Исследование процессов нейрогенеза в инъекционных моделях БА
Возникновение ряда мнестических нарушений может быть связано с изменением процессов нейрогенеза в мозге. Действительно, возрастное снижение нейрогенеза в субвентрикулярной зоне (СВЗ) и субгранулярной зоне (СГЗ) гиппокампа является одной из причин функциональных нарушений памяти (Kuhn et al., 1996; Kempermann et al., 1998, 2002).
Исследование нейрогенеза в биопсийном материале, полученном от больных БА, с использованием маркерных белков дифференциров- ки показало увеличение экспрессии белков незрелых клеток, таких как даблкортин (Dcx), полисиалилированный белок клеточной адгезии PSA-NCAM, нейрогенный фактор дифференцировки и TUC-4 (Jin et al., 2004b). В то же время, у пациентов с пресенильной стадией БА не было обнаружено усиления пролиферации нейральных предшественников, хотя число глиальных элементов, образованных de novo, возрастало (Boekhoom et al., 2006). Было также установлено, что при БА происходит нарушение механизмов созревания новообразованных нейронов в зубчатой фасции (ЗФ) (Li et al., 2008). В других работах было многократно показано, что при БА пролиферация предшественников и их число существенно снижаются (см. обзоры Hamilton et al., 2013; Lazarov et al., 2010).Использование нетрансгенных моделей БА также не позволяет сделать определенного заключения относительно влияния разных фрагментов белка-предшественника амилоида на нейрогенез. Так, введение в боковые желудочки мозга мышей AP(1-42) или AP(25-35) снижало уровень пролиферации клеток в СВЗ в течение последующих 5 дней (Haughey et al., 2002a). Похожие результаты были получены Х. Ли и П. Зу (Li, Zuo, 2005), которые показали, что интрацеребровентрикулярная инъекция Ар(25-35) сопровождается не только снижением нейрогенеза, но и нейродегенеративными изменениями в гиппокампе. Вместе с тем, увеличение пролиферации клеток в СВЗ также было продемонстрировано у мышей линии B6/DBA после введения AP(1-42) (Sotthibundhu et al., 2009).
Стимулирующий эффект AP(1- 42) на стволовые клетки был опосредован активацией рецептора p75NTR. Как видно из приведенных данных, результаты этих немногих работ так же противоречивы, как и полученные на мутантных животных. Практически отсутствуют данные для каких-либо других видов грызунов, кроме мышей.Следует отметить, что в значительной степени противоречивость получаемых данных может быть обусловлена различиями в используемых авторами методических подходах к детекции процессов нейрогенеза даже на сходных экспериментальных моделях. В первую очередь это связано с различными протоколами введения тимидинового аналога 5’-бромо-2’-дезоксиуридина (BrdU), который чаще всего используется для детекции пролиферации. У людей применение этого подхода невозможно, что затрудняет и без того непростую интерпретацию получаемых результатов. Так, единственное исследование (Jin et al., 2004b), выполненное на материале, полученном от людей, и демонстрирующее увеличение содержания Dcx в гиппокампе при БА, основано только на данных вестерн-блот-анализа, который не позволяет сделать выводов о местоположении и распределении клеток, экспрессирующих этот белок. В то же время, гистологическое исследование в когорте более молодых пациентов с БА позволило сделать вывод о том, что изменения пролиферации в гиппокампе связаны, главным образом, с изменением числа глиальных и эндотелиальных клеток (Boekhoorn et al., 2006).
В нашей работе исследование пролиферации было проведено с использованием многократного введения BrdU в течение 5 дней после инъекции AP(25-35) в боковые желудочки мозга крыс (Степаничев и др., 2009). Было установлено, что AP(25- 35) оказывал влияние на процессы пролиферации клеток и в СВЗ, и в СГЗ, но характер этих влияний был различным. В СВЗ число клеток, которые образовались в группе животных, инъецированных этим пептидом, не отличалось от контрольных крыс или крыс, получавших AP(35-25), но оставалось более высоким через 12 дней после операции, что может свидетельствовать об усилении пролиферации и/или нарушении миграционных процессов, связанных с действием AP(25-35).
В СГЗ пролиферация клеток была снижена по сравнению с другими группами животных и через 6, и через 12 дней после операции.Негативное влияние AP(1-42) и AP(25-35) при интрацеребровентрикулярном введении мышам было замечено в работе (Haughey et al., 2002a) также при субхроническом введении BrdU. Авторы описали этот эффект на качественном уровне при унилатеральном введении 5 нмоль AP(1-42) и AP(25-35). Они показали также, что аппликация этих пептидов в культуру нейросфер, полученных из эмбриональной ткани, приводила к нарушению пролиферации и нейрональной дифференцировки нейросфер (Haughey et al., 2002a,b), связанному с дисбалансом Ca2+. Следует отметить, что в этой работе авторы использовали существенно меньшую дозу пептидов и более поздние сроки по сравнению с нашим исследованием. В то же время, был обнаружен стимулирующий пролиферацию нейросфер эффект AP(1-42) в культуре, полученной из СВЗ (Lopez-Toledano, Shelanski, 2004; Sotthibundhu et al., 2009). Предполагают, что эти эффекты обусловлены активацией рецептора p75NTR.
Можно предположить, что стимулирующий эффект Ap связан с длительным воздействием на относительно многочисленную популяцию делящихся клеток, которая включает в себя как непосредственно стволовые клетки, так и незрелые пролиферирующие нейробласты. Для того чтобы исследовать эффекты AP(25-35) на более однородную по составу группу клеток, была использована иная схема введения BrdU, при которой этот предшественник вводили крысам только в течение первых суток после инъекции AP(25-35). Иммуногистохимическая детекция включившегося BrdU была проведена через 6, 12 и 28 дней после инъекций AP(25-35) или растворителя. Было установлено, что общее число ВМи+-клеток в СВЗ у контрольных крыс при такой схеме введения было меньше, чем в предыдущей серии экспериментов, и на протяжении всего времени наблюдения не подвергалось существенному изменению. В то же время, через 6 дней после введения AP(25-35) число новых клеток было значительно увеличено. Число ВМи+-клеток не отличалось достоверно от контрольного уровня уже через 12 дней после операции, но все еще оставалось весьма высоким.
Это также свидетельствует в пользу предположения о том, что AP(25-35) не только усиливает пролиферацию в СВЗ, но и снижает скорость миграции новых клеток по ростральному миграционному пути.Иная картина изменений наблюдается при исследовании пролиферации и созревания клеток в СГЗ. Так, при субхроническом введении BrdU наблюдалось существенное снижение числа ВМи+-клеток в СГЗ ЗФ мозга крыс как через 6, так и через 12 дней после инъекции AP(25-35) по сравнению с контролем или пептидом AP(35-25). Тем не менее, при исследовании популяции «новых» клеток, образовавшихся через 1 день после операции, нам не удалось выявить каких-либо различий в числе ВМи+-клеток ни на одном из исследованных сроков. В то же время, AP(25-35) существенно изменял характер созревания клеток. Так, через 28 дней после инъекции AP(25-35) доля ВМи+-клеток, экспрессировавших маркер незрелых нейронов Dcx, была значительно выше в этой группе крыс по сравнению с контролем. Кроме того, доля BrdU+-клеток, экспрессировавших маркер зрелых нейронов NeuN, была достоверно ниже по сравнению с контролем.
Для того чтобы понять, какие механизмы могут быть задействованы в опосредовании эффектов AP(25-35), необходимо кратко упомянуть об основных процессах, контролирующих нейрогенез. Наши представления об этих процессах в основном базируются на исследованиях механизмов деления клеток и экспрессии маркеров нейробластов. Стволовые клетки и предшественники делятся в специфическом микроокружении нейрогенной ниши. Нейроны ЗФ получают возбуждающую глутаматергическую иннервацию из энторинальной коры, а ГАМК-ерги- ческая тормозная иннервация обеспечивается интернейронами самой ЗФ. Помимо этого нейроны ЗФ получают входы из разных отделов мозга, передача сигнала в которых опосредована различными медиаторными системами (Андреева и др., 1985; Гасанов, Меликов, 1986; Отмахов, 1993; Lie et al., 2004). Наиболее исследована роль глутамата и ГАМК в контроле деления и дифференцировки клеток в СГЗ ЗФ.
Считается, что глутаматная трансмиссия, опосредованная NMDA-рецепторами, обратно коррелирует с уровнем пролиферации в гиппокампе (Lie et al., 2004). В то же время, сами нейробласты не экспрессируют NRl-субъединицу NMDA-рецепторов, играющую ведущую роль в нейротрансмиссии (Deisseroth et al., 2004), однако эти клетки деполяризуются в ответ на аппликацию ГАМК, а, кроме того, ГАМК-ергическая стимуляция способствует дифференцировке гиппокампальных предшественников типа 2 (Tozuka et al., 2005). По-видимому, ГАМК играет более выраженную роль в процессах пролиферации и дифференцировки новых клеток в СГЗ гиппокампа как в обычных условиях, так и в условиях патологии (Аниол, Сте- паничев, 2007; Deisseroth, Malenka, 2005). Кроме того, пептидные нейромедиаторы также могут оказывать влияние на нейробласты СВЗ и СГЗ (Lie et al., 2004).Нейральные стволовые клетки, подобно стволовым клеткам из других органов и тканей, экспрессируют рецепторы к большому набору ростовых факторов, что свидетельствует о возможной регуляции пролиферации этих клеток такими молекулами. На сегодняшний день получены свидетельства того, что практически все известные ростовые факторы в той или иной мере обладают способностью воздействовать на нейрогенез. Например, повышение уровня BDNF в нейрогенных областях приводит к усилению пролиферации предшественников (Benraiss et al., 2001). Установлено, что пролиферация клеток, взятых из субвентрикулярной области in vitro, требует наличия нескольких ростовых факторов (Reynolds, Weiss, 1992). К их числу, наряду с уже упомянутым BDNF, относятся эпидермальный фактор роста (EGF) и основный фактор роста фибробластов (bFGF). Предполагается, что эти же ростовые факторы стимулируют нейрогенез in vivo (Kuhn et al., 1997; Doetsch et al., 2002).
Можно предположить, что отрицательное влияние Ар на пролиферацию клеток связано с нарушением трофического обеспечения. Так, установлено, что агрегированный Ар негативно влияет на уровень BDNF у трансгенных мышей (Peng et al., 2009).
Введение АР(1-42) в мозг крыс также приводит к снижению содержания этого нейротрофина (Christensen et al., 2008). С другой стороны, длительное введение АР(1-42), АР(1-40) или АР(25-35) приводило к увеличению экспрессии BDNF (Tang et al., 2000).Клетки пролиферативных областей мозга могут находиться под влиянием и других внешних факторов, среди которых хотелось бы особо остановиться на воздействии свободных радикалов. ГАМК-ергические нейроны Аммонова рога и ЗФ могут служить источником NO. При этом оказывается, что области, богатые нит- рергическими нейронами, анатомически тесно связаны с областями интенсивного нейрогенеза. Такое наблюдение справедливо как для гиппокампа, где нитрергичес- кие нейроны гранулярного слоя и хилуса находятся в непосредственной близости от пролиферирующих клеток СГЗ ЗФ (Estrada, Murillo-Carretero, 2005), так и для СВЗ (Moreno-Lopez et al., 2000). Последняя богата аксонами нитрергических нейронов, образующими здесь густую сеть, в петлях которой расположены кластеры пролиферирующих нейробластов. Отростки нейронов, экспрессирующих nNOS, наблюдаются также и на всей протяженности рострального миграционного русла, где они тесно переплетаются с цепочками пролиферирующих и мигрирующих клеток. В самих нейробластах, мигрирующих от субвентрикулярной области, экспрессии nNOS не отмечается, и лишь в некоторых клетках, уже достигших гранулярного и перигломерулярного слоев обонятельной луковицы, начинает экспрессироваться nNOS. Указанные области обонятельной луковицы являются местами, где происходит дифференцировка нейральных предшественников, что, по мнению Е.Р. Матарредона и др. (Matarredona et al., 2004), может говорить как о дифференцировке нейральных предшественников в нитрергические нейроны, так и о временной экспрессии nNOS в процессе дифференцировки нейронов. Все это позволяет задуматься о возможном участии нитрергических механизмов в регуляции процессов нейрогенеза не только на ранних этапах онтогенеза (Santacana et al., 1998; Peunova et al., 2001), но и в мозге взрослых животных (Moreno-Lopez et al., 2000). Также известно, что NO подавляет пролиферацию многих типов клеток in vitro (Murillo-Carretero et al., 2002), при этом блокируя пролиферативные эффекты некоторых ростовых факторов, в частности EGF. NO подавляет пролиферацию предшественников нейронов в СВЗ (Moreno-Lopez et al., 2000). При этом авторам не удалось наблюдать аналогичного эффекта в ЗФ. А. Ченг и др. (Cheng et al., 2003) установили, что система NO участвует в механизме действия BDNF на дифферен- цировку нейральных предшественников. В частности, было показано, что BDNF повышает экспрессию nNOS в предшественниках нейронов в СВЗ, останавливая их пролиферацию и запуская дифференцировку, а ингибирование nNOS блокирует дифференцирующее действие BDNF.
В настоящее время остается неясным, как на молекулярном уровне NO оказывает свое антипролиферативное и дифференцирующее действие. С.Р. Джеффри с соавторами (Jaffrey et al., 2001) полагают, что в качестве такого механизма выступает осуществляемое NO нитрозилирование -SH групп цистеиновых остатков различных рецепторных и регуляторных белков с образованием нитрозотиолов, что приводит к изменению активности белков-мишеней. В подтверждение этой концепции авторы работы (Murillo-Carretero et al., 2002) показали, что NO может обра-
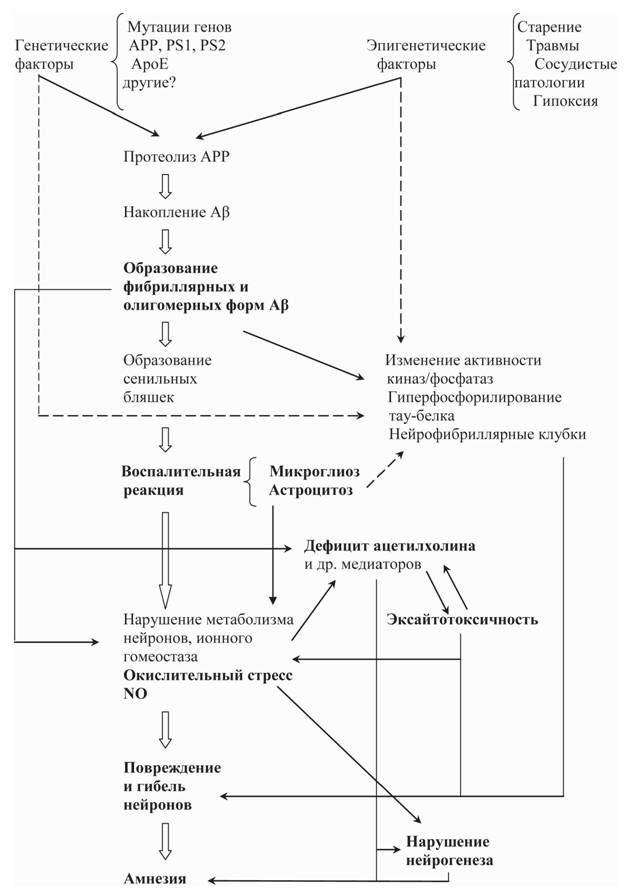
Рис. 1. Гипотетическая схема событий, ведущих к возникновению амнезии альцгеймеровского
типа
Жирным шрифтом выделены процессы, развитие которых можно наблюдать в так называемой «амилоидной» модели БА
тимо нитрозилировать рецептор EGF. Рецептор EGF представляет собой мембранную тирозинкиназу, димеризующуюся при связывании EGF и взаимно фосфори- лирующуюся по остаткам тирозина. Воздействие NO приводит к ингибированию реакции фосфорилирования рецептора EGF, а также к замедлению пролиферации, вызванной EGF.
Можно предположить, что помимо NO и другие свободные радикалы и метаболиты окислительного стресса могут оказывать воздействие на обсуждаемые процессы. Например, аппликация АР(1-40) в культуру стволовых клеток человека приводила к увеличению в них содержания карбонильных групп в белках и продуктов ПОЛ (Mazur-Kolecka et al., 2006). Окислительный стресс, вызванный радиоактивным облучением (Fishman et al., 2009) или экспозицией в атмосфере, обогащенной озоном (Rivas-Arancibia et al., 2010), ведут к угнетению нейрогенеза. В пользу этого свидетельствует также подавление нейрогенеза в результате эмоционального стресса (Heine et al., 2005; Dagyte et al., 2009). Известно, что и острый, и хронический эмоциональный стресс сопровождаются изменениями свободнорадикального окисления (Айрапетянц и др. 2006) и в ряде случаев повреждением нейронов гиппокампа (Тишкина и др., 2009). Возможно, что дополнительный негативный эффект стресса на нейрогенез связан с выбросом стрессорных гормонов в кровь (Fuchs et al., 2001). Косвенным свидетельством того, что свободные радикалы до определенной степени могут контролировать и/или изменять процессы пролиферации клеток в мозге, является ее усиление вследствие двигательной активности (Van Praag et al., 1999a,b). Было показано, что умеренная физическая нагрузка повышает антиоксидантную защиту организма в целом (Gomez-Cabrera et al., 2008) и мозга в частности (Radak et al., 2007), и снижает вероятность возникновения БА (Radak et al., 2010). Таким образом, окислительный стресс, вызываемый в гиппокампе введением AP(25-35) в боковые желудочки, может быть одной из причин угнетения пролиферации и замедления дифференцировки в СГЗ ЗФ.
6.