); б ≈ МЮВЮКН ПЕОКХЙЮЖХХ; ц ≈ КНЙЮКХГЮЖХЪ ЦЕМНБ; д ≈НЯМНБМШЕ ЯРПСЙРСПМШЕ ЦЕМШ,
ЧЫХИ АЕКНЙ, БХПСЯМСЧ ПХАНМСЙКЕНРХДПЕДСЙРЮГС, РХЛХДХМЙХМЮ- ГС Х дмйОНКХЛЕПЮГС.- оНГДМХЕ ЦЮЛЛЮ-АЕКЙХ ЯХМРЕГХПСЧРЯЪ ОНЯКЕ ПЕОКХЙЮЖХХ БХПСЯМНИ дмй; ЩРН, ОНЛХЛН ЛМНЦХУ ДПСЦХУ, АНКЭЬНИ АЕКНЙ ЙЮОЯХДЮ Х ЦКХЙНОПНРЕХД gC.рПЮМЯЙПХОЖХЪ БХґПСЯМШУ ЦЕМНБ НАЕЯОЕВХБЮЕРЯЪ ЙКЕРНВМНИ пмй-ОНКХЛЕПЮГНИ II. нАПЮГНБЮМХЕ ЙЩО-ЯРПСЙРСП, ХУ ЛЕРХКХПНБЮМХЕ, ОНКХЮДЕМХКХПНґБЮМХЕ БХПСЯМШУ Лпмй РЮЙФЕ ЙЮРЮКХГХПСЧРЯЪ ЙКЕРНВМШЛХ ТЕПЛЕМРЮЛХ.
мЕЙНРНПШЕ ЦЕМШ БХПСЯЮ ОПНЯРНЦН ЦЕПОЕЯЮ РХОЮ I ХДЕМРХТХґЖХПНБЮМШ. пЮЯОНКНФЕМХЕ ХУ МЮ ТХГХВЕЯЙНИ ЙЮПРЕ ЦЕМНЛЮ ОНґЙЮГЮМН МЮ ПХЯ. 59, ЯРПЕКЙЮЛХ СЙЮГЮМН МЮОПЮБКЕМХЕ ХУ РПЮМЯґЙПХОЖХХ. йЮЙ БХДМН ХГ ЩРНИ ЯУЕЛШ, ЯБЕПУПЮММХЕ, ПЮММХЕ Х ОНГДМХЕ ЦЕМШ ДХЯОЕПЦХПНБЮМШ ОН БХПСЯМНИ УПНЛНЯНЛЕ Х ХЛЕґЧР ПЮГМСЧ НПХЕМРЮЖХЧ.
б ПЕОКХЙЮЖХХ БХПСЯМНИ дмй НЯМНБМСЧ ПНКЭ ХЦПЮЕР ЯХЯРЕґЛЮ ПЕОКХЙЮЖХХ ЙКЕРНВМНИ дмй, НДМЮЙН Б ЕЕ НЯСЫЕЯРБКЕМХХ БЮФМНЕ ГМЮВЕМХЕ ХЛЕЧР БХПСЯМШЕ ЦЕМШ Х ХУ ОПНДСЙРШ, Б ВЮЯРґМНЯРХ РХЛХДХМЙХМЮГЮ.
аНКЕЕ РНЦН, ПЕОКХЙЮЖХЪ БХПСЯМНИ дмй ЙЮРЮКХГХПСЕРЯЪ БХПСЯМНИ дмй-ОНКХЛЕПЮГНИ, ЙНРНПЮЪ БГЮХЛНґДЕИЯРБСЕР Я БХПСЯХМДСЖХПНБЮММШЛ дмй-ЯБЪГШБЮЧЫХЛ АЕКЙНЛ, НАПЮГСЧЫХЛ ЙНЛОКЕЙЯШ Я дмй, БШЪБКЪЕЛШЕ Я ОНЛНЫЭЧ ЩКЕЙґРПНММНИ ЛХЙПНЯЙНОХХ. нДМЮЙН Б ЮЙРХБМН ПЮГЛМНФЮЧЫХУЯЪ ЙКЕРЙЮУ ПЕОКХЙЮЖХЪ БХПСЯМНИ дмй ЛНФЕР НАЕЯОЕВХБЮРЭЯЪ Х ОПХ ДЕКЕМХХ АНКЭЬХУ (ДН 15≈20%) СВЮЯРЙНБ БХПСЯМНЦН ЦЕМНґЛЮ, МЮОПХЛЕП, НАКЮЯРХ РХЛХДХМЙХМЮГМНЦН ЦЕМЮ. б РН ФЕ БПЕЛЪ ЦЕМШ ЦПСООШ АЕРЮ ХЦПЮЧР АНКЭЬСЧ ПНКЭ Б ПЕЦСКЪЖХХ БХПСЯ- ХМДСЖХПНБЮММШУ ЯХМРЕГНБ Х ХУ БШЙКЧВЕМХХ.. б МЕДЕКЪЫХУЯЪ ЙКЕРЙЮУ БХПСЯШ ЦЕПОЕЯЮ, КХЬЕММШЕ ВЮЯРХ ЯБНХУ ПЮММХУ ЦЕМНБ, МЕ ЯОНЯНАМШ Й ПЕОКХЙЮЖХХ Х ЯСЫЕЯРБСЧР Б БХДЕ ОКЮГЛХДННАґПЮГМШУ ТНПЛ, ЙЮЙ ЩРН ХЛЕЕР ЛЕЯРН Б МЕПБМШУ ЙКЕРЙЮУ [Roiz- man б. et al., 1985].йЮЙ Х ЯКЕДНБЮКН НФХДЮРЭ, ОПХ РЮЙНИ БЕКХВХМЕ ЦЕМНЛЮ, ЙЮґЙСЧ ХЛЕЧР ЦЕПОЕЯБХПСЯШ, ХУ ПЕОКХЙЮЖХЪ МЕ ЛНФЕР АШРЭ МЕґОПЕПШБМНИ Х ОПНХЯУНДХР, РЮЙ ЯЙЮГЮРЭ, ЙБЮМРЮЛХ, ЕДХМХЖЕИ ЙНґРНПШУ ЪБКЪЕРЯЪ ПЕОКХЙНМ. яОЕЖХЮКЭМН ОПНБЕДЕММШЕ ХЯЯКЕДНґБЮМХЪ ОНЙЮГЮКХ, 'ВРН РЮЙХЛ ПЕОКХЙНМНЛ ЪБКЪЕРЯЪ СВЮЯРНЙ дмй, ЯНДЕПФЮЫХИ МЕ АНКЕЕ 15000 ОЮП МСЙКЕНРХДНБ [Kwong A., Frenґkel N., 1984].
оСЯРШЕ БХПСЯМШЕ ЙЮОЯХДШ ТНПЛХПСЧРЯЪ Б ЪДПЮУ, ГДЕЯЭ ФЕ ОПНХЯУНДХР СОЮЙНБЙЮ Б МХУ БМНБЭ ЯХМРЕГХПНБЮММНИ БХПСЯМНИ дмй, БГЮХЛНДЕИЯРБСЧЫЕИ Я дмй-ЯБЪГШБЮЧЫХЛ БХПСЯМШЛ АЕКЙНЛ. оПХ ОПНУНФДЕМХХ ВЕПЕГ ЪДЕПМСЧ ЛЕЛАПЮМС БХПХНМШ ОПХНАПЕРЮЧР БМСРПЕММЧЧ, Ю ЖПХ БШУНДЕ ХГ ЙКЕРЙХ ≈ БМЕЬМЧЧ НАНКНВЙС.
йКЮЯЯХТХЙЮЖХЪ ЦЕПОЕЯБХПСЯНБ НЯМНБЮМЮ МЮ ЯНБНЙСОМНЯРХ ОПХГМЮЙНБ, БЙКЧВЮЪ ЯРЕОЕМЭ ЦНЛНКНЦХХ, МЮКХВХЕ ЮМРХЦЕММНЦН ПНДЯРБЮ Х АХНКНЦХВЕЯЙХЕ ЯБНИЯРБЮ.
цПСООЮ ЦЕПОЕЯБХПСЯНБ ВЕКНБЕЙЮ ЯНЯРНХР ХГ БХПСЯНБ ЦЕПОЕЯЮ ВЕКНБЕЙЮ ДБСУ ЯЕПНБЮПНБ Х БХПСЯЮ ЦЕПОЕЯЮ ЙНПНБ; ЦПСООЮ ЦЕПґОЕЯБХПСЯНБ ЯБХМЕИ БЙКЧВЮЕР БХПСЯШ КНФМНЦН АЕЬЕМЯРБЮ Х ЮАНПРЮ КНЬЮДЕИ. й ЩРНЛС ФЕ ОНДЯЕЛЕИЯРБС (ЮКЭТЮ-ЦЕПОЕЯБХПСґЯШ) НРМНЯЪР БХПСЯШ БЮПХЖЕККШ-ГНЯРЕП, ЦЕПОЕЯЮ ЖЕМНОХРЕЙНБ, ЯКСВМНИ ЩЙГЮМРЕЛШ КНЬЮДЕИ, ЦЕПОЕЯЮ ЙНЬЕЙ, КНЬЮДХМШИ БХґПСЯ ЦЕПОЕЯЮ РХОЮ 2, БХПСЯ ЦЕПОЕЯЮ ЯНАЮЙ. й ОНДЯЕЛЕИЯРБС АЕРЮ- ЦЕПОЕЯБХПСЯНБ НРМНЯЪРЯЪ ЖХРНЛЕЦЮКНБХПСЯШ ВЕКНБЕЙЮ Х ЛШЬЕИ, ЯНЯРЮБКЪЧЫХЕ ДБЮ НРДЕКЭМШУ ПНДЮ, Ю РЮЙФЕ ЖХРНЛЕЦЮКНБХПСЯШ ЯБХМЕИ, ЛШЬЕИ Х ЛНПЯЙХУ ЯБХМНЙ. нР ОПЕДЯРЮБХРЕКЕИ ОПЕДШґДСЫЕЦН ОНДЯЕЛЕИЯРБЮ ЩРХ БХПСЯШ НРКХВЮЧРЯЪ ЛЕДКЕММНИ ПЕґОПНДСЙЖХЕИ, ТНПЛХПНБЮМХЕЛ ОНД ХУ БНГДЕИЯРБХЕЛ ЦХЦЮМРЯЙХУ ЯКХБЬХУЯЪ ЙКЕРНЙ Б ЙСКЭРСПЮУ.
мЮПЪДС Я ОПНДСЙРХБМНИ ХМТЕЙЖХЕИ, ЯНОПНБНФДЮЧЫЕИЯЪ ЦХґАЕКЭЧ ГЮПЮФЕММШУ ЙКЕРНЙ, ДКЪ ХМТЕЙЖХХ ЦЕПОЕЯБХПСЯЮЛХ УЮґПЮЙРЕПМЮ ОЕПЯХЯРЕМЖХЪ АЕГ ЦХАЕКХ ОНПЮФЕММШУ ЙКЕРНЙ, Ю ЛМНґЦХЕ БХПСЯШ, БШГШБЮЧЫХЕ ЙЮЙ ЛЕЯРМШЕ, РЮЙ Х ЯХЯРЕЛМШЕ ОНПЮґФЕМХЪ, НАКЮДЮЧР БШПЮФЕММШЛХ НМЙНЦЕММШЛХ ОНРЕМЖХЪЛХ. нМЙНЦЕММШЕ ОНРЕМЖХХ МЮАКЧДЮЧРЯЪ С БХПСЯНБ НАНХУ ПЮЯЯЛНРґПЕММШУ ОНДЯЕЛЕИЯРБ, МН Б МЮХАНКЭЬЕИ ЯРЕОЕМХ НМХ БШПЮФЕМШ С ЦЮЛЛЮ-ЦЕПОЕЯБХПСЯНБ ≈ С БХПСЯЮ щОЬРЕИМЮ ≈ аЮПП, Ю РЮЙФЕ ЯУНДМШУ Я МХЛ БХПСЯНБ НАЕГЭЪМ (ЮРЕКЕЯ, ЯЮИЛХПХ), ЦЮЛЛЮ-ЦЕПґОЕЯБХПСЯНБ ОРХЖ (БНГАСДХРЕКЪ АНКЕГМХ лЮПЕЙЮ ЙСП Х ЯУНДМНЦН ГЮАНКЕБЮМХЪ ХМДЧЬЕЙ) Х С ЦЕПОЕЯБХПСЯЮ ЙПНКХЙНБ. нМЙНЦЕМґМШЕ ОНРЕМЖХХ ЯБЪГЮМШ Я НЯНАЕММНЯРЪЛХ ЯРПНЕМХЪ ЦЕМНЛЮ БХПСґЯНБ. рЮЙ, ЦЕМНЛШ НМЙНЦЕММШУ ЦЕПОЕЯБХПСЯНБ НАЕГЭЪМ ЮРЕКЕЯ, ЯЮИЛХПХ Х ЮНРСЯ ХЛЕЧР ОНБРНПМШЕ НАНЦЮЫЕММШЕ ц+ж СВЮЯРЙХ дмй (м-дмй), ЙНРНПШЕ, МЕ НАКЮДЮЪ ЙНДХПСЧЫХЛХ ЯБНИЯРБЮґЛХ, ЪБКЪЧРЯЪ ЯХЦМЮКНЛ ДКЪ СОЮЙНБЙХ ЦЕМНЛЮ. щРХ ОНЯКЕДНБЮґРЕКЭМНЯРХ ПЕЦСКЪПМН НАМЮПСФХБЮЧРЯЪ Б НОСУНКЪУ (КХЛТНЛЮУ), БШГБЮММШУ СЙЮГЮММШЛХ БХПСЯЮЛХ [Fucks P. et al., 1985].
цЕПОЕЯБХПСЯШ БШГШБЮЧР ПЮГМННАПЮГМШЕ ОЮРНКНЦХВЕЯЙХЕ ОПНЖЕЯЯШ: НЯРПШЕ ЛЕЯРМШЕ Х ЯХЯРЕЛМШЕ ОНПЮФЕМХЪ, КЮРЕМРМШЕ ХМТЕЙЖХХ Я ОНПЮФЕМХЕЛ ЖЕМРПЮКЭМНИ МЕПБМНИ ЯХЯРЕЛШ, МЕНОКЮґЯРХВЕЯЙХЕ ОПНЖЕЯЯШ,- нАШВМН ЙКХМХЙЮ .ЦЕПОЕРХВЕЯЙНИ ХМТЕЙЖХХ ОНКХЛНПТМЮ. рЮЙ, ЦЕПОЕЯБХПСЯШ ВЕКНБЕЙЮ НАНХУ ЯЕПНБЮПНБ БШґГШБЮЧР ЯРНЛЮРХР, ОСГШПЭЙНБШЕ ОНПЮФЕМХЪ ЙНФХ Х ЯКХГХЯРШУ НАНКНВЕЙ, ЦЕПОЕЯ ЦКЮГЮ, ОНКНБШУ НПЦЮМНБ Х, БНГЛНФМН, ДЮФЕ ПЮЙ ЬЕИЙХ ЛЮРЙХ (ЦЕПОЕЯБХПСЯ ЯЕПНБЮПЮ 2). оНЯКЕ НЯРПНИ ХМґТЕЙЖХХ БХПСЯ ЛНФЕР .ОЕПЯХЯРХПНБЮРЭ Б МЕПБМШУ Х ДПСЦХУ ЙКЕРґЙЮУ, БШГШБЮЪ ОЕПХНДХВЕЯЙХЕ НАНЯРПЕМХЪ. ■
оЕПЯХЯРЕМЖХЪ ЦЕПОЕЯБХПСЯЮ ВЕКНБЕЙЮ БШГБЮМЮ МЕ РНКЭЙН ОЕПХНДХВЕЯЙХЛ ╚СЯЙЮКЭГШБЮМХЕЛ╩ БХПСЯЮ НР ХЛЛСММШУ ПЕЮЙґЖХИ, МН Х ХГЛЕМВХБНЯРЭЧ БХПСЯЮ, ВРН ДНЙЮГШБЮЕРЯЪ БШДЕКЕМХґЕЛ НРКХВЮЧЫХУЯЪ ЬРЮЛЛНБ БХПСЯЮ НР НДМНЦН Х РНЦН ФЕ АНКЭґМНЦН [Lewis л. et al., 1984]. жХРНЛЕЦЮКНБХПСЯ ВЕКНБЕЙЮ БШГШґБЮЕР КЮРЕМРМСЧ ХМТЕЙЖХЧ Я ПЮГМННАПЮГМШЛ ЙКХМХВЕЯЙХЛ РЕґ
ВЕМХЕЛ (КХУНПЮДЙНИ, ОНПЮФЕМХЕЛ КЕЦЙХУ Х ДПСЦХУ НПЦЮМНБ). оЕПЕДЮБЮЪЯЭ БМСРПХСРПНАМН, БХПСЯ НАСЯКНБКХБЮЕР ПЮГБХРХЕ СПНДЯРБ ОКНДЮ. бХПСЯ щОЬРЕИМЮ ≈ аЮПП С ЕБПНОЕИЖЕБ Х ДПСґЦХУ ,-ОПЕДЯРЮБХРЕКЕИ ЕБПНОЕНХДМНИ ПЮЯШ ВЮЫЕ БЯЕЦН БШГШБЮЕР НЯРПН ОПНРЕЙЮЧЫЕЕ ГЮАНКЕБЮМХЕ Я ПЮГБХРХЕЛ ХЛЛСМХРЕРЮ ОПНґРХБ МЕЦН ≈ ХМТЕЙЖХНММШИ ЛНМНМСЙКЕНГ. с ЮТПХЙЮМЖЕБ НМ МЕґПЕДЙН ЪБКЪЕРЯЪ ОПХВХММНИ ГКНЙЮВЕЯРБЕММНИ КХЛТНЛШ аЕПЙХРґРЮ Х МЮГНТЮПХМЦЕЮКЭМНЦН ПЮЙЮ. бОПНВЕЛ, ПЮЯНБЮЪ НВЕПВЕММНЯРЭ ЩРХУ МНГНКНЦХВЕЯЙХУ ТНПЛ ДЮКЕЙН МЕ ЮАЯНКЧРМЮ. яРНКЭ ФЕ ОНґКХЛНПТМШ Х ЛМНЦНКХЙХ ЦЕПОЕРХВЕЯЙХЕ ГЮАНКЕБЮМХЪ С ФХБНРМШУ.
оНДКХММЮЪ ЩБНКЧЖХНММЮЪ ЙКЮЯЯХТХЙЮЖХЪ ЦЕПОЕЯБХПСЯНБ ОНЙЮ -МЕ ПЮГПЮАНРЮМЮ, РЮЙ ЙЮЙ, Я НДМНИ ЯРНПНМШ, ЦЕМНЛ ХУ ДНґЯРЮРНВМН БЕКХЙ Х ЯКНФЕМ ДКЪ ЮМЮКХГЮ МЮ НЯМНБЕ ЯЕЙБЕМХПНБЮґМХЪ, Я ДПСЦНИ ЯРНПНМШ, ПЮГМШЕ ЦЕМШ ЦЕПОЕЯБХПСЯНБ ЛНЦСР ХЛЕРЭ ПЮГКХВМНЕ ОПНХЯУНФДЕМХЕ. рЮЙ, ДБЕ ЛЮКШЕ PHK БХПСЯЮ щОґЬРЕИМЮ≈ аЮПП ЯУНДМШ Я PHK ЮДЕМНБХПСЯНБ VAIХ VAII [Rose л. et al.,. 1981], ЖХРНЛЕЦЮКНБХПСЯ Х БХПСЯ ЛХЕКНАКЮЯРНГЮ ОРХЖ ХЛЕЧР ЯУНДМШЕ ОНЯКЕДНБЮРЕКЭМНЯРХ [Spector D., Vacqui- er J., 1983] V-mycХ ЯННРБЕРЯРБЕММН Я-РСЯ[Rasmussen R. et al.,
1985] . цЕМНЛ ЖХРНЛЕЦЮКНБХПСЯЮ НАКЮДЮЕР ОНЯКЕДНБЮРЕКЭМНЯРЪґЛХ, ЦНЛНКНЦХВМШЛХ РЮЙНБШЛ Б ЦЕМНЛЕ МНПЛЮКЭМШУ ЙКЕРНЙ [Jeang K-, Hayward G., 1983], Х Р. О. бШЬЕ НРЛЕВЮКНЯЭ МЮКХґВХЕ РЮМДЕЛМШУ ОНЯКЕДНБЮРЕКЭМНЯРЕИ, НАЫХУ ДКЪ дмй ЦЕПОЕЯґБХПСЯЮ Х ЦЕМЮ, ЙНДХПСЧЫЕЦН ЯХМРЕГ ПХАНЯНЛМНИ PHK (28S).
оПХ ХЯЯКЕДНБЮМХХ АХАКХНРЕЙХ ЦЕМНЛЮ ЙКЕРНЙ ВЕКНБЕЙЮ МЮИґДЕМШ СВЮЯРЙХ ЦНЛНКНЦХХ Я ЦЕМНЛНЛ ЖХРНЛЕЦЮКНБХПСЯЮ ВЕКНБЕґЙЮ≈ Б EcoRI,ТПЮЦЛЕМРЮУ R,bХ dБ НАКЮЯРХ ЦЕМНБ ДКЪ ПХАНґЯНЛМНИ пмй. рЮЙЮЪ ЦНЛНКНЦХЪ МЕ БШЪБКЕМЮ ОПХ ЯПЮБМХРЕКЭґМНЛ ХЯЯКЕДНБЮМХХ дмй ЛШЬЕИ Х ЛНПЯЙХУ ЕФЕИ [Shaw G. et al., 1985]. щРН ГЮЯРЮБКЪЕР ДСЛЮРЭ Н ГЮУБЮРЕ ЦЕМНБ ВЕКНБЕЙЮ, Ю МЕ Н АНКЕЕ ДКХРЕКЭМНИ ЩБНКЧЖХХ ДЮММШУ СВЮЯРЙНБ ЦЕМНЛЮ ЖХРНґЛЕЦЮКНБХПСЯЮ. оПХ ХЯЯКЕДНБЮМХХ ЪДЕПМНЦН ЮМРХЦЕМЮ (ЛНКЕЙСґКЪПМЮЪ ЛЮЯЯЮ 72 000) БХПСЯЮ щОЬРЕИМЮ ≈ аЮПП АШКЮ ОНЙЮГЮМЮ ХЛЛСМНКНЦХВЕЯЙЮЪ АКХГНЯРЭ ЕЦН Й ЙКЕРНВМНЛС АЕКЙС (62000) Б ХЛЛСМНАКНРРЕ [Luka J. et al., 1984]. б ДЮММНЛ ЯКСВЮЕ, ОН-БХґДХЛНЛС, ХЛЕЕР ЛЕЯРН ЯУНДМШИ ТЕМНЛЕМ ≈ ГЮУБЮР ЙКЕРНВМШУ ЦЕМНБ Х ХУ ЩБНКЧЖХЪ Б ЯНЯРЮБЕ ЦЕМНЛЮ БХПСЯЮ. б дмй ЖХРНґЛЕЦЮКНБХПСЯЮ ВЕКНБЕЙЮ МЮИДЕМШ 5 СВЮЯРЙНБ ЦНЛНКНЦХХ Я дмй. ЙКЕРНЙ МНПЛЮКЭМШУ РЙЮМЕИ ОКЮЖЕМРШ, ОЕВЕМХ, ЙХЬЕВМХЙЮ, КЕИґЙНЖХРНБ ВЕКНБЕЙЮ, Ю РЮЙФЕ Я дмй ПЮЙНБШУ ЙКЕРНЙ. йПНЛЕ- РНЦН, ЦНЛНКНЦХВМШЕ дмй ЖХРНЛЕЦЮКНБХПСЯЮ ВЕКНБЕЙЮ СВЮЯРЙХ- АШКХ НАМЮПСФЕМШ Б дмй ЦЕПОЕЯБХПСЯНБ ЮНРСЯ Х ЙХРЮИЯЙНЦН, УНЛЪЙЮ, МН МЕ ЛШЬХ. щРХ СВЮЯРЙХ МЕ ЯННРБЕРЯРБНБЮКХ д/Х-ОН- ЯКЕДНБЮРЕКЭМНЯРЪЛ Х ЦЕМЮЛ Я-РСЯ[Rueger R. et al., 1984]. щРН ЯБХДЕРЕКЭЯРБСЕР Н РНЛ, ВРН Б УНДЕ ЩБНКЧЖХХ ЦЕПОЕЯБХПСЯШ ЛНЦґКХ ОНКСВЮРЭ ЦЕМШ ХГ ПЮГМШУ ХЯРНВМХЙНБ Х ОПЕФДЕ БЯЕЦН-ХГ. ЙКЕРНЙ, Б ЙНРНПШУ НМХ ОЮПЮГХРХПНБЮКХ.
бШЬЕ ЦНБНПХКНЯЭ НА НМЙНЦЕММШУ ОНРЕМЖХЪУ ЦЕПОЕЯБХПСЯНБ (ВРН, ЙНМЕВМН, ЯБЪГЮМН Я ХМРЕЦПЮЖХЕИ ХУ ЦЕМНЛНБ Х/ХКХ НРДЕКЭґМШУ ЦЕМНБ), Ю РЮЙФЕ Н БНГЛНФМНЯРХ ХУ ОЕПЯХЯРЕМЖХХ Б БХДЕ ОКЮГЛХД. б ЩРНЛ ЯКСВЮЕ ЛНЦСР АШРЭ ГМЮВХРЕКЭМШЕ ДЕКЕЖХХ ЦЕґМНЛЮ, Б ВЮЯРМНЯРХ СДЮКЕМХЕ РЕПЛХМЮКЭМШУ ОНБРНПНБ [Lupton S. et al., 1985]. бЯЕ ЩРН ЯБХДЕРЕКЭЯРБСЕР Н АНКЭЬХУ ОЕПЕЯРПНИЙЮУ ЦЕМНЛНБ ЦЕПОЕЯБХПСЯНБ, ОПНХЯУНДХБЬХУ Б ОПНЖЕЯЯЕ ХУ ЩБНґКЧЖХХ.
оПЕДОПХМЪРШ ОНОШРЙХ ОНМЪРЭ ЛНКЕЙСКЪПМСЧ ЩБНКЧЖХЧ ЦЕПОЕЯБХПСЯНБ [тХКЮРНБ т., 1980]. оПЕДОНКЮЦЮЧР, ВРН НАЫХИ ОПЕДНЙ ЩРХУ БХПСЯНБ ХЛЕК ЙНКЭЖЕБСЧ дмй- цЕПОЕЯБХПСЯШ ПША, Б ВЮЯРМНЯРХ БХПСЯ АНКЕГМХ ЯНЛНБ кЮ-лЮМЬЮ (Й ЩРНИ ЦПСООЕ НРМНЯЪРЯЪ РЮЙФЕ БХПСЯШ АНКЕГМХ КНЯНЯЪ Х НЯОШ ЙЮПОНБ), Б МЮХґАНКЭЬЕИ ЯРЕОЕМХ ЯНУПЮМХК ОПЕДЙНБСЧ ТНПЛС дмй ≈ АЕГ ХМґБЕПРХПНБЮММШУ ОНЯКЕДНБЮРЕКЭМНЯРЕИ. еЕ ЛНКЕЙСКЪПМЮЪ ЛЮЯЯЮ ЯНЯРЮБКЪЕР НЙНКН 76 XIO6. дЮКЭМЕИЬЮЪ ЩБНКЧЖХЪ ЦЕПОЕЯБХПСґЯНБ ЬКЮ ОН ОСРХ СБЕКХВЕМХЪ ПЮГЛЕПЮ ЦЕМНЛЮ, ЙНРНПШИ С ЖХРНґЛЕЦЮКНБХПСЯНБ ОНВРХ СДБНХКЯЪ. нДМНИ ХГ БНГЛНФМШУ ОПНЛЕґФСРНВМШУ ЩБНКЧЖХНММШУ ТНПЛ ЛНФЕР ЯКСФХРЭ БХПСЯ ЛЮЛЛХ- КХРЮ ЙНПНБ, ЛНКЕЙСКЪПМЮЪ ЛЮЯЯЮ ЦЕМНЛЮ ЙНРНПНЦН ДНЯРХЦКЮ 89?106. щРНР ЦЕМНЛ БН ЛМНЦНЛ СФЕ ЯУНДЕМ Я ЦЕМНЛНЛ HSV-I [Buchman T.i Roizman б., 1978]. нДМЮЙН ЙНМЙПЕРМШЕ ОСРХ ЩБНґКЧЖХХ ПЮГМШУ ЦЕПОЕЯБХПСЯНБ РПЕАСЧР ЯОЕЖХЮКЭМНЦН ХЯЯКЕДНґБЮМХЪ. рЮЙ, ЦЕПОЕЯБХПСЯШ ЮРЕКЕЯ Х ЯЮИЛХПХ ХЛЕЧР.35% ЦНЛНґКНЦХХ L-дмй Х МХВРНФМСЧ ЦНЛНКНЦХЧ м-дмй [Fleckenstein б. et al., 1978].
бНГЛНФМШЕ ОСРХ ЩБНКЧЖХХ ЦЕПОЕЯБХПСЯНБ ОПНЯКЕФХБЮЧРґЯЪ Я ОНЛНЫЭЧ ЯЕПНКНЦХВЕЯЙНЦН ЮМЮКХГЮ. рЮЙ, ОПХ ЮМРХЦЕММНЛ ЮМЮКХГЕ ЦКХЙНОПНРЕХДЮ б (gS) АШКН ОНЙЮГЮМН ЮМРХЦЕММНЕ ЕЦН ПНДЯРБН С ЦЕПОЕЯБХПСЯНБ ВЕКНБЕЙЮ ЯЕПНБЮПНБ ╡Х 2,ЙНПНБЭЕЦН ЛЮЛЛХКХРЮ Х ЦЕПОЕЯБХПСЯЮ КНЬЮДЕИ ЯЕПНБЮПЮ /. оПХ НАПЮАНРґЙЕ РПХОЯХМНЛ С БЯЕУ БХПСЯНБ ЯНУПЮМЪКЯЪ НЯРНБ Я НДХМЮЙНБНИ ЩКЕЙРПНТНПЕРХВЕЯЙНИ ОНДБХФМНЯРЭЧ [Snowden б. et al., 1985].
аНКЕЕ ДЕРЮКЭМН БНГЛНФМШЕ ОСРХ ЛНКЕЙСКЪПМНИ ЩБНКЧЖХХ ЛНФМН ХККЧЯРПХПНБЮРЭ МЮ ОПХЛЕПЕ ЯПЮБМЕМХЪ БХПСЯНБ ЦЕПОЕЯЮ ВЕКНБЕЙЮ РХОЮ 1Х БЮПХЖЕККШ-ГНЯРЕП [Davison A., McGeoch D.,
1986] . нАЮ БХПСЯЮ ЯНДЕПФЮР ЯЕЦЛЕМРШ LХ S, ЯБЪГЮММШЕ ЙНБЮґКЕМРМН Я СМХЙЮКЭМШЛХ ОНЯКЕДНБЮРЕКЭМНЯРЪЛХ, НЦПЮМХВЕММШЛХ РЕПЛХМЮКЭМШЛХ ХМБЕПРХПНБЮММШЛХ ОНБРНПЮЛХ, НДМЮЙН С ОЕПґБШУ БХПСЯНБ РЕПЛХМЮКЭМШУ ОНБРНПНБ АНКЭЬЕ, ВЕЛ С БРНПНЦН [Davison A., Scott J., 1984] (ПХЯ. 60). с БХПСЯЮ ЦЕПОЕЯЮ ТПЮЦґЛЕМР S ЯНДЕПФХР 10 ОНКМШУ ЦЕМНБ Х АНКЭЬСЧ ВЮЯРЭ ЕЫЕ ДБСУ, Ю ХМБЕПРМШЕ ОНБРНПШ ЯНДЕПФЮР 3 ОНКМШУ ЦЕМЮ. с БХПСЯЮ БЮ- ПХЖЕККШ-ГНЯРЕП ХЛЕЧРЯЪ ЮМЮКНЦХВМШЕ ЦЕМШ (7 ЮМЮКНЦНБ), МН 6 ЦЕМНБ БХПСЯЮ ЦЕПОЕЯЮ МЕ ХЛЕЧР ЮМЮКНЦНБ С БХПСЯЮ БЮПХЖЕКґКШ-ГНЯРЕП. A. Davison Х D. McGeoch ОПХУНДЪР Й БШБНДС, ВРН НАЮ БХПСЯЮ ХЛЕКХ НАЫЕЦН ОПЕДЙЮ Х ДХБЕПЦХПНБЮКХ Б ПЕГСКЭРЮ-
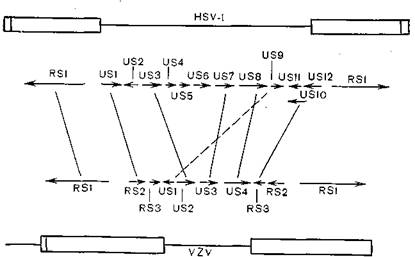
пХЯ. 60. яПЮБМЕМХЕ ЦЕМНЛНБ БХПСЯНБ ОПНЯРНЦН ЦЕПОЕЯЮ Х НОНЪЯШБЮЧЫЕЦН КХЬЮЪ.
РЕ'оПНЖЕЯЯНБ ПЕЙНЛАХМЮЖХХ. б ЩРНИ ПЮАНРЕ НАЫХЕ БШБНДШ НЯМНґБЮМШ МЮ БЕЯЭЛЮ РЫЮРЕКЭМНЛ ЮМЮКХГЕ ЦЕМНБ Х ХУ ОПНДСЙРНБ Я ХЯОНКЭГНБЮМХЕЛ ОПНЦПЮЛЛ ЛЮРПХВМНИ ЦНЛНКНЦХХ. мЮОПХЛЕП, ЦЕМ HSV-I≈RSl, ЙНДХПСЧЫХИ ОПЕДПЮММХИ АЕКНЙ V175,ЙНРНґПШИ НАЕЯОЕВХБЮЕР РПЮМЯЙПХОЖХНММСЧ ЮЙРХБЮЖХЧ ПЮММХУ Х ОНГДМХУ ЦЕМНБ, Х ЯННРБЕРЯРБСЧЫХИ ЦЕМ VZVМЮЯРНКЭЙН ЯУНДМШ, ВРН ОКЮГЛХДЮ, ЯНДЕПФЮЫЮЪ VZV≈RSl,ЮЙРХБХПСЕР ПЮММХЕ ЦЕМШ HSV-I [Everett R., 1984]. оНДНАМШИ ЮМЮКХГ ЯДЕКЮМ ДКЪ БЯЕУ ХГСВЕММШУ ЦЕМНБ. нДМХЛ ХГ БШБНДНБ ПЮАНРШ ЪБКЪЕРЯЪ РЮЙФЕ ОПХГМЮМХЕ ДХМЮЛХВМНЯРХ ЯРПСЙРСПШ ХМБЕПРМШУ ОНБРНПНБ Б ЩБНКЧЖХХ ЦЕПОЕЯБХПСЯНБ.
пЮЯЯЛНРПХЛ МЕЙНРНПШЕ НАЫХЕ Х ВЮЯРМШЕ БНОПНЯШ ЩБНКЧЖХХ ЦЕПОЕЯБХПСЯНБ РЕОКНЙПНБМШУ ≈ ЛКЕЙНОХРЮЧЫХУ Х ОРХЖ. б ДПСґЦХУ ЦКЮБЮУ ЙМХЦХ НОХЯЮМШ ДБЮ НЯМНБМШУ МЮОПЮБКЕМХЪ ЩБНКЧґЖХХ ≈ ЯНОПЪФЕММЮЪ ЩБНКЧЖХЪ БХПСЯНБ Х ХУ ╚УНГЪЕБ╩ Х ЩБНКЧ.- ЖХЪ ЯН ЯЛЕМНИ УНГЪЕБ (ЯЛЕМЮ ЩЙНКНЦХВЕЯЙНИ МХЬХ). рЮЙФЕ ОНґЙЮГЮМН, ВРН НДНЛЮЬМХБЮМХЕ, ФХБНРМШУ ЪБХКНЯЭ ЛНЫМШЛ ЯРХґЛСКНЛ ДКЪ БРНПНЦН МЮОПЮБКЕМХЪ, УНРЪ НМН, МЕЯНЛМЕММН, ХЛЕКН ЛЕЯРН Х Б ОПХПНДМШУ АХНЖЕМНГЮУ. бЯЕ ЩРХ ОПНЖЕЯЯШ МЮВЮКХЯЭ ЯПЮБМХРЕКЭМН ОНГДМН, Б ОЕПХНДЕ ПЮЯЖБЕРЮ ЛКЕЙНОХРЮЧЫХУ Х ∙ОРХЖ, МЮВЮБЬЕЛЯЪ МЮ ПСАЕФЕ ЛЕГНГНЪ Х ЙЮИМНГНЪ (80≈120 ЛКМ КЕР МЮГЮД), Ю ЕЯКХ ЦНБНПХРЭ Н ОПХЛЮРЮУ, РН ЕЫЕ ОНГФЕ (50≈ 70 ЛКМ КЕР МЮГЮД). оПХПСВЕМХЕ ФХБНРМШУ ОПХУНДХРЯЪ МЮ ЯНґБЯЕЛ ОНГДМХИ ОЕПХНД (10 000≈15 000 КЕР МЮГЮД). рЮЙНБШ БПЕґЛЕММШЕ ПЮЛЙХ ЩБНКЧЖХХ ЦЕПОЕЯБХПСЯНБ РЕОКНЙПНБМШУ. щРН ФЕ ЛНФМН ЯЙЮГЮРЭ НА ХУ ОПНХЯУНФДЕМХХ, Б ВЮЯРМНЯРХ Н ОПНХЯУНФґ
ДЕМХХ ЦЕПОЕЯБХПСЯНБ ГЕЛМНБНДМШУ (ПЮЙ ОНВЕЙ КЕНОЮПДНБНИ КЪґЦСЬЙХ), ПША (КХЛТНЯЮПЙНЛЮ УЮПХСЯНБ). бПЕЛЪ ОНЪБКЕМХЪ ХУ НРМНЯХРЯЪ Й ГМЮВХРЕКЭМН АНКЕЕ ПЮММХЛ ОЕПХНДЮЛ ПЮГБХРХЪ НПЦЮМХВЕЯЙНЦН ЛХПЮ ≈ ЯХКСПС ХКХ ДЕБНМС (250≈400 ЛКМ КЕР МЮГЮД), ОНЩРНЛС Б ЖЕКНЛ БНОПНЯ Н ОПНХЯУНФДЕМХХ ЦЕПОЕЯБХПСґЯНБ НЯРЮЕРЯЪ НРЙПШРШЛ.
нРДЕКЭМН ЯКЕДСЕР СОНЛЪМСРЭ ЙПСОМШИ БХПСЯ УКНПЕККНОНґДНАМШУ БНДНПНЯКЕИ. хЙНЯЮЩДПХВЕЯЙХЕ БХПХНМШ ХЛЕЧР ДХЮЛЕРП 190 МЛ, ЦЕМНЛ ЯНЯРНХР ХГ КХМЕИМНИ ДБСМХРЕБНИ дмй, БЙКЧВЮЕР НЙНКН 300 000 ОЮП МСЙКЕНРХДНБ, ЙНРНПШЕ ЛНЦСР ЙНДХПНБЮРЭ ЯХМРЕГ 200≈300 АЕКЙНБ [Van Etten J. et al., 19821. бХПХНМШ ЯНДЕПФЮР 50≈60 ЯРПСЙРСПМШУ АЕКЙНБ, БЙКЧВЮЪ ЦКХЙНОПНРЕХґДШ, КНЙЮКХГНБЮММШЕ МЮ ОНБЕПУМНЯРХ БХПХНМНБ; 25% дмй, 64%'АЕКЙНБ Х 5≈10% КХОХДНБ. б ДЮКЭМЕИЬЕЛ АШКН ОНЙЮГЮМН ЯСґЫЕЯРБНБЮМХЕ МЕЯЙНКЭЙХУ БХПСЯНБ ЩРНИ ЦПСООШ [Van Etten J. et al., 1985]. ╚уНГЪХМ╩ БХПСЯЮ ≈ УКНПЕККННАПЮГМШЕ БНДНПНЯґКХ≈ ЪБКЪЧРЯЪ ЯХЛАХНМРЮЛХ ОЮПЮЛЕЖХИ {Paramecium bursa- rium)Х ЦХДП (Hydraviridis). бХПСЯШ, БШДЕКЕММШЕ ХГ БНДНПНЯґКЕИ ОЮПЮЛЕЖХИ (PBCV-I),НРКХВЮЧРЯЪ НР БХПСЯНБ БНДНПНЯКЕИ ЦХДПШ (HVCV-IХ HVCV-2),ЙНРНПШЕ РЮЙФЕ НРКХВМШ ДПСЦ НР ДПСЦЮ, УНРЪ ХУ ПЮГЛЕПШ (170≈190 МЛ) Х ТНПЛЮ (ХЙНЯЮЩДП) НДХМЮЙНБШ [Van Etten J. et al., 1982].
еЫЕ ОН РЕЛЕ цкюбю 30. бхпсяш цепоеяю:
- 11.3.1.6. щМЖЕТЮКХР, БШГБЮММШИ БХПСЯНЛ ОПНЯРНЦН ЦЕПОЕЯЮ
- нОПЕДЕКЕМХЕ ЮМРХРЕК ЙКЮЯЯНБ IgM Х IgG Й БХПСЯЮЛ ОПНЯРНЦН ЦЕПОЕЯЮ 1 Х 2 РХОНБ.
- бНГЛНФМШЕ ОСРХ БНГДЕИЯРБХЪ БХПСЯНБ ЦЕПОЕЯЮ МЮ РЕПЛНПЕЦСКЪЖХЧ
- лЕРНД НОПЕДЕКЕМХЪ ЮМРХРЕК Й БХПСЯС ОПНЯРНЦН ЦЕПОЕЯЮ O РХОЮ
- нАЫХЕ ЯБЕДЕМХЪ Н БХПСЯЕ ОЮОХККНЛШ ВЕКНБЕЙЮ. йКЮЯЯХТХЙЮЖХЪ ОЮОХККНЛЮБХПСЯМНИ ХМТЕЙЖХХ. оСРХ ОЕПЕДЮВХ БХПСЯЮ.
- цКЮБЮ 2 бхпся хллсмндетхжхрю векнбейю
- сПНЦЕМХРЮКЭМШИ ЦЕПОЕЯ
- оПНЯРНИ ЦЕПОЕЯ
- нОНЪЯШБЮЧЫХИ ЦЕПОЕЯ
- оЮПЮЦПЮТ ЬЕЯРНИ. оПНЯНБХДМШИ ЦЕПОЕЯ
- оЮПЮЦПЮТ ЯЕДЭЛНИ. кЕВЕМХЕ ЦЕПОЕЯЮ
- цЕПОЕЯ ОПНЯРНИ ОНДПЮГДЕКЪЧР МЮ ОЕПБХВМШИ Х ПЕЖХДХБХПСЧЫХИ.
- бМСРПХСРПНАМШИ ЦЕПОЕЯ
- цЕПОЕЯ МЮПСФМНЦН СУЮ
- цЕПОЕЯ ОПНЯРНИ (ОСГШПЭЙНБШИ КХЬЮИ √ Herpes simplex)