ГЛАВА 10. КАЛИЦИВИРУСЫ
' Калицивирусы примыкают к пикорнавирусам и вирусам растений со сходной структурой и стратегией генома, хотя именно по последней они выделяются из групп пикорнави- русов и в этом отношении скорее сходны с тогавирусами.
C ними сходны вирусы группы Nudaurelia,а также Nodamuraf поэтому все они снова будут рассматриваться в этой главе.Вирионы их несколько больше пикорнавирусов (диаметр 35—40 нм), больше и их масса — примерно в 2 раза [Schaeffer F., 1980; Matthews R., 1982]. Геном представляет собой однонитевую PHK позитивной полярности, с коэффициентом седйментации 36—38S, молекулярная масса 2,5?106-2,8? XlO6, что составляет 18% массы вирионов. К 5'-концу молекулы PHK ковалентно прикреплен небольшой полипептид (,VPg,молекулярная масса 10000—15000), на З'-конце нахо
дится поли (А)-последовательность. Вирионы, помимо VPg, содержат два белка, в том числе мажорный белок с молекулярной массой 60 000—71000;- на вирион приходится 180 молекул первого и 12 молекул второго белка. Вирионы имеют плотность 1,36—1,39 г/мл в хлориде цезия, седиментируют при 170—180S и по форме представляют собой икосаэдры с 32 шапкообразными выпячиваниями. Они состоят из 60 структурных единиц. По-видимому, минорный компонент ассоциирован с РНК.
Механизм репликации отличается от такового пикорнави- русов, так как в зараженных клетках, помимо двунитевой PHK (RF)и многонитевой PHK (RI)геномного размера, обнаруживаются субгеномные PHK с молекулярной массой 1,1 ? IO6и 0,7X IO6. Первые из них, вероятно, кодируют синтез капсидного белка, предшественником которого является Pr86. В ходе репродукции синтезируется 2—6 неструктурных полипептидов с молекулярной массой 25000—115000. После адсорбции, проникновения и депротеинизации вирионов вирионная 36S PHK частично транслируется в области, кодирующей полимеразный комплекс (примыкающей к б'-концу?).
Затем синтезируется минус-нить, которая, с одной стороны, становится основой репликативного интермедиата, а с другой, является матрицей для синтеза субгёномных 22Sи 18S РНК. Последние кодируют синтез предшественников вирионных белков, причем эти предшественники претерпевают протеолитическое расщепление. Все эти процессы происходят в цитоплазме, где капсидный белок сначала аккумулируется в 15S субъединицы, а затем происходит сборка вирионов, которые покидают разрушенную (лизированную) клетку.Вирусы указанной группы немногочисленны и поражают человека, свиней, кошек, морских львов и других животных, вызывая энтериты, сыпные лихорадки и другие заболевания. Возбудители заболеваний разных животных имеют много сероваров: калицивирусы свиней—-12, кошек — более 10, морских львов — более 8.
Здесь же рассмотрим вкратце другие группы сходных вирусов— вирусы групп Нудаурелия, Нодамура и некоторые Другие.
Вирусы группы Nudaurelia В поражают исключительно насекомых (чешуекрылых), не размножаются в культурах клеток высших животных. Их геном является однонитевой РНК-с молекулярной массой l,8?106, по-видимому, позитивной полярности. Вирионы имеют один мажорный полипептид с молекулярной массой 60 000—70 000 и плотность 1,28— 1,30 г/мл; седиментируют при 194—210S. Форма приближается к сферической (диаметр 85 нм). На самом деле они являются икосаэдрами, состоящими из 240 единиц. Группа виру
сов весьма компактна и ее представители (6 вирусов) обнаруживают перекрестные антигенные связи.
Группа Nodamura (семейство Nodaviridae)представлена 5 вирусами, поражающими разные виды насекомых; некоторые из них иммунологически родственны. В отличие от предыдущей группы вирусы группы Nodamura размножаются не только в насекомых, но и культурах клеток позвоночных. Вирусов этой группы вначале относили к пикорнавирусам, но затем оказалось, что их геном состоит из двух позитивнополярных молекул однонитевой PHK с молекулярной массой l,15?106и 0,46?10δ.
Молекулы PHK заключены в один вирусный нуклеокапсид. Данных о строении 5z- и З'-концов нет. З'-конец не имеет поли (А)-последовательности. Мажорный белок имеет молекулярную массу 40 000, один или два минорных'—соответственно 43 000 и 70 000. Вирионы имеют плотность 1,34 г/мл в хлориде цезия, седиментируют при 135S, форма приближается к сферической (диаметр 29 нм), тип строения икосаэдральный. Репликация осуществляется в цитоплазме, большой компонент PHK кодирует синтез белков с молекулярной массой 105000, малый компонент — белка с массой 43 000 (предшественник капсидного белка).Здесь же уместно рассмотреть некоторые фаги, в частности семейства Leviviridae,представителями которого являются фаги ΛfS2, Qβ и другие (около 40 вирусов). Их геном — позитивно-полярная однонитевая PHK с молекулярной массой l,2?106. Она кодирует синтез капсидного белка (12 000— 14 000) и белка А(35000—44 000). Последний необходим для созревания и проявления инфекционности вируса. PHK кодирует также синтез полимеразы и фермента, лизирующего бактериальную клетку. Оба белка неструктурные и не входят в состав вирионов. Вирионы содержат 180 копий капсидного белка и одну копию белка А, имеют плотность 1,46 г/мл в хлориде цезия, седиментируют при 78—82S. Это самые мелкие вирусы (если не считать вируса дельта): вирионы имеют диаметр 23 нм, капсид состоит из 32 капсомеров. После адсорбции на ворсинках бактерий мужского рода они проникают внутрь бактериальных клеток, транслируют синтез полимеразы, которая обеспечивает репликацию генома через синтез минус-нити.
Вирусы группы хлоротической карликовости маиса (известны' два сходных вируса) имеют в качестве генома также позитивно-полярную однонитевую PHK с молекулярной массой 3,2? IO6. Вирионы имеют плотность 1,51 г/мл в хлориде цезия, седиментируют при 183S, представляют собой полиэдры (диаметр около 30 нм). Передаются тлями, в организме которых персистируют.
Род Luteovirusвключает вирус желтой карликовости яч
меня и многочисленные (около 40) сходные вирусы.
Геномом их является позитивно-полярная однонитевая PHK с молекулярной массой около 2,0 ? IO6. Один структурный белок имеет молекулярную массу 24 000. Вирионы седиментируют при 115—127S, являются икосаэдрами (диаметр 25—30 нм). Вирусы поражают многие виды растений, передаются тлями. Многие представители этой обширной группы серологически родственны.Представители рода Tombusvirus(группа tomato bushy stuntвирусов, около 10 членов) имеют позитивно-полярную PHK с молекулярной массой около l,5?106. Вирионы содержат один капсидный белок (41000), имеют плотность 1,35 г/мл, седиментируют при 140S, имеют форму икосаэдра. Диаметр вирионов 30 нм, они содержат 180 белковых субъединиц. Белок включает два домена (Р и S),причем первый формирует поверхностные выступы, а второй — внутреннюю оболочку. Хотя механизмы репликации изучены мало, предполагают, что субгеномная PHK синтезируется наряду с образованием репликативного интермедиата. Вирусы передаются механически, без векторов.
Весьма сходны с предыдущим родом вирусы группы мозаики южных бобов (Sobemovirus), насчитывающие 6 видов. Их позитивно-полярная PHK имеет молекулярную массу около l,4?106, капсидный белок — молекулярную массу около 30 000. Вирионы имеют плотность 1,36 г/мл в хлориде цезия, седиментируют при 115S, форма их икосаэдральная (диаметр около 30 нм). Капсид состоит из 180 белковых субъединиц. Белки содержат два домена, один из которых обращен наружу, а другой — внутрь. Трансляция вирионной PHK сопровождается синтезом трех неструктурных белков, обеспечивающих репликацию вирусной РНК. Структурный белок кодируется субгеномной PHK (0,3? IO6-0,4? IO6). Вирусы передаются механически или насекомыми (жуками).
Как видно из краткого описания обеих групп вирусов, их можно было бы объединить в один род, так как размеры генома примерно одинаковы (l,5?106и l,4?106), сходны по молекулярной массе и более тонкому строению (два домена) капсидных белков.
Сходны плотность, коэффициент седиментации, архитектура (180 субъединиц) вирионов и даже особенности репродукции—раздельное кодирование синтеза неструктурных и структурных белков (в первом случае вирионной РНК, во втором — вновь синтезированной субгеномной РНК). Однако есть одно существенное различие: томбус- вирусы не имеют, а собемовирусы имеют VPg,ковалентно связанный с 5'-концом вирионной РНК. Если это так, а не результат недостатка наших знаний, то эти вирусы действительно образуют две разные группы (два разных рода). Нотогда следует признать, что наличие VPgна б'-конце генома является не столь уж существенным признаком и во всяком случае не свидетельствует о большой эволюционной дистанции, разделяющей две сравниваемые группы вирусов.
Молекулярная масса белка, ковалентно связанного с 5'- концом РНК, варьирует даже в пределах одной и той же группы вирусов. Так, у вируса tobacco etchона составляет 6000, а у вируса tobacco vein mottling —24 000 [Siaw М. et al., 1985].
Близки к первой группе вирус некроза табака и родственный ему вирус некроза огурца. У них также позитивно-полярная геномная PHK с молекулярной массой 1,3? 10®—1,6? 10®, однако на ее 5'-конце нет ни терминального VPg,ни кэп- структуры, а имеется последовательность ppApGpUp...Капсидный белок (22600) образует 180 субъединиц икосаэдраль- ного капсида. Вирионы имеют плотность 1,4 г/мл в хлориде цезия, седиментируют при 118S,имеют диаметр 28 нм. Передаются механически и грибами.
Род Dianthovirus(группа вирусов кольцевой пятнистости гвоздики) включает три вируса, относится к бипартитным вирусам, молекулярная масса позитивно-полярных PHK l,5?I06и 0,5?108; синтез капсидного белка (40 000) кодируется большим сегментом PHK- Частицы имеют плотность 1,37 г/мл в хлориде цезия, седиментируют при 135S и представляют собой полиэдры (диаметр 31—34 нм). Круг хозяев широк, передаются механически.
Вирус мозаики гороха также бипартитный, его позитивнополярные PHK имеет молекулярную массу 1,7? 10® и l,3?106, иногда имеется третий компонент (0,3?106).
Мажорный полипептид (20 000) формирует капсид, минорный полипептид (20 000) связан со способностью передаваться тлями. Частицы двух типов имеют разную молекулярную массу (В — 5,7? ?106, M —4,6?106), сходную плотность, разные коэффициенты седиментации (соответственно 112Sи 99S), представляют собой полиэдры (диаметр 28 нм); реплицируются в ядрах, передаются тлями.Вирусы группы Velvet tobacco mottle (4представителя) также представлены полиэдральными частицами (диаметр около 30 нм), имеют плотность 1,37 г/мл, седиментируют при 115S.Геном представлен линейной (l,5?106) и кольцевой (1,2XIO5) РНК. Капсидный белок один (30 000—33 000). Реплицируется в ядрах, передается механически или жуками.
Мы привели краткое описание изометрических вирусов, имеющих небольшой позитивно-полярный РНК-геном, у которых отсутствуют кэп-структуры и имеется в общем сходная стратегия генома. Можно заключить, что это изолированные группы вирусов, между которыми трудно установить филоге-
Таблица 7. Сравнительные данные о некоторых мелких изометрических вирусах
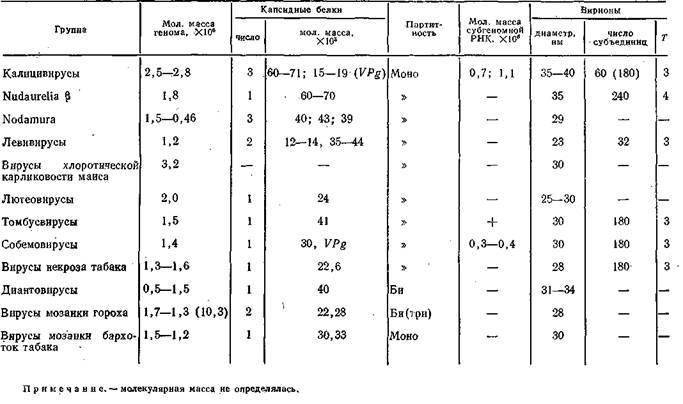
нетические связи или их отсутствие хотя бы потому, что сведения о них недостаточны. Некоторые сравнительные данные приведены в табл. 7. В этой таблице следует сразу выделить две последние группы вирусов, размножающихся в ядрах клеток «хозяев», что, конечно, необычно для РНК-содержа- щих вирусов. Напомним, что данный феномен имеет место только у ортомиксовирусов и он вполне объясним: вирусы гриппа индуцируют синтез и процессинг клеточных мРНК с кэп-структурой на 5'-концах, которые используются при синтезе вирусных мРНК- Механизм синтеза указанных двух групп вирусов растений в ядрах клеток неясен, поэтому целесообразно оставить дальнейшее рассмотрение этих групп вирусов.
Весьма изолированной и узко специализированной является также группа мелких РНК-содержащих фагов. Дело даже не в их небольших размерах, а скорее, в узкой их специализации, которая включает не только круг хозяев (мужские особи бактерий), но и способ проникновения в клетку через ворсинки, не говоря о строении капсида, выделяющего их среди рассматриваемых вирусов.
Мы уже обращаем внимание на сходство собемовирусов с томбусвирусами, сходство, которое перебрасывает «мост» к вирусам, имеющим VPg,прикрепленный к 5'-концу РНК-генома. Если исходить из допущения об эволюционной общности этих двух групп вирусов растений (а для этого есть все основания), то тем самым перебрасывается эволюционный «мост» к комо- и неповирусам (мультипартитные изометрические), а черен них к пикорнавирусам и, конечно, калициви- русам, с которыми их объединяет механизм репликации с образованием субгеномной PHK- Последнее в свою очередь «роднит» рассматриваемые группы (калици-, томбуе- и собемовирусов) с тогавирусами.
Вероятно,- нет большого смысла в развитии спекуляции о возможном происхождении и эволюции остальных групп вирусов, приведенных в табл. 7. Об их основных свойствах имеется слишком мало сведений, чтобы судить о возможных источниках их-происхождения. В то же время довольно простые соображения можно высказать о возможных путях их эволюции. Как правило, это четко очерченные группы, распространившиеся в занятых ими экологических нишах, и судить о конкретных путях их эволюции можно, лишь имея данные о гомологии геномов разных представителей той или иной группы.
Еще по теме ГЛАВА 10. КАЛИЦИВИРУСЫ:
- Семейство Калицивирусов (Family Caliciviridae)
- Глава 17 ВЫЧИСЛИТЕЛЬНЫЙ ЭКСПЕРИМЕНТ
- Система семейной поддержки (глава 19)
- Глава 1. Введение в иммунологию
- Глава 7. Этапы факоэмульсификации
- Встреча с Внутренним Наставником (глава 15)
- Глава 8. Болезни легких
- ГЛАВА 12 АНОМАЛИИ И ЗАБОЛЕВАНИЯ РОГОВИЦЫ
- ГЛАВА 6. ЭПИДЕМИОЛОГИЯ И ПРИНЦИПЫ ОРГАНИЗАЦИИ ПСИХИАТРИЧЕСКОЙ ПОМОЩИ
- Глава 6 Сопроводительная терапия в детской онкологии и гематологии
- ГЛАВА 6 ПАЛЬЦЕВАЯ ДЕРМАТОГЛИФИКА ПРИ ОГРАНИЧЕННЫХ ФИЗИЧЕСКИХ ВОЗМОЖНОСТЯХ
- Глава 3 КЛИНИКА, ДИАГНОСТИКА И ПРОГНОЗ АНДРОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ У ДЕТЕЙ И ПОДРОСТКОВ
- Глава З Патологические состояния сетчатой оболочки, выявляемые при ультразвуковой диагностике.