1H МР-спектры мозга: отнесение сигналов, функция и метаболизм 1Н МР-видимых соединений
В 1H МР-спектрах мозга в норме (рис 1) детектируются сигналы N-ацетильной группы N-ацетиласпартата (NAA, 5=2,0 ppm), протонов группы -N+(CH3)3 холинсодержащих соединений (Cho, 5=3,2 ppm), фосфокреатина + креатина (Cr, 5=3,0 ppm), -СН2-групп глутамата и глутамина (Glx, 5=2,2 и 3,7 ppm), сигнал 1,3, 4,6 - протонов шестичленного кольца инозитола (ml, 5=3,56 ppm) (Diehl et al., 1992).
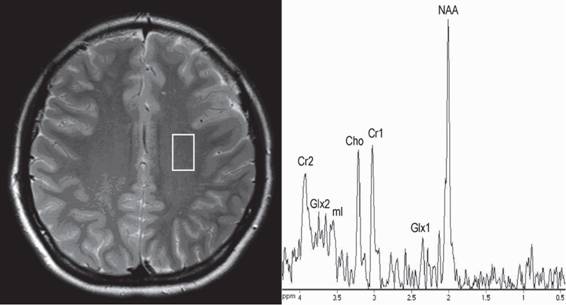
Рис. 1. Спектр белого вещества головного мозга (теменная доля), норма
NAA - N-ацетиласпартат, Glx - глутамат и глутамин, Cr - креатин+фосфокреатин, Cho - холинсодержащие соединения, ml - миоинозитол
В условиях гипоксии и ишемии в спектрах появляются сигналы метильной группы лактата (Lac, 5=1,34 ppm). Сигнал метильных и метиленовых протонов (Lip, 5=0,9 и 1,3), возникающий при некоторых патологических состояниях, обычно относят к жирнокислотным фрагментам липидов (Diehl et al., 1992).
Два сигнала - NAA и ml, используются как клеточные маркеры. NAA - нейрональный маркер, локализуется в нейронах; интенсивность этого сигнала пропорциональна уровню функционально полноценных нейронов в исследуемом объеме (Baslow, Guilfoyle, 2006). Сигнал ml - маркер астроцитов; рост интенсивности этого сигнала указывает на активацию глии или активацию пролиферации астроцитов (Ross et al., 1997). Остальные сигналы относятся к соединениям, распространенным во всех типах клеток нервной ткани. Их используют как метаболические маркеры (Ross, Bluml, 2001). Сигналы Cr и Cho является показателями энергетического и липидного обмена, соответственно. Интенсивность сигнала GLx характеризует уровень возбуждающего нейромедиатора глутамата.
Метаболизм NAA хорошо изучен. Это соединение синтезируется в митохондриях нейронов из аспартата и ацетил-коэнзима А под действием L-аспартат-N- ацетилтрансферазы и в незначительных количествах образуется как продукт катаболизма нейротрансмиттера N-ацетиласпартилглутамата. Разлагается NAA путем гидролиза в олигодендроцитах (рис.
2) (Baslow, Guilfoyle, 2006).Известно лишь одно заболевание - болезнь Канавана, при котором регистрируется превышающий норму уровень NAA (Baslow, Guilfoyle, 2006). Оно обусловлено отсутствием фермента гидролиза NAA, аспартоацилазы. При других заболеваниях уровень NAA либо снижается, либо не отличается от нормы. Это дает основания для заключения, что снижение NAA происходит вследствие нарушений процессов синтеза этого соединения и отражает снижение активности митохондрий нейронов.
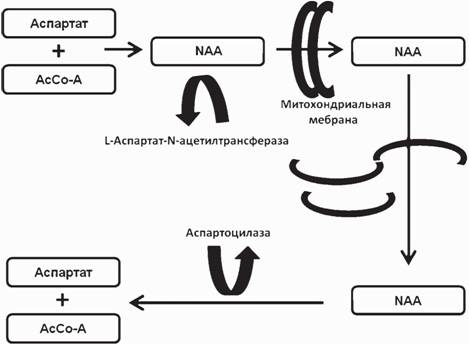
Рис. 2. Схема метаболизма NAA
Креатин не только транспортируется в мозг из печени, почек и поджелудочной железы, но и синтезируется в клетках мозга. Это соединение образуется в две стадии с участием аргинин-глицин-амадинотрансферазы и гуанидиноацетатметил- трансферазы. Показано, что в нейронах и глиальных клетках в большом количестве экспрессируются гены обоих ферментов (Brassiant et al., 2007). Считают (Brassiant et al., 2007), что основное количество церебрального креатина эндогенного происхождения. Фосфокреатин, продукт фосфорилирования креатина, является запасным источником макроэргов и используется для синтеза АТФ, при этом он превращается в креатин. Поэтому в условиях острой ишемии суммарное содержание креатина и фосфокреатина постоянно (Demougeot et al., 2004).
Сигнал Cho представляет собой суперпозицию резонансов холина, фосфохоли- на (PCho) и глицерофосфохолина. Основной вклад в сигнал вносит PCho, который в мозге, в частности - нейронах, синтезируется в трех последовательных реакциях переметилирования из фосфатидилэтаноламина (Magret et al., 1996). Холинсодержащие соединения являются предшественниками синтеза и продуктами распада мембранных фосфолипидов (Ross, Bluml, 2001).
4.
Еще по теме 1H МР-спектры мозга: отнесение сигналов, функция и метаболизм 1Н МР-видимых соединений:
- 5. Аминокислоты: метаболизм и функции
- 22 функции СП мозга
- Филогенетическое развитие ответной функции коры мозга.
- Психофизиологические функции основных структур головного и спинного мозга
- 25.функции сред мозга
- Нарушение функции мотонейронов спинного мозга.
- Нарушения функции спинного мозга.
- 24.Функции продолговатого мозга: и моста
- Онтогенез ответной функции коры мозга.
- Нарушения функций мозга за пределами нигростриатной системы
- Двигательные расстройства, связанные с нарушением функций коры большого мозга.
- Двигательные расстройства, связанные с нарушением функций спинного мозга.
- Основные функции коры большого мозга.
- Электрическая стимуляция спинного мозга - эффективный способ регуляции локомоторных функций
- 31 Соврем представл о Локализация функций в коре головного мозга (см. гл. 6, 7, 8)
- Дискуссия! а есть ли общее в выполняемых функциях оболочки ОС и оболочек мозга человека?!
- История вопроса о тормозящей функции двигательной зоны коры большого мозга.