Исследование механизмов инициации локомоторной активности при эпидуральной стимуляции спинного мозга
Как было отмечено выше, параметры восстановительной ЭССМ подбирают эмпирически. На выбор параметров ЭССМ, необходимых пациенту с конкретной патологией, требуется время, в то время как многие авторы отмечают, что эффективность ЭССМ тем выше, чем быстрее после опреации она была начата (Тома и др., 2010; Макаровский и др., 2012а).
Таким образом, исследования, проведенные на животных с повреждениями спинного мозга, моделирующими патологию, должны предшествовать применению ЭССМ в лечебных мероприятиях, выбор параметров ЭССМ должен быть объективно обоснован.ЭССМ, использующаяся в клинических целях, получила неожиданное развитие в контексте изучения локомоции, после того как было показано, что приложение эпидуральной стимуляции к задней поверхности спинного мозга на уровне сегмента L2 может вызывать у спинальных больных ритмические движения ног (Dimitrijevic et al., 1998; Shapkova, Schomburg, 2000; Герасименко, 2002). Этот феномен, породивший много споров об адекватности данного метода для активации ГШД у человека, ставит необходимость проведения экспериментов на интактном спинном мозге и выяснения механизмов влияния ЭССМ на двигательные функции спинного мозга. Такая возможность была реализована на модели мезэнцефалической кошки. Основное преимущество данной модели состоит в сохранности мезенцефалической локомоторной области ствола головного мозга и отсутствии спонтанной локомоторной активности, обусловленной гипоталамической локомоторной областью, мешающей изучению стимуляционных воздействий (Шик и др., 1966).
На модели децеребрированной кошки проводилось картирование дорсальной поверхности спинного мозга для определения наиболее эффективного сегмента для вызова локомоторной активности (Лавров, 2002; Герасименко и др., 2003; Мусиенко, 2005). Последовательно стимулировали L3-L7 сегменты спинного мозга. Использовали импульсы тока длительностью 0,5 мс, амплитудой от 0-100 мкА и частотой следования импульсов 0,5-100 Гц.
Картирование спинного мозга проводилось в ростро-каудальном и медиолатеральном направлениях. При стимуляции L2 сегмента у всех животных вызывалась только тоническая активность в мышцах задних конечностей. Стимуляция сегмента L3 могла вызывать локомоторную активность, однако она возникала после довольно длительной фазы тонического сокращения мышц (15-20 с). Наиболее легко локомоция вызывалась при расположении электрода на уровне L4-5 сегмента.При стимуляции этих сегментов локомоторная активность появлялась практически без фазы тонической активации мышц, а порог вызова локомоции был наиболее низким. Стимуляция более каудальных сегментов L6, L7 не приводила к появлению ритмической активности, вызывая только тоническую активацию мышц задних конечностей. Сравнительный анализ зоны L3 и L4 для запуска локомоторной активности выявил следующие особенности. Оказалось, что если время формирования ритмической активности в проксимальных мышцах (m. quadriceps и m. semitendinosus) при стимуляции этих локусов практически не отличалось, то в дистальных мышцах (m. tibialis ant. и m. gastrocnemius) латентный период появления ритмики укорачивался приблизительно
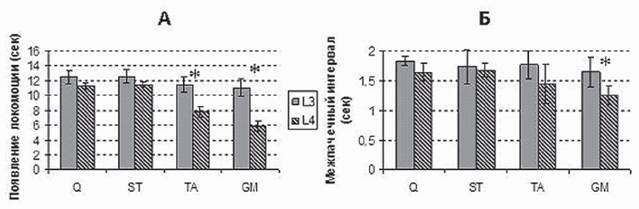
Рис. 4. Гистограммы зависимости времени появления ритмической активности в мышцах от уровня стимуляции спинного мозга (А), величины средних интервалов между соседними ЭМГ пачками для мышц нижних конечностей (Б), показаны доверительные интервалы (p=0,05). Q - m. quadriceps, ST - m. semitendinosus, TA - m. tibialis anterior, GM - m. gastrocnemius
в два раза при стимуляции L4 в сравнении с L3 (рис. 4). При этом частота следования пачек ЭМГ активности в данных мышцах была выше по сравнению с ритмической активностью, опосредуемой стимуляцией вышележащего сегмента L3 (рис. 4Б).
Дополнительным аргументом в пользу большей эффективности области сегмента L4 в инициации локомоторной активности служат данные об эффектах последействия стимуляции.
Если после прекращения эпидуральной стимуляции на уровне L3, локомоторная активность продолжала продуцироваться еще в течение 3-4 с, то после прекращения стимуляции на уровне L4 сегмента, длительность ее последействия была в 2-3 раза больше (рис. 5). Феномен продолжения локомоторной активности после выключения стимуляции свидетельствует о циклической активации внутриспинальной нейронной сети, входящей в состав ГТТТД.Дополнительные аргументы в пользу ведущей роли сегментов L4-L5 в формировании шагательного паттерна были получены при проведении дополнительной математической обработки (Богачева и др., 2005; Богачева, 2006). При помощи преобразования Гильберта были построены огибающие сигналов ЭМГ, поскольку именно огибающие отражают мышечные усилия (Андреева, Хуторская, 1987). При сравнении кросскорреляционных функций (ККФ) ЭМГ с сигналами стимула было установлено, что в случаях стимуляции сегментов L2, L3, L6 наблюдаются значимые пики на ККФ, частота появления которых точно соответствует частоте стимуляции, а в случае L4, когда возможно появление локомоции, пики на ККФ на частоте стимуляции становятся незначимыми и возникает гармоника 0,9-1,0 Гц, соответствующая частоте шагания. Кросскорреляционный анализ связи сигналов ЭМГ с сигналами стимула показывает, что эта связь, значимая для случаев со стимуляцией сегментов L2 и L6, ослабевает в случае L4 (рис. 6). Это означает, что возникающая локомоторная активность при стимуляции L4-L5 сегментов осуществляется не по типу стимул-ответ, а за счет генерации собственного внутриспинального ритма, опосредованного активацией генератора шагательных движений.
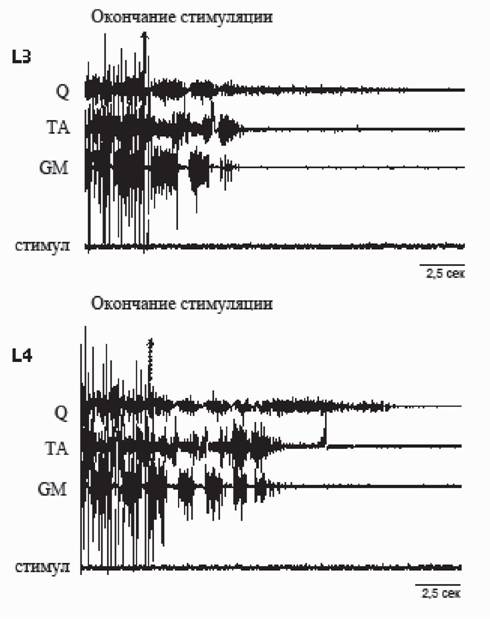
Рис. 5. «Эффект последействия» при эпидуральной стимуляции на уровне сегментов L3 и L4. Обозначения те же, что на рис. 4
Итак, наиболее эффективной зоной для вызова локомоторной активности является область, расположенная на уровне L4 сегмента. Далее было исследовано, на какие параметры стимулирующих воздействий оптимально отвечает система.
Эффект частоты стимуляции исследовали в серии экспериментов с использованием фиксированной интенсивности стимулирующего воздействия, равной двум порогам, для рефлекторных ответов мышц задних конечностей на стимулы, прикладываемые к дорсальной поверхности спинного мозга на уровне L4. Было установлено, что формирование локомоторной активности происходит при частоте стимуляции 5-10 Гц. При более низкой частоте (1-2 Гц) локомоторная активность не вызывается, как правило, на каждый стимул формируются отдельные ответы. Вместе с тем было отмечено, что даже на одиночную стимуляцию рефлекторные ответы в мышцах-антагонистах возникают
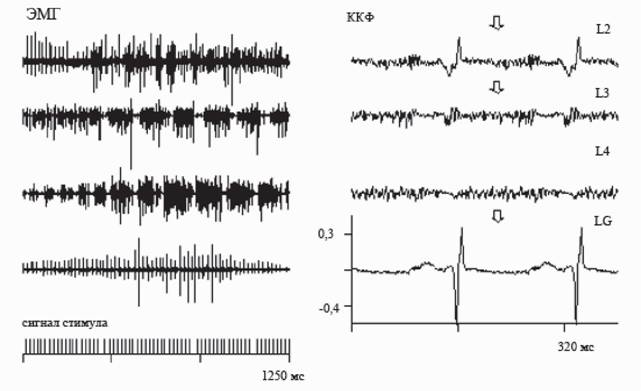
Рис. 6. Слева паттерны ЭМГ-активности, зарегистрированные при эпидуральной стимуляции сегментов L2, L3, L4, L6. Справа кросскорреляционные функции (ККФ) паттернов активности и сигналов стимула.
Стрелки - значимые пики, соответствующие частоте стимуляции
реципрокно. Это говорит о том, что при низкочастотной стимуляции можно подобрать параметры стимуляции, вызывающие локомоторную активность. При использовании частоты стимуляции свыше 20 Гц локомоторная активность быстро угасает и для ее поддержания требуется повышение интенсивности стимула (рис. 7). Тот факт, что эф-
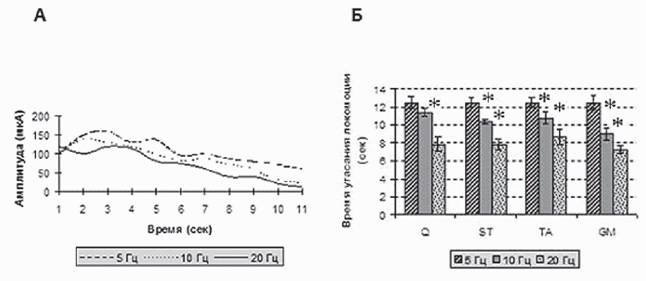
Рис. 7. А - изменение амплитуды пачечной активности для m. gastrocnemius при различных частотах эпидуральной стимуляции сегмента L4. Б - длительность угасания ритмической активности при разных частотах эпидуральной стимуляции
фект последействия выражен значительно сильнее при стимуляции 5 Гц, также доказывает, что данная частота стимуляции является более эффективной для активации ГШД.
Рассмотрение автокорреляционных функций ЭМГ, зарегистрированных при различных частотах стимуляции (1-20 Гц), выявляет в случае стимуляции 5 Гц четко выраженную частоту, отвечающую частоте шагания, а связь со стимулом в этом случае становится незначимой (рис.
8). Это свидетельствует об эффективности данной частоты в запуске ГШД, продуцирующего шагательный паттерн в присущем ему ритме.Ю.П. Герасименко с соавторами (Gerasimenko et al., 2008), анализируя случаи применения ЭССМ в клинике, делают вывод о том, что за последнее десятилетие был достигнут значительный прогресс в использовании ЭССМ для восстановления функции поддержки веса тела и передвижения у пациентов с тяжелыми повреждениями спинного мозга, не способных самостоятельно стоять и ходить. Этот прогресс обусловлен, главным образом, достижениями, полученными в экспериментах с лабораторными животными, имеющими полное пересечение спинного мозга на среднем грудном уровне и у людей с полным перерывом спинного мозга. Рис. 9, взятый из упомянутого обзора, иллюстрирует сходство локомоторных нейронных сетей (ГШД) у человека и животных.
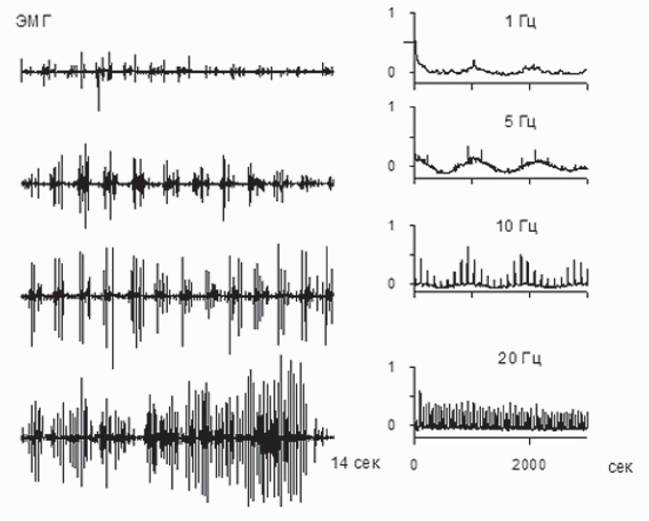
Рис. 8. Паттерны ЭМГ-активности при различных частотах эпидуральной стимуляции сегмента L4 (слева) и их автокорреляционные функции (справа)
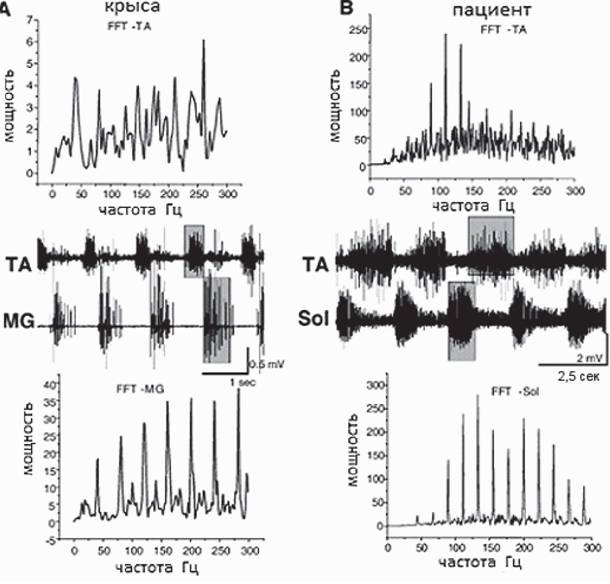
Рис. 9. Спектральные характеристики мышц сгибателя (TA) и разгибателя (MG) у спинальной крысы (A) и мышц сгибателя (TA) и разгибателя (Sol) ноги пациента с полным перерывом спинного мозга (B) во время шагоподобной активности, вызванной ЭССМ (S1,40 Гц для крысы и L2, 25 Гц для человека)
Спектральный анализ получен быстрым Фурье-преобразованием. Цит. по (Gerasimenko et al., 2008)
С тех пор появились новые исследования, позволившие определить структуры спинного мозга, стимуляция которых инициирует поддержку веса тела. В условиях гравитационного поля земли ходьба, как и другие произвольные движения, не могут осуществляться только за счет ритмичных движений конечностей. Поддержка веса тела, позы является обязательным компонентом активных движений. Позная компонента локомоции также утрачивается в отсутствие супраспинальных влияний.
Одним из достижений последних лет является то, что в спинном мозге были определены места, электрическая стимуляция которых приводит к появлению функции поддержки веса тела, эти локусы находятся в сакральном отделе спинного мозга (рис. 10).
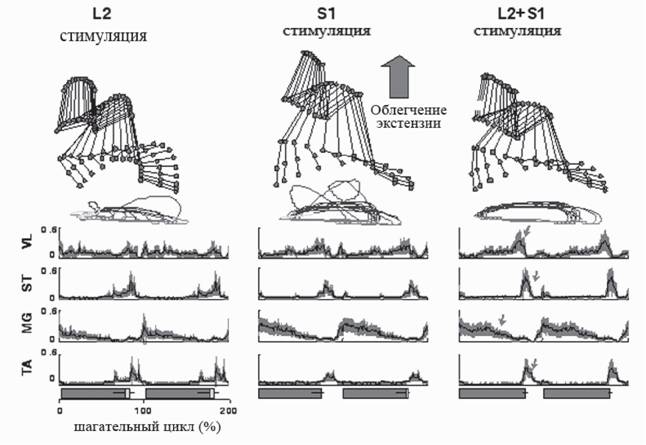
Рис. 10. Эффект одновременной эпидуральной стимуляции L2 и S1 сегментов спинного мозга крыс, спинализированных на уровне Th9
Вверху - реконструкция движений суставов задней конечности по видеозаписи, временные срезы. Средний ряд - реконструкция траектории движений стопы, демонстрирует положение стопы во время фазы опоры и фазы переноса. Нижний ряд - суммарные интегрированные электромиографические записи четырех мышц задней конечности: VL - vastus lateralis; ST - semitentinosus; MG - medial gastrocnemius; TA - tibialis anterior
Стрелками показаны пачки ЭМГ-активности, возникающие при одновременной стимуляции L2+S1. Внизу - продолжительность шагательного цикла с фазой опоры (серый) и фазой переноса (белый). Цит. по (Courtine et al., 2009)
В экспериментах на взрослых крысах, хронически спинализированных на нижнегрудном уровне (Th9), исследовались эффекты одновременной электрической эпидуральной стимуляции нескольких сегментов спинного мозга для инициации локомоторного поведения. Селективная электрическая стимуляция с частотой 40 Гц поясничного L2 сегмента, в котором локализован генератор шагательных движений, вызывала ша- гательную ритмику в задних конечностях. Как было сказано выше, сегмент спинного мозга L2, хоть и не является оптимальным для вызова локомоции на модели децеребри- рованной кошки (интактный спинной мозг), но при его стимуляции также может быть вызвана ходьба. То же можно сказать и про высокие частоты стимуляции. 5 Гц - это та частота, при которой ритмические локомоторные движения вызываются наименьшими токами, однако и высокие частоты могут вызывать локомоцию при больших амплитудах стимула. На экспериментальной модели крысы с хронической перерезкой спинного мозга частота 40 Гц и сегмент L2 оказались оптимальными для вызова локомоции. Однако вызванные шагоподобные движения осуществлялись с волочением стопы и без поддержки веса тела (отсутствовала фаза опоры). Электрическая стимуляция с частотой 40 Гц области локализации моторных пулов мышц-разгибателей (крестцовый S1 сегмент) вызывала ритмическую активность в задних конечностях с преимущественной активацией экстензорных мышц (разгибателей) и появление выраженной фазы опоры. Одновременная стимуляция L2 и S1 сегментов значительно улучшала качество шагательных движений. Крысы демонстрировали хорошо координированную локомоторную активность с правильной постановкой стопы (см. рис. 9) (Courtine et al., 2009).
Позднее на крысах с полной перерезкой спинного мозга было показано, что уже через неделю после перерезки мозга при стимуляции L2 и S1 сегментов и одновременном фармакологическом воздействии (активация серотониновых рецепторов) животные способны были шагать по движущейся ленте тренажера, самостоятельно поддерживая вес тела; при этом кинематические характеристики суставов конечностей практически не отличаются от этих характеристик у нормальных животных (Герасименко и др., 2011а,б).
Таким образом, можно считать доказанным, что электрическая стимуляция поясничного отдела спинного мозга является неспецифическим активатором локомоторных генераторов. Исследования, проведенные на экспериментальных животных, показали, что структурами спинного мозга, оптимальными для вызова локомоции, являются сегменты L2-L4, для поддержания веса тела - сегменты S1-S2. Частота ЭССМ 5 Гц является эффективной для вызова движений на модели децеребрированной кошки (интактный спинной мозг в отсутствие супраспинальных влияний). Частота ЭССМ 40 Гц эффективна для вызова локомоции и поддержки веса тела на модели спинальной крысы (полный перерыв спинного мозга в нижнегрудной области).
5.
Еще по теме Исследование механизмов инициации локомоторной активности при эпидуральной стимуляции спинного мозга:
- Электрическая стимуляция спинного мозга - эффективный способ регуляции локомоторных функций
- Применение метода эпидуральной электростимуляции спинного мозга в терапии спинного мозга
- Биофизические основы электрической стимуляции спинного мозга
- Неинвазивная стимуляция спинного мозга для вызова шагательных движений
- Тактика при компрессии спинного мозга
- Ремоделирование нервной ткани при повреждениях спинного мозга
- Результаты исследования биоритмов спинного мозга в группе Ог
- Результаты исследования биоритмов спинного мозга в группе К
- Расстройства движений при повреждении спинного мозга
- Основные типы операций при различных опухолях спинного мозга.
- 3. 4. 3. Анализ МРТ – иследования головного и спинного мозга при рассеянном склерозе
- Результаты исследования биоритмов спинного мозга в группе От
- Обсуждение результатов исследования биоритмической активности спинного мозга в норме