Циклические инфекционные (моно)процессы
Генерализация поксвирусной инфекции. Антигенная детерминанта покс- вирусов. Иммунные ответы на поксвирусы. Трансмиссионный потенциал ВНО. Продолжительность противооспеннаго иммунитета.
BHO легко обнаруживается иммунной системой человека. Он индуцирует полный спектр клеточно-опосредованных и гуморальных иммунных ответных реакций. Заболевший начинает активно выделять вирус в первую неделю от начала болезни, когда происходит вскрытие оспенных элементов на слизистой ротовой полости, зева и глотки. Количество же антител, нейтрализующих инфекционность вируса, достигает своего максимума между 12-м и 15-м днями болезни. Но уже на 10-е сутки болезни зараз- ноетъ больного для окружающих резко снижается. Через 4 недели от начала болезни вирус уже невозможно выделить из отделяемого носоглотки и из мочи реконвалесцента. Выздоровление у заболевших людей полное (Ма- ренникова С. C., Щелкунов С. H., 1998). Таким образом, во взаимоотношениях BHO с организмом человека просматривается определенная стратегия паразитизма. Попробуем понять, из чего она складывается.
Генерализация поксвирусной инфекции, C 1940-х гг. наиболее адекватной моделью инфекционного процесса, вызываемого BHO у человека, считается эктромелия мышей (Mark R. L. et al., 1991; Fenner F., 1948). Возбудитель болезни (Ectromelia virus, ЕѴ) относится к семейству Orthopoxvirus. Ортопоксвирусы консервативны. Они имеют более чем 90 % гомологии в центральном 100 кДа регионе генома (Esteban D. J., Buller R. M., 2005). Сходство патогенезов болезней, вызываемых BHO и ЕѴ, включает быстрое размножение вируса при его низкой инфицирующей дозе, сходство цитокиновых ответов на вирус и сходные кожные поражения на последней стадии болезни, включающие орофарингеальную секрецию вируса, критическую для его трансмиссии (Chaudhri G. et аі., 2006). Генерализация вируса эктромелии изучена в деталях (рис.
43).Первыми клетками, вступающими в контакт с поксвирусами. являются полиморфоядерные лейкоциты (макрофаги и моноциты). За счет градиента продуцируемых ими хемоаттрактантов они «притягивают» другие фагоцитирующие клетки и запускают каскад самых разнообразных реакций, которые «в сумме» рассматриваются учеными как «иммунные». В составе макрофагов и моноцитов вирус достигает дренажных лимфатических узлов примерно через 8 ч. Разрушившиеся фагоцитирующие клетки становятся источником вируса, а через 2-3-е суток его обнаруживают уже в крови (первичная виремия), костном мозге, печени и селезенке. В пробах крови больного натуральной оспой в этот период можно обнаружить как инфицированные, так и неинфицированные лейкоциты, но в основном репликация BHO происходит в моноцитах/макрофагах, тогда как Т-лимфоциты остаются неинфицированными, но при этом наблюдается выраженная лимфопения. Вирус распространяется из лимфатических узлов фаго- цитирущими клетками крови по внутренним органам через выносящие (эфферентные) лимфатические сосуды. Процесс напоминает показанный на рис. 28 для возбудителя сибирской язвы, но, естественно, без участия бактериальных факторов вирулентности. Затем вирус проникает в эпителий кожи и слизистых, где начинается его репликация. Появляются энантемы и экзантемы, инфицирование которых вторичной микрофлорой определяет эволюцию кожных элементов из везикул в пустулы. Количество вируса через 4-5 суток уже превышает емкость ретикуло-эндотелиальной системы.
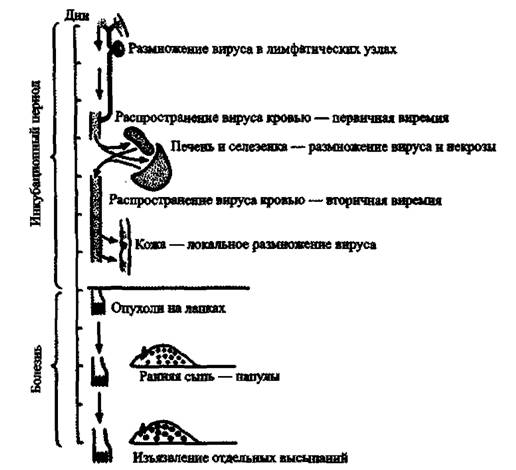
Рис. 43. Генерализация ортопоксивурусов на примере вируса жтромелии у мышей. По F. Fenner (1948)
Процесс генерализации возбудителя болезни осуществляется благодаря макрофагам. Вирус ведет себя как их паразит и быстро размножается. В отличии от ВИЧ (см. разд. 3.2) он не поддерживается в макрофагах в качестве эндосимбионта, а активно их разрушает, затем попадает в кровь, где может «встретиться» со специфическими антителами
и он вновь проникает в кровяное русло.
Вторичная виремия обычно соответствует началу клинической манифестации болезни. При всех формах эктромелии у мышей и натуральной оспы у людей развиваются паренхиматозные изменения во внутренних органах. Циркулирующие в крови специфические антитела выявляются уже на 7-е сутки после инфицирования человека ВНО, и их максимальный уровень достигается на 14-е сутки. Одновременно с нарастанием количества специфических антител происходит снижение количества вируса в тканях (более подробно об этих процессах см. в работах Esteban D. J., Buller R. M., 2005; McFadden G., 2004; Маренниковой С. C,, Щелкунова С. H., 1998; Mark R. L etal., 1991).Считается, что в основе патогенеза поксвирусных инфекций лежит способность вирусов к интерференции с защитными иммунными механизмами макроорганизма (Stanford М. et al., 2007). По крайней мере, это так выглядит, если смотреть,на процессы взаимодействия осповирусов со своими многоклеточными хозяевами с точки зрения, предполагающей их совместную эволюцию. Особое значение учеными придается антихемокиновой стратегии ортопоксвирусов, заключающейся в блокировании вирусными белками хемокиновой активности клеток иммунной системы хозяина. Но судя по древности хемокиновых белков, система взаимоотношений посредством сигнальных молекул между прокариотами (в том числе, вирусами) и эукариотами сложилась еще в архее, т. е. до образования многоклеточной жизни. Поэтому такая стратегия не оригинальна для ортопоксвирусов; в отношении фагоцитирующих клеток она используется большинством ДНК-вирусов. Стратегия включает синтез инфицированными фагоцитирующими клетками вирусных гомологов хемокинов или хемокиновых рецепторов и других белков, связывающих хемокины, синтезируемые неинфицированными фагоцитирующими клетками (рис. 44; табл. 12).
Поскольку функции хемокиновых рецепторов, посредством которых вирус взаимодействует с клеткой, различаются, то воздействие на них вирусными белками на уровне макроорганизма приводит к разным и даже противоположным эффектам. Это и истощение локальной хемокиновой активности, и индукция межклеточных сигналов, усиливающих вирусную репликацию, и стимуляция миграции инфицированных клеток либо, наоборот, нейтрализация их способности к миграции.
Но рассмотрение тонкостей антихемокиновых стратегий вирусов не входит в задачу данной работы (подробно с антихемокиновыми стратегиями ортопоксвирусов в организме млекопитающих можно ознакомиться по публикациям Stanford М. et а1„ 2007; Alcami А., 2007; Smith S. A., Kotwal G. J., 2002; Маренни- ковой С. С. и Щелкунова С. H., 1998). Тем более что с ними нет полной ясности, свидетельствующей о том, что антихемокиновая стратегия BHO в организме человека сформировалась в результате их коэволюции.Так G. McFadden (2004) обратил внимание исследователей на то, что маловирулентный ортопоксвирус, возбудитель оспоподобной болезни — аластрима (alastrim), мало уступает собственно BHO по числу генов белков — ингибиторов иммунных ответных реакций макроорганизма (см. табл. 12). Поэтому он усомнился в правильности общепринятого понимания механизмов, благодаря которым BHO вызывает смертельную болезнь В связи с его сомнениями выскажу и свое. BHO имеет большое количество «выключенных» генов вирулентности. Особенно это бросается в глаза при сравнении его генома с геномами маловирулентных ортопоксвирусов,
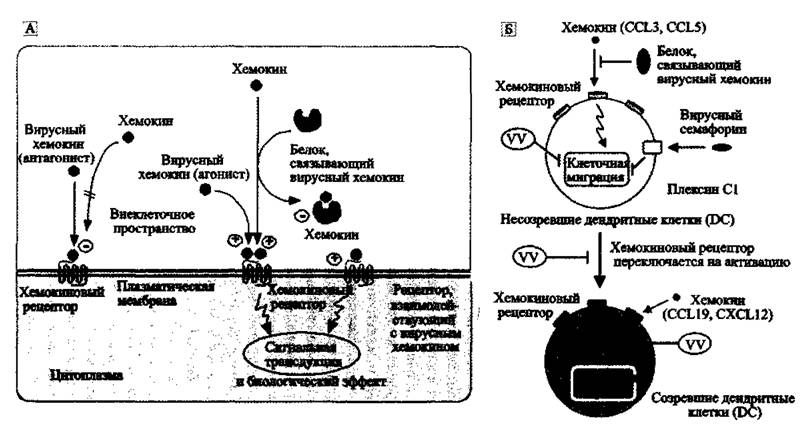
Рис, 44, Модуляция хемокиновой системы вирусами
А. Экспрессия вирусных гомологов хемокинов, рецепторов хемокинов и хемокинсвязывающих белков. Б. Частный случай антихемокиновой стратегии — ингибирование миграции созревающих дендритных клеток (DC). Вирусные хемокин- связывающие белки интерферируют с хемокинами CCL3 и CCL5. Не созревшие DC экспрессируют воспалительные хемокиновые рецепторы (CCR1, CXCR1) и мигрируют в инфицированные ткани в ответ на такие хемокины, как CCL3 и CCL5. Созревшие DC экспрессируют CCR7 и CXCR4 и мигрируют во вторичные лимфоидные органы в ответ на CCLl 9 и CXCLl 2, где они представляют антиген «наивным» Т-клеткам. По A. Alcami (2007)
Портрет серийного убийцы: нммуномодулирующне гены ВНО[4]
Таблица 12
| Иммуномодулирующее действие | Variola major[5] | Variola minor** | Комментарии |
| IFN -α/β-ингибитор | B20R | D9R | He проверено |
| IFN-y-ингибитор | B9R | H9R | Ингибирование человеческого IFN-γ |
| еІР2сс-мимикрия | C3L | D3L | Не проверено |
| dsRNA-BP | E3L | C3L | Не проверено |
| Ингибитор фактора некроза опухолей | G2R | G2R | Не проверено |
| Ингибитор N F-κΒ | PlL | PlL | Не проверено |
| TLR-Citгнальный ингибитор | A52R | A56R | Не проверено |
| IL-18-связывающий белок | D5L | B6L | Ингибитор человеческого IL-18 |
| Хемокинсвязывающие белки | G3R | A46L | G3R ингибирует у людей хемокины CC |
| Фактор роста вирусов | G3R | B3R | Связывает и активирует человеческий EGF-R (erbBl) |
| Ингибитор комшіемента/SPICE | D12L | B18L | Ингибирует каскад комплемента у людей |
| GM-CSF/IL-2-ин гибитор | A46R | A49L | Не проверено |
| Семафорин | A42R | A45R | Не проверено |
| Серпины | B13R, B25R, C2L | D14R, D2R | Не проверено |
у исходного (Senkcvich Т.
G. et аі, 1994). Недавно у белка, синтезируемого BHO и способного связывать рецептор фактора некроза опухолей, обнаружили домен, инициирующий рекрутинг макрофагов к поверхности слизистых и кожи, в месте внедрения вируса (Alejo А. et аі., 2006).Инфицирование ВИЧ всегда кончается смертью больного, количество же смертельных исходов при натуральной оспе сильно варьирует во время разных эпидемий и в разных группах населения. Поэтому интерес представляют реакции макроорганизма на заведомо смертельные дозы ВНО. Патогенез натуральной оспы при инфицировании такими дозами вируса изучали Р. В. Jahrling et аі. (2004) на обезьянах. Общей реакцией макроорганизма на проникновение вируса является рост количества фагоцитирующих клеток — моноцитов, чем позже наступает пик лейкоцитоза, тем дольше живет инфицированное животное. BHO высевается из моноцитов и смывов из носоглотки уже на вторые сутки после инфицирования, хотя еще его не удается выделить из плазмы. Через 2-3 суток у инфицированных приматов уже можно наблюдать отчетливо выраженные кожные поражения (везикулы и пустулы). Количество кожных поражений зависит от инфицирующей дозы. Вирусная репликация в макрофагах сопровождается явлением, которое из-за его неблагоприятного исхода назвали «цитокиновым штормом*- («cytokine storm»). Оно проявляется каскадной активацией и выбросом инфицированными макрофагами значительных количеств различных лимфокинов (monocyte chemoattractant protein I, macrophage inflammatory protein 1β, IFN-γ, IL-6 идр.) и развитием клиники токсемии и шока, приводящих больного к смерти (рис. 45).
Через 2-3-е суток после инфицирования BHO у животных развивалась коагулопатия, проявившаяся геморрагическими высыпаниями в грудной и брюшной полостях и в передней камере глаза. Электронная микроскопия демонстрировала репликацию вируса в большинстве тканей внутренних органов обезьян и, особенно, в лимфатических узлах и селезенке. Фибриновые отложения и тромбы ассоциировались с выраженным размножением вируса в макрофагах.
Т-лимфоцитозависимые области лимфоидной ткани обычно выглядели истощенными, тогда как макрофаги, моноциты, клетки эндотелия активно продуцировали вирус. Гибель Т-лимфоцитов происходила в результате неспецифического апоптоза (bystander apoptotic), инициированного инфицированными макрофагами. У единственного выжившего животного развился стерильный иммунитет.Антигенная детерминанта поксвирусов. Основной антигенной детерминантой поксвирусов и мишенью для протективных антител является консервативный белок Ll (другое название LIR). У вируса вакцины, вируса оспы обезьян и BHO этот белок различается отдельными аминокис-
157
3,1. Циклические инфекционные (моно)процессы
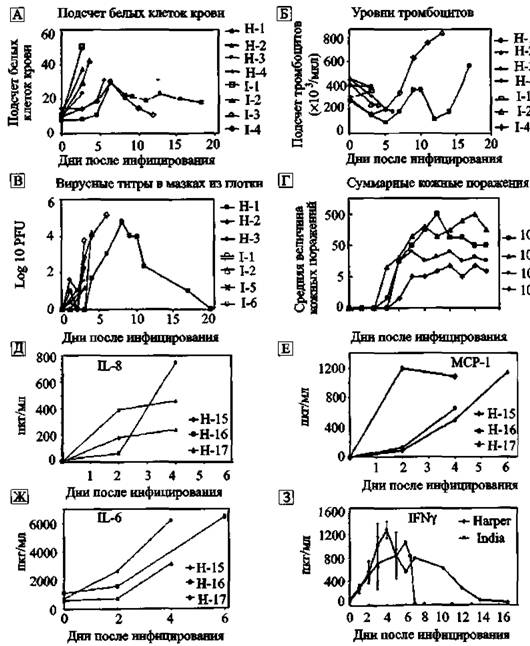
Рис. 45. Динамика патогенетических реакций обезьян на заведомо смертельные дозы BHO штаммов Harper и India
А. Общее количество белых клеток крови. Б. Общее количество тромбоцитов. В. Инфекционные титры вируса в смывах из гортани. Г. Кожные поражения как функция дозы вируса {штамм India). Д. Концентрация IL-S (пкг/мл). Е. Концентрация хемоаттрактантного белка моноцитов (MCP-I) в пкг/мл, Ж. Концентрация IL-6 в пкг/мл. 3. Среднее количество IFN-у в пкг/мл в сыворотке обезьян, инфицированных штаммами India 7124 и Harper (109 pfu). По Р. В. Jahriing et аі. (2004)
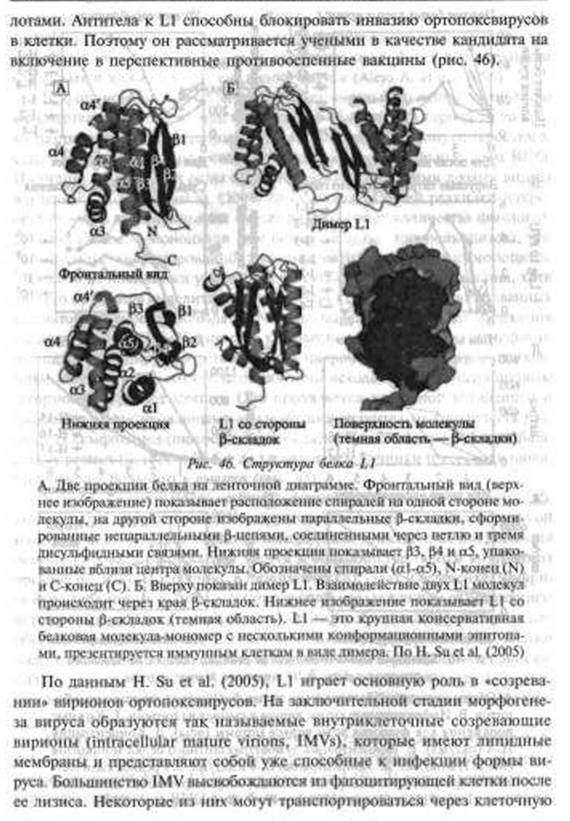

фических антител. Во-первых, антитела могут непосредственно связывать вирус, вызывая его агрегацию и препятствуя адсорбции и интернализации в клетках. Во-вторых, они могут связывать вирус, вызывая его разрушение с помощью комплемента или опсонизации, а затем посредством фагоцитоза. Антитела могут связывать инфицированные клетки хозяина, вызывая цитотоксические реакции со стороны клеток-киллеров и др.
Истощение Т-клеток CD8 (цитотоксические клепки, способные убивать другие клетки, к антигенам которых они примированы) посредством моноклональных антител или генного нокаута, сказывалось незначительно на развитии инфекционного процесса. В антивирусной функции CD8 Т-кле- ток много неясного, Spriggs et al (1992) показали, что трансгенные мыши, утратившие CDS+ Т-клетки, способны также как и мыши в контроле, пережить инфицирование большими дозами вируса эктромелии. Но у таких трансгенных мышей синтезировалось меньше вирусоспецифических IgG1 что предполагает участие активированных CD8 Т-клеток в освобождении организма от вируса. R. Xu et аі. (2004) показали способность мышей, дефектных по MHC класса Н(-/-) и инфицированных сублетальными дозами вируса вакцины, выживать и полностью освобождаться от вируса при наличии активных CD8+ Т-клеток. Их данные показывают участие CDS+ Т-клеток в снижении смертности животных, инфицированных вирусом вакцины, однако детали этого процесса пока не ясны. Мыши с таким генетическим дефектом имели значительно меньшее по сравнению с нормой количество CDS+ Т-клеток, и во время инфекции у них развивались обширные некротические повреждения в кортексе надпочечников, характерные для инфекционного процесса, вызванного вирусом вакцины.
По данным G. Chaudhri et аі. (2006), именно В-клетки играют ключевую роль в освобождении организма мыши от вируса эктромелии. При дефиците В-клеток и при наличии CD8 Т-клеточных ответов животные неизбежно погибают от прогрессирующей инфекции. Любопытно то, что в их опытах введение животным, дефицитным по В-клеткам, «наивных» В-кле- ток или специфической к вирусу эктромелии сыворотки позволяло добиться стерильного иммунитета.
О важной роли гуморального звена иммунитета в противодействии BHO свидетельствуют и старые работы. По данным А. Downie et аі. (1958), геморрагическая форма натуральной оспы с летальным исходом развивалась при отсутствии у заболевшего нейтрализующих антител после предшествующей вакцинации. У таких больных развивалась выраженная ви- ремия, вирус в высоких титрах обнаруживали в фарингеальном тракте.
Полученные данные свидетельствуют в пользу того, что ортопоксви- русные инфекции не контролируются реликтовой системой иммунитета.
Уничтожение таких вирусов осуществляется сложившейся уже в позвоночных организмах системой гуморального и клеточного иммунитета с преимущественной ролью В-лимфоцитов. Благодаря хорошей «заметности» и ранней презентации T- и В-клеткам антигенов ортопоксвирусов, одновременно являющихся критическими для их морфогенеза, специфические антитела обладают выраженным протективным действием.
Трансмиссионный потенциал ВНО. Способность BHO передаваться от человека к человеку в иммунокомпетентных популяциях весьма ограничена. По данным V. Bhatnagar et аі. (2006), обобщивших статистическую информацию по 51 вспышке «завозной» натуральной оспы, зарегистрированной в Европе и Северной Америке в период после 1945 г., в среднем каждая такая вспышка ограничивалась четырьмя случаями болезни с одним летальным исходом. Количество передач (генераций) BHO от заболевшего человека к здоровому редко превышало 3; а 30 % заносных вспышек вообще не сопровождалась передачами BHO от человека к человеку. Такую неспособность оспы к быстрому распространению в популяциях людей нельзя объяснить только предварительными массовыми вакцинациями. Например, вспышка натуральной оспы среди невакцинированного населения Abakaliki (Нигерия) в 1967 г. ограничилась всего 31 случаем болезни (Eichner M., Dietz К., 2003). В свете этих данных невозможно понять, каким образом жестокая пандемия натуральной оспы 1870-1874 гг. вспыхнула практически одновременно на нескольких континентах, если не предположить, что вирус проникал в популяции людей из каких-то синхронно активизирующихся природных очагов. В Европе натуральная оспа во время пандемии 1870-1874 гг. нс «обращала внимания» даже на двукратную противооспенную вакцинацию населения отдельных городов (детали распространения натуральной оспы в ту пандемию см. в книге Бразоля Л. E., 1875).
Продолжительность противооспенного иммунитета. Вакцинация обычно предупреждает заражение BHO в течение, по меньшей мере, 5 лет. При развитии болезни ее симптомы у вакцинированных лиц менее выражены, чем у невакцинированных. Так как специфические антитела считаются первой линией обороны против вторжения возбудителей инфекционных болезней, их обычно выявляют для оценки иммунной защиты индивидуума. Поданным S. Gallwitz et аі. (2003), использовавших ферментативный иммуноанализ для поиска антител к вирусу вакцины у лиц, вакцинированных 30-60 лет назад, их можно обнаружить у 65 % обследуемых, вакцинированных однократно; и у 80 % обследуемых, вакцинированных два раза и более. S. Crotty et аі. (2003) продемонстрировали присутствие у лиц, вакцинированных более 50 лет назад, специфических к вирусу вакцины В-клеток памяти. Количество таких клеток после вакцинации снижается
в течение нескольких лет до уровня, представляющего примерно десятую часть от достигнутого максимума. Далее количество противооспенных антител (0,1 % от общего количества IgG+ В-клеток) не меняется практически на протяжении всей оставшейся жизни вакцинированного. Этим объясняется выраженная антительная реакция на противооспенную ревакцинацию. В опытах S. Е. Frey et аі. (2003) вакцинация ранее вакцинированных лиц давала очень хороший результат даже при десятикратном разведении вакцины.
** *
В эволюционном аспекте проблемы инфекционных болезней инфекционный процесс в многоклеточных организмах представляет собой попытку микроорганизмов установить с фагоцитирующими клетками взаимоотношения, существовавшие между ними до формирования многоклеточной жизни. Такие взаимоотношения могут развиваться сразу как паразитические (оротопоксвирусы, возбудители чумы или сибирской язвы) либо начинаться с симбиоза (возбудители бруцеллеза, туберкулеза, проказы). В медицинском аспекте проблемы инфекционный процесс — это процесс диссеминации фагоцитирующими клетками микроорганизмов по органам и тканям, сопровождающийся выбросом молекул межклеточного общения (хемокинов и лимфокинов) в количествах, значительно превышающих физиологическую норму, и, соответственно, проявляющийся развитием патологических реакций (т. е. болезни). Его продолжительность лимитируется T- и В-клеточной составляющей иммунной системой хозяина, сформировавшейся уже в процессе эволюции позвоночных организмов, При рассмотрении инфекционного процесса данного типа речь идет не о конкретном количестве суток, в течение которых начинается и прекращается инфекционный процесс, а о способности клеточной и гуморальной иммунной системы его контролировать. Например, при туберкулезе средний инкубационный период составляет 5-10 недель.
У иммунокомпетентного человека клеточный иммунный ответ на гематогенную диссеминацию возбудителя болезни возникает через 2-10 недель, что останавливает его размножение. У переболевших туберкулезом людей остается относительная невосприимчивость к повторному заражению туберкулезной палочкой.
Для инфекционной болезни, развивающейся вследствие инфекционного процесса, лимитируемого клеточной и гуморальной иммунной системой, характерны следующие периоды: инкубационный, продромальный, нарастания симптомов, разгара болезни, угасания клинических проявлений болезни, выздоровления (реконвалесценции) с формированием стерильного иммунитета. Такие инфекционные процессы я предлагаю назвать циклическими инфекционными процессами. Обычно они представляют собой монопроцессы, т. е. вызываются одним микроорганизмом.
На примере инфекционных процессов, вызванных BHO и другими отопоксвирусами, мы видим, что до достижения гуморальной и клеточной иммунной системой определенного максимума реакции строгий антропоноз {зооноз) должен сменить своего хозяина, чтобы сохраниться как биологический вид. В нашем восприятии времени BHO меняет хозяина на 8-10-е сутки от начала болезни. Та патогенность (вирулентность) микроорганизма, которая характеризуется в нашем восприятии времени быстрым инкубационным периодом и непродолжительной болезнью, завершающейся либо гибелью организма, либо его выздоровлением, — это проявление определенной стратегии паразитизма микроорганизма в многоклеточном хозяине. Условно назовем ее первой стратегией паразитизма, или стратегией мора. Антропонозному (зоонозному) паразиту стратегия первого типа дает преимущества в высокоплогных популяциях хозяина.
3.2.
Еще по теме Циклические инфекционные (моно)процессы:
- Брюшной тиф
- Псевдотуберкулез
- Малярия
- 12.1. Рассеянный склероз
- Болезни костей, суставов и опухоли мягких тканей
- Глава 18. РЕВМАТИЧЕСКИЕБОЛЕЗНИ. ЭНДОКАРДИТЫ
- Циклические инфекционные (моно)процессы
- СОДЕРЖАНИЕ