Строение и физиологические функции а-синуклеина
Интерес к свойствам небольшого растворимого белка а-синуклеина резко возрос после того, как была установлена взаимосвязь между нарушением его функции и развитием ряда нейродегенеративных заболеваний, включая БП.
Многолетний патологический процесс при БП характеризуется прогрессирующей потерей ДА-ергических нейронов в компактной части черной субстанции и ряде других структур мозга и образованием сферических эозинофильных цитоплазматических включений (телец Леви) в соме нервных клеток, а также веретено- или нитеподобных ветвящихся нейритов Леви и нейроаксональных сфероидов в отростках нейронов; эти включения являются морфологическим субстратом нейродегенерации (Del Tredici, Braak, 2004). Накопление телец и нейритов Леви в нервной ткани рассматривается в качестве критического диагностического фактора БП (Eriksen et al., 2005).Главным компонентом телец и нейритов Леви являются агрегации неправильно свернутого и токсичного для клетки высококонсервативного белка а-синуклеина с молекулярной массой 19 кДа (140 аминокислот), который локализуется в основном в аксонах и их пресинаптических терминалях (Shults, 2006). Ген а-синуклеина (SNCA) расположен на длинном плече четвертой хромосомы и состоит из семи экзонов, пять из которых транскрибируются. Агрегации а-синуклеина характерны для целой группы заболеваний, которые рассматриваются как синуклеинопатии (табл.)
Таблица
Заболевания, сопровожающиеся агрегацией а-синуклеина, по (Spillantini et al., 1998)
а-синуклеинопатии
Болезнь Паркинсона Спорадические формні Наследственные формы Деменция с тельцами Леви Мультисистемная атрофия Синдром Ши-Драгера Стриатонигральная дегенерация
Оливопонтоцеребеллярная дистрофия (болезнь Дежерина-Тома)
Болезнь Галлервордена-Шпатца
Другие заболевания, при которых возможны синуклеиноиммунореактивные поражения Болезнь Альцгеймера Болезнь Пика
Амиотрофический латеральный склероз Травматические поражения мозга
Альфа-синуклеин широко распространен в мозге (Jakes et al., 1994; Iwai et al., 1995).
В полном транскрипте, а-синуклеине-140, выделяют три области (рис. 1). N-концевой участок (основания 1-65) включает сайты по крайней мере двух ауто- сомно-доминантных наследственных мутаций, вызывающих БП, - замена аланина на треонин в позиции 53 (A53T) и замена аланина на пролин в позиции 30 (A30P); он содержит шесть неполных повторов 11-аминокислотных остатков с высококонсервативным гексамерным мотивом (KTKEGV). Предполагается, что N-концевой участок способен образовывать структуру из двух а-спиралей, соединенных коротким прямолинейным участком, и типичную для липид-связывающих доменов аполипопротеинов класса A2. Было показано, что липидное окружение стимулирует сворачивание а-синуклеина, а также ускоряет его агрегацию (Lee et al., 2002). Центральный гидрофобный участок (основания 66-95) включает NAC-последовательность (non-amyloid component), склонную к агрегации. Впервые NAC-пептид был выделен из амилоидных бляшек коры головного мозга пациентов, страдавших болезнью Альцгеймера (Ueda еt al., 1993). И лишь затем было обнаружено, что а-синуклеин является основным компонентом телец Леви при БП. C-концевой участок (основания 96-140) отрицательно заряжен за счет кислых аминокислот. В этом регионе локализованы несколько сайтов фосфорилирования (Tyr-125, -133 и -136 и Ser-129), а также домен, отвечающий за шаперонную активность а-синуклеина (основания 125-140) (Recchia et al., 2004).Функции этого распространенного белка, количество которого в растворимой цитозольной мозговой фракции составляет не менее 1% от всего белка, остаются недостаточно выясненными. Однако его локализация в пресинаптических термина- лях (Iwai et al., 1995; Jakes et al., 1994), ассоциация с дистальным резервным пулом синаптических везикул (Zhang et al., 2008) и нарушение синаптической передачи при нокаутировании животных по а-синуклеину или при избыточной экспрессии этого белка позволяют считать, что а-синуклеин играет определенную роль в регу-
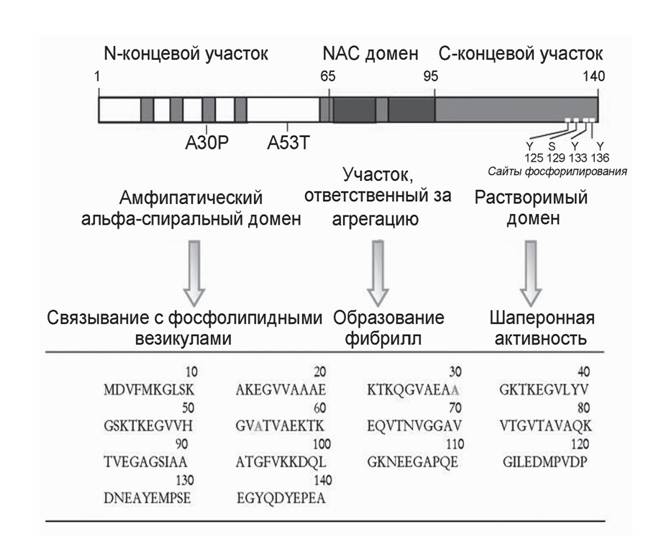
Рис.
1. Аминокислотная последовательность и домены а-синуклеина человека, по (Recchia et al.,2004) ляции высвобождения нейромедиаторов, синаптической функции и пластичности (Lashuel et al., 2012).
Анализ мышей-нокаутов по гену а-синуклеина показал, что у них заметно сокращается время, необходимое для восстановления способности к выбросу ДА в стриатуме после повторной электрической стимуляции, падает уровень ДА в ст- риатуме и наблюдается снижение локомоторного ответа на введение амфетамина (в результате уменьшения выброса ДА в синаптическую щель и инвертирования действия дофаминового транспортера на локомоторную активность) (Abeliovich et al., 2000). У мышей-нокаутов по гену Snca обнаружены нарушения гиппокампальных синаптических реакций в ответ на длительную высокочастотную стимуляцию, способную истощить готовые к выбросу и резервные пулы синаптических везикул, а также нарушения процесса пополнения готовых к выбросу пулов из резервных депо. Эти результаты могут означать, что а-синуклеин контролирует пополнение синаптических везикул и их перемещение из резервного пула к области высвобождения синаптических везикул (Cabin et al., 2002; Abeliovich et al., 2000). Кроме того, показано, что истощение а-синуклеина с помощью антисмысловых олигонуклеотидов вызывает уменьшение резервного пула синаптических везикул в первичной культуре нейронов гиппокампа (Murphy et al., 2000).
У трансгенных мышей с гиперэкспрессией человеческого а-синуклеина определены нарушения экзоцитоза синаптических везикул и уменьшение высвобождения нейромедиатора (Nemani et al., 2010). Подобные эффекты наблюдались при гиперэкспрессии а-синуклеина в генетических моделях БП на грызунах (Gaugler et al., 2012; Lundblad et al., 2012) и на клеточной линии PC12 (Larsen et al., 2006). Гиперэкспрессия а-синуклена вызывает снижение количества готовых к высвобождению везикул (Gaugler et al., 2012), рециркуляции синаптических пузырьков после эндоци- тоза, размера синаптического пула рециркулирующих везикул (Nemani et al., 2010), обратного захвата ДА в ДА-ергических терминалях (Lundblad et al., 2012).
Возможная роль а-синуклеина в регуляции синаптического гомеостаза не связана исключительно с его непосредственным взаимодействием с синаптическими пузырьками. Показано, что а-синуклеин может действовать как белок-шаперон по отношению к пресинаптическому белковому комплексу SNARE, непосредственно вовлеченному в процесс высвобождения нейромедиаторов, включая ДА (Burre et al., 2010). Совокупность данных свидетельствует, что а-синуклеин играет важную роль в перемещении синаптических везикул и регуляции везикулярного экзоцитоза и, возможно, контролирует ряд белков, связанных с синаптическим гомеостазом.
Тот факт, что нокауты по тому или иному синуклеину (а, P и у) жизнеспособны, позволяет считать, что синуклеины не являются ключевыми участниками в механизмах высвобождения нейромедиатров, но, возможно, вносят свой вклад в долгосрочную регуляцию и поддержание функций нервных терминалей (Chandra et al., 2004).
1.2.
Еще по теме Строение и физиологические функции а-синуклеина:
- Молекулярные механизмы патогенеза болезни Паркинсона и перспективы превентивной терапии
- Строение и физиологические функции а-синуклеина