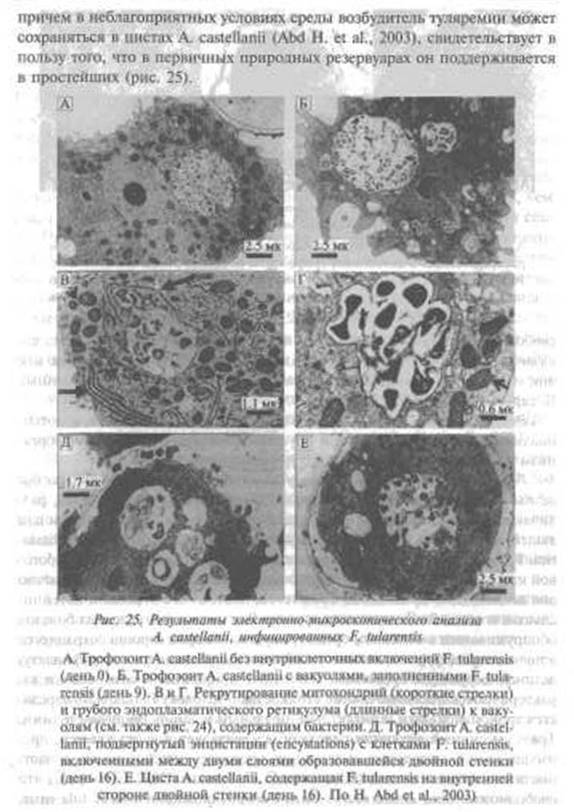
Границы феномена сапронозного существования патогенных для людей микроорганизмов.
О том, что границы этого феномена очень широки, свидетельствует много косвенных факторов. Например, все так называемые возбудители опасных и особоопасных инфекционных болезней (чумы, мелиоидоза, сапа, туляремии, сибирской язвы, бруцеллеза, лихорадки Ky и др.), одновременно являющиеся и потенциальными агентами биологического оружия, имеют много сходства в эпидемиологии, биологических свойствах и в клинике вызываемых ими поражений людей, объяснить которое возможно только в том случае, если предположить, что они являются истинными паразитами (или эндосимбионтами) почвенных простейших.
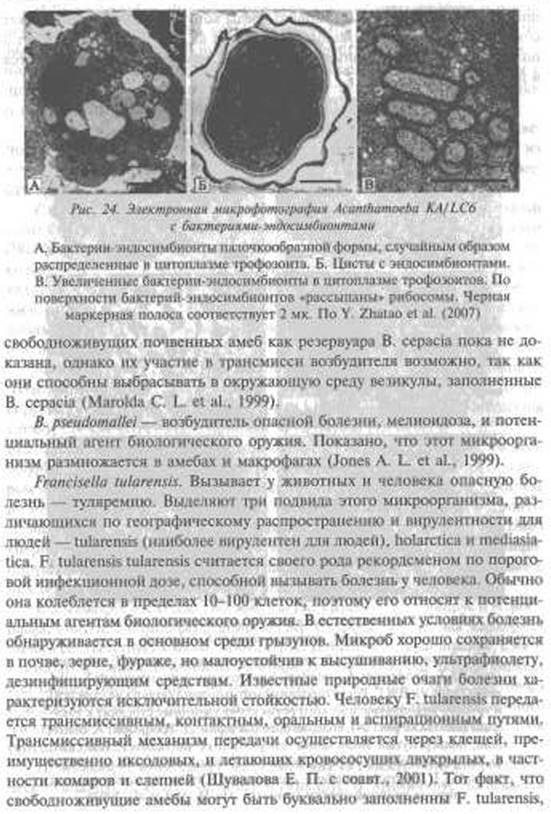
В пользу такого предположения говорит «привязанность» вызываемых ими вспышек болезней к конкретным местностям; отсутствие у них резистентности детоксикационного типа к антибиотикам (ненужной для внутриклеточного паразитизма); вовлечение в инфекционный процесс лимфатических узлов и фагоцитирующих клеток крови; а также «запутанность» вопроса о факторах их патогенности и токсинах. Ни один из обнаруженных у них таких «факторов» не может однозначно в эксперименте прояснить клинику вызываемой микроорганизмом у людей болезни, больше похожую на реакцию организма на суперантигенный раздражитель. Не обладают они и конгагиозностъю, т, е. способностью передаваться от одного теплокровного организма к другому при контакте, что обычно предполагает воздушно-капельный механизм передачи возбудителя болезни непосредственно от одного заболевшего к другому. Редкие случаи «перехода» не контагиозной бубонной чумы в контагиозную вторично-легочную лишь исключения, подтверждающие вышеприведенные наблюдения. Ни больные с такой формой чумы, ни появившиеся через инфицирование от них больные с первично-легочной чумой не способны поддерживать возбудитель чумы в природе, так как в 100 % известных случаев (без лечения антибиотиками) они погибают. И если этим возбудителям инфекционных болезней не придумывать эпидемиологию (см. «Исторические свидетельства»), то понять, как они существуют в природе без резервуара среди простейших, практически невозможно.Vibrio cholerae. Возбудитель холеры является членом семейства Vibrio- пасеае, о котором известно, что оно прекрасно размножается в простейших A. polyphaga и N. gruberi. Их выживание в пределах цист N. gruberi предполагает то, что свободноживущие амебы могут длительно сохранять холерный вибрион при неблагоприятных условиях среды (Thom S. et аі.,1992). Достоверно установлены биоценотические связи холерного вибриона, обитающего в воде, с водными организмами, в том числе и с растениями. Например, была показана способность ряски, зоопланктона (Islam М. et аі., 1990) и низших ракообразных (Голубев Б. П., 1993) поддерживать высокую концентрацию холерного вибриона. М. Islam et аі. (1990) также выявили активное размножение и длительную (до 15 месяцев!) персистенцию некультивируемых вибрионов Эль Тор в культуре сине-зеленых водорослей, которые признаются ими возможным резервуаром холеры в межэпидемические периоды. А. С, Марамович и М. И. Наркевич (1993) пришли к выводу о том, что водный гиацинт является природным резервуаром холеры Эль Тор. Он обеспечивает существование вибрионов в межэпидемические периоды и способствует реализации водного пути передачи возбудителя. О. В. Бухарин и В. Ю. Литвин (1997) экспериментально показали, что водный гиацинт способствует размножению холерных вибрионов. Их концентрация в его стеблях и листьях была в 300 раз выше, чем в воде.
Эти и другие данные позволили В. И. Пушкаревой и В. Ю, Литвину (Пушкарева В. И., Литвин В. Ю., 1994; Пушкарева В. И., 1994), выдвинуть гипотезу о клонально-селекционном механизме изменения бактерий в почвенных и водных сообществах, способном обеспечивать формирование эпидемически значимых вариантов возбудителей сапронозов в их природных резервуарах. Гипотеза весьма интересная, поэтому ее стоит привести ниже,
В процессе естественной циркуляции холерных вибрионов среди гид- робионтов — хозяев возбудителя в водоемах, имеет место селекция TOK- сигенных клонов и их накопление в бактериальной популяции при благоприятных условиях (активном пассировании через хозяев).
Всего же у холерных вибрионов в водной экосистеме формируется, как минимум, две экологические ниши. Одна из них (непосредственно водная среда) наиболее характерна для авирулентных штаммов. Вторую нишу (сообщество водных организмов) населяет преимущественно вирулентная часть микробной популяции, устойчивая к перевариванию в организме гидробион- тов — первичная функция токсигенности холерных вибрионов, возможно, как раз и состоит в защите бактериальной популяции, обитающей в водоемах, от хищничества простейших и других гидробионтов. Их объем относительно друг друга меняется. Например, при изменении численности хозяев или сдвигах в структуре водного сообщества. В разных условиях и в разные сезоны в водной популяции вибрионов могут доминировать то токсиген- ные, то атоксигенные клоны холерных вибрионов. Изменение уровня токсигенности всей популяции вибрионов во времени происходит за счет выхода в водную среду нарастающего числа токсигенных вибрионов из организма погибших инфузорий, где они накапливаются благодаря селективному преимуществу перед атоксигенными и слаботоксигенными вибрионами. Прогревание воды до температуры 20°С и резкое увеличение трофности водоемов в июле — августе определяют пик численности холерных вибрионов, тесно сцепленный с пиком численности планктона. Интенсивно пассируясь среди гвдробиошов, вибрионы окончательно выходят из покоящегося состояния (число бактериологически высеваемых культур максимально); численность и вирулентность водной популяции вибрионов резко возрастает, достигая эпидемически значимых показателей. Именно к этому периоду неизменно приурочен пик заболеваемости людей в очагах умеренных широт. Первичные и независимые случаи инфицирования холерой людей, связаны с водоемами, после чего распространение холеры может принять и вторичный (эстафетный) характер, в виде классической вспышки (по работе Бухарина О. В. и Литвина В. Ю., 1997).Приведенная выше клонально-селекционная теория состыковывается с гениальными прозрениями М. Пепенкофера (1885) следующим образом.
Если принять нетоксигенные клоны холеного вибриона за фактор X, то гидробионты, через которые пассируются токсигенные клоны возбудителя холеры, можно считать тем фактором Y, существование которого Пет- тенкофер так настойчиво отстаивал, а сами токсигенные клоны представляют собой фактор Z. Однако только ли для холерного вибриона верна эта теория Петтенкофера?Легионеллы. Для легионелл возможность сапронозного существования установлена бактериологическими методами еще в 1950-х гг. Вспышки легионеллезов обычно сопровождаются высокой смертностью среди заболевших людей. Поэтому экология легионелл и механизмы их проникновения в человеческие популяции находятся в поле зрения ученых уже не менее 30 лет, с момента печально известной вспышки болезни в Филадельфии в 1976 г. В данной работе легионеллы рассматриваются как -«опережающий объект» в исследованиях сапронозов, что предполагает перенесение выявленных при их изучении закономерностей на менее исследованные микроорганизмы, обитающие в почвенных и водных экосистемах.
В пределах водных сообществ легионеллы существуют в ассоциации с планктоном или как составная часть биопленок (Rogers J, et al., 1994). Амебы имеют критическое значение для существования легионелл в окружающей среде и для их способности вызывать инфекционную болезнь у людей (НаіЪ О. et аі., 2000). Схематическое изображение взаимодействия L. pneumophila и одноклеточных животных в окружающей среде и механизма ее трансмиссии к человеку приведено на рис. 22.
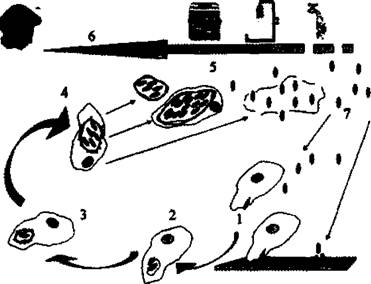
Рис. 22. Механизм взаимодействия L pneumophila и одноклеточных животных в окружающей среде и их трансмиссии к человеку
I — L. pneumophila из биопленок, формируемых совместно с другими бактериями или из их суспензий, инфицирует Protozoa. Проникновение в простейшие осуществляется по механизму фагоцитоза; 2 — после проникновения в амебу L. pneumophila остается в мембранно-связанной вакуоле, которая рекрутируется органеллами, такими как митохондрии, и не сливается с лизосомами;
3 — в пределах 4 ч после проникновения вакуоль с L.
pneumophila окружается эндоплазматическим ретикулумом; 4 — L. pneumophila реплицируется в пределах специализированной вакуоли; 5 — точный механизм «выхода» бактерий из простейших неизвестен, но считается, что возможна их экскреция в составе везикул. Могут лизироваться и сами амебы; б — передача L. pneumophila к людям осуществляется механически, через воздух, выбрасываемый кондиционерами, с каплями воды в душевых и т. п. способами; 7 — Legionellae могут длительное время выживать в окружающей среде. Они повторно инфицируют Protozoa или ре колонизируют биопленки. По О. Harb et аі. (2000)Взаимоотношения легионелл и Protozoa, нашедшие свое отражение в патогенности этих бактерий для людей, могут быть следующими. Амебы после превращения в цисты, создают бактериям надежное убежище от враждебных условий внешней среды {температура, влажность, дезинфектанты и т. п.). Этим можно объяснить способность легионелл распространяться и населять инженерные системы зданий, прошедшие специальную обработку. По данным J. Barker et аі. (1995), реплицирующаяся в амебах L. pneumophila почти в 1000 раз более резистентна к антимикробным
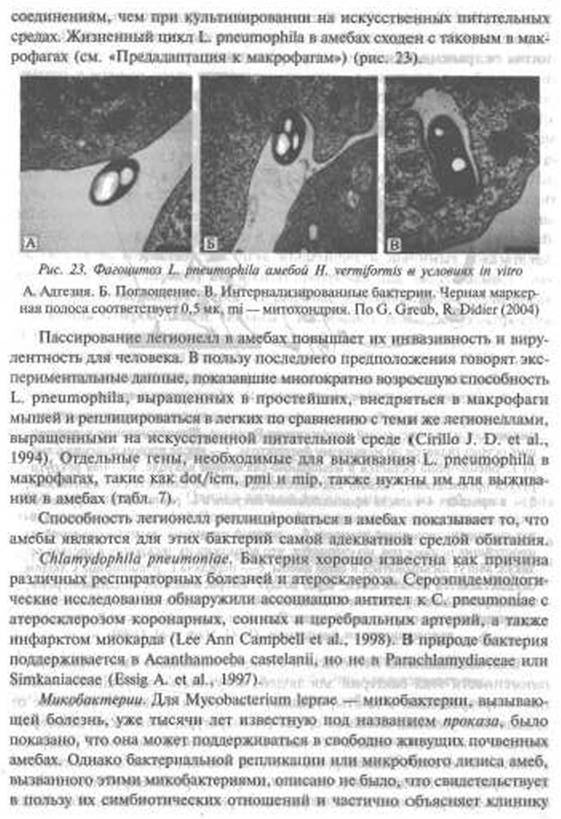
• По О Hart» Cl иі. (2001».


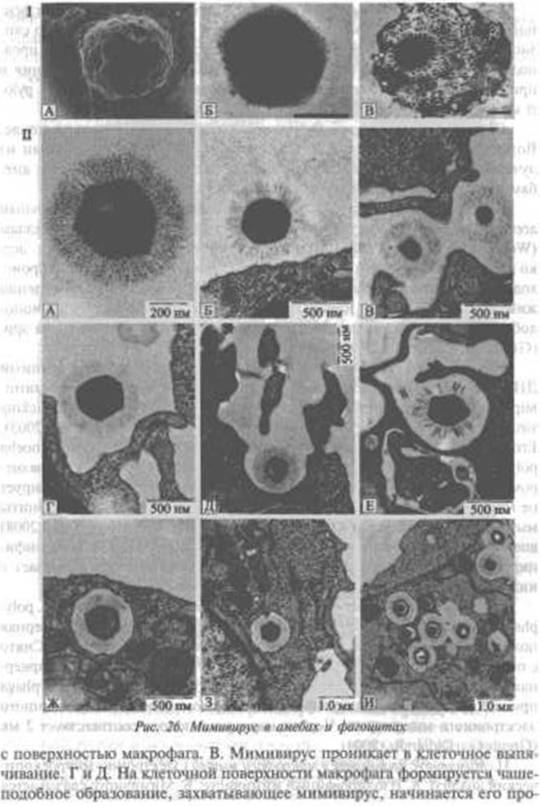
никновенме в клетку. Е. Попошсиие мікрофагом мммшшруса. Ж. Большая гладкая поверхность эндоиитнчсскоЛ вніикулы макрофага, содержащей мішивирус. 3. Везикула, содержащая мпмивирус, глубоко уходит в пито- ісіа іму И- Везикулы, содержащие ынмивирусы, случайно сливаются друг с другом Черные маркерные полосы соответствуют указанным размерам (Ghigo E- ct Bl., 2008).
Таксономически мнмнянрус занимает положение между Poxviridae (к этому таксону относится возбудитель натуральной оспы), Iridoviridac (к ним относятся вирус африканской лихорадки свиней — Afncan wine fever, π многие вирусы земноводных, рыб и насекомых) н Phycodnaviridae Участие мимнпируса в патологии человека пока не установлено (La Scola В et аі, 2003). Е. Ghigo CtaL (2008) выдвинули гипотезу, что APMV способен реплицироваться в альвеолярных макрофагах млекопитающих и вызывать у них пневмонии. Судя по обстоятельности их работы, они скоро ее проверят.
Энтеровирусы Обычно это возбудители KHiue1IHux ннфекпнй у детей, но у некоторых людей они могут вызывать менингоэкисфолкты н асептические менингиты. Передаются между людьми фекально-оральным путем. Широко распространены в морской воле и часто коіггаминируют моллюсков. Их появление в воде носит спорадический характер, но иногда они месяцами могут заражать морскую воду и воду в дельтах рек (Lo S. et аі., 1976). Поэтому была высказана гипотеза, что іи природным резервуаром являются простейшие, живущие в морской воде Однако такая рать простейших в поддержании энтсровнрусов нс была доказана в эксперименте. Вирусы сорбировались на поверхности амебы и нс проникали внутрь клетки и нс размножались в ней. Исследования продолжаются и окончательных выводов пока нс сделано (Greub G.. Didier R., 2004).
/)шпп. В истории к эпидемиологии гриппа многое говорит за его сапронозное происхождение. Поиск среди простейших животных вирусов гриппа нс проводился из-за отсутствия такой постановки задачи (выгоднее бесконечно создавать «вакцину от птичьего гриппа·) Оливко если предположитъ наличие у этих вирусов природного резервуара среди простейших почвы и гкдробиOHTOtl, то проще объяснить следующие особенности гриппозных эпидемий и пандемий
I) ограниченное число подтипов вирусов гриппа А. В и C человека, вызывающих іриппозные эпидемии и пандемии: и как следствие этого феномена — «возвращенке· в человеческие популяции одних и тех же вирусов !напряму, вирус, циркулировавший в 1890-1900 гг., соответствует эталонному штамму А/Сиипшур/1/57/ (H2N2); а вирус, циркулировавший в период C 1900-го по 1917 г. сходен с вирусом Л/Гонконг/І/37/ (H3N2); есть и другие весьма показзтельные примеры такого сходства);
2) отсутствие среди реассортантов вирусов гриппа, циркулировавших среди людей в разных регионах мира, эпидемически значимых вариантов, или вариантов, имеющих селективные преимущества перед вирусами нерекомбинантной природы;
3) доминирование водного механизма передачи возбудителя инфекции в природе, проявляющегося одновременными вспышками гриппозной инфекции у разных таксонов животных, вызванных одним штаммом вируса;
4) появление локальных вспышек гриппозной инфекции у млекопитающих и птиц при отсутствии очевидной связи с «заносом» из других регионов;
5) возможность развития тяжелых форм гриппа при отсутствии пандемии и выделение вирусов гриппа у заболевших людей, не соответствующих по антигенной структуре подтипу вируса, доминирующему в данный период времени (так называемые «асннхронизмы»);
6) отсутствие надежных вирусологических доказательств «положения о непрерывности эпидемического процесса при гриппе» (обычно их подменяют результатами серологических обследований, либо ссылками на «невозможность выделения вируса в межэпидемический период», «снижением вирулентности вируса», наличием «каких-то дефектов вируса, не позволяющих их выделение в обычных лабораторных моделях» и т. п.);
7) консервативность вируса гриппа, выделяемого от дикой водоплавающей птицы;
8) то обстоятельство, что все известные подтипы вируса гриппа А выделены от птиц, причем от одной особи может выделяться несколько се- ровариантов одновременно;
9) способность вируса размножаться во всех тканях, где есть эпителиальная ткань и в фагоцитирующих клетках макроорганизма;
10) «поразительная скорость распространения эпидемий гриппа» — если допустить то, что инфицирование водных источников вирусом гриппа происходит одновременно на обширных территориях, например, в результате изменения экологического равновесия гидробионтов;
11) появление новых серотипов вируса у млекопитающих и домашних птиц как результат «отрыва» вируса от своего природного резервуара и, соответственно, снятия с него селективного давления, существовавшего в природном резервуаре (подробнее см, Супотницкий М. В., 2006),
Э. И. Коренберг (2006) в пользу своей гипотезы сапронозного существования в природе вируса гриппа приводит следующие факты;
• вирусы гриппа А способны длительно сохраняться во внешней среде (в воде месяц при температуре 22 0C и до 6-8 месяцев при 4 °С);
• все известные подтипы вируса гриппа А обнаружены у диких птиц (в основном у водоплавающих и околоводных);
• у водоплавающих птиц грипп протекает бессимптомно и как кишечная инфекция, следовательно, у них нет непосредственной передачи вируса от особи к особи, что могло бы обеспечить его поддержание в природе;
• пандемии гриппа обычно начинаются в теплых регионах Юго-Восточной Азии.
ВИЧ. Попытки установитъ роль Protozoa (Entamoeba histolytica и Giardia lamblia), патогенных обитателей кишечника человека, в распространении ВИЧ, предпринимались еще в начале 1990-х гг., правда, на низком методическом уровне. Иммунологическими методами установлено присутствие ВИЧ в Е. histolytica в течение 48 ч после экспозиции к инфицированным ВИЧ культурам клеток людей (G. lamblia не взаимодействовали с ВИЧ). Но вирус не передавался посредством амеб в неинфицированные клетки. От двух ВИЧ-инфицированных пациентов были выделены ВИЧ-положи- тельные амебы. Их лизировали и полученный лизат добавляли к неинфи- цированным мононуклеарным клеткам крови человека, однако клеток, инфицированных ВИЧ, обнаружено не было (Brown М. et аі., 1991). Эти эксперименты свидетельствуют о том, что ВИЧ способен инфицировать, по крайней мере, отдельные виды простейших. Целесообразно поиски инфицированных ВИЧ простейших продолжить с помощью методов молекулярной диагностики в районах Африки, где население наиболее инфицировано этим вирусом.
Теперь мы можем очертить границы феномена микроорганизмов, способных к сапронозному существованию в природе и одновременно являющихся патогенными для людей. К таким микроорганизмам относятся те из них, которые начинают свое размножение в организме позвоночных в фагоцитирующих клетках. То, что феномен сапронозного существования установлен лишь для отдельных микроорганизмов, опасных для человека и позвоночных животных, во многом связано с тем, что эти исследования только начались.
Признание сапронозного характера существования в природе возбудителей опасных для человека инфекций требует от ученых, с одной стороны, осознания того, что существуют природные очаги инфекционных болезней иные, чем изучаемые сегодня в рамках представлений о природно-очаговых болезнях, когда первичный резервуар возбудителя инфекционной болезни ищут среди позвоночных животных; с другой — необходимость применения новых определений для описания таких очагов и эпидемиологии поддерживающихся в них патогенных микроорганизмов. Подроб-

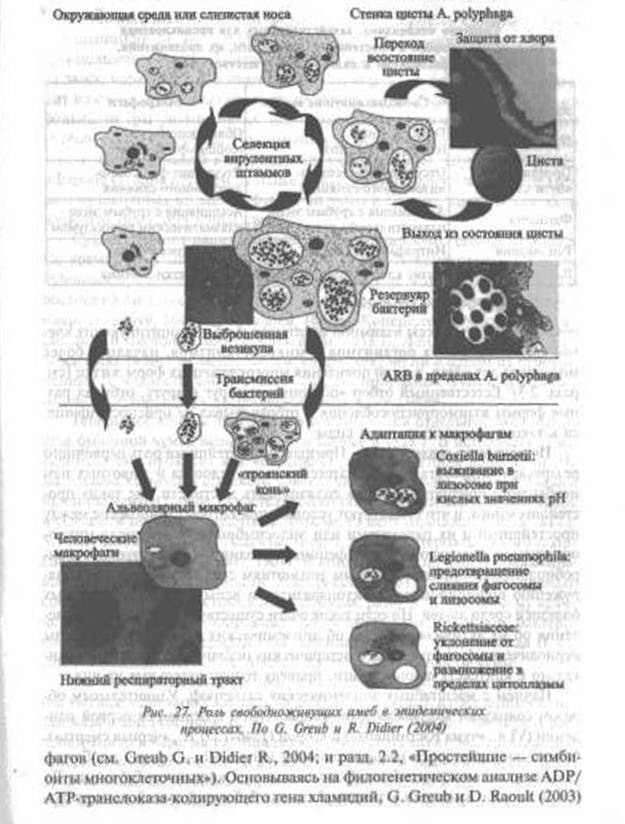
Таблица 8
Сходство механизмов, задействованных для проникновения L- pneumophila в простейшие н макрофаги, их продвижения, репликации и дальнейшего существования*
| Стадия жизненного цикла | Свободноживущие амебы | Макрофаги |
| Проникновение | Обвивающий (соіШщІ-фагшштоз | Обвивающий (соШпг)-фагоцитоз |
| Продвижение внуги клетки | Отсутствие фагосомно- лизосомного слияния | Отсутствие фагосомно- лизосомного слияния |
| Фагосома | Ассоциация с грубым эндоплазматическим ретикулумом | Ассоциация с грубым эндоплазматическим ретикулумом |
| Репликация | Инграфагосомальная | И нтрафагосомальная |
| Выход | Лизис клетки-хозяина | Лизис клетки-хозяина |
* По G. Greub и R. Didier (2004).
показали, что процессы взаимной адаптации амеб, фагоцитирующих клеток многоклеточных организмов и микроорганизмов, начались более миллиарда лет назад, т. е. до появления многоклеточных форм жизни (см. разд. 2.3). Естественный отбор «подгонял» их друг к другу, отбирал разные формы вэаимоприспособления и отбраковывал не приспособившиеся к такому сосуществованию виды.
Исторические свидетельства. Признав за простейшими роль первичного резервуара микроорганизмов, патогенных для человека и животных нам необходимо признать и то, что должны быть местности, где таких простейших много, и что существуют условия, при которых равновесие между простейшими и их паразитами или эндосимбионтами из числа микроорганизмов, нарушаются. Этот феномен сегодня мы фиксируем по микробиологически подтвержденным эпизоотиям среди животных, по обнаружению инфицированных эктопаразитов и вспышкам инфекционных болезней среди людей. Но если такие очаги существуют постоянно, то сведения об их активизации, т. е. об эпидемических катастрофах, должны периодически фиксироваться в исторических источниках. Ниже, применительно к контексту данной книги, приведу только отдельные из них.
Начнем с масштабных эпидемических катастроф. Удивительным образом совпадают местности, охваченные чумой в период ее первой пандемии (VI в., «чума Юстиниана») и второй (1346-1351 гг., «черная смерть»). Совпадают даже сроки распространения пандемий — каждая приблизительно «укладывается» в 5 лет. Обе пандемии развились как бубонные. Во время второй пандемии у многих больных бубонной чумой развивалась вторично-легочная чума; этот феномен более подробно изложен в книге (Супотницкий М. В., Супотницкая Н. C., 2006). Современные объяснения появления чумы в Европе предполагают только одну возможность — ее занос кораблями. Но известно, что, во-первых, бубоная чума не выходит из своих природных очагов и не заносится ни больными людьми, ни больными грызунами, ни их эктопаразитами; во-вторых, классических очагов чумы, например, в понимании Д. К. Заболотного (1926) или таких, которые описаны в современном учебнике Е. П. Шуваловой.(2001), в Европе сегодня нет. Тем не менее существуют исторические свидетельства о чудовищных вспышках бубонной чумы в Европе, когда людям казалось, что «наступил конец света».
Сузим горизонт нашего видения до эпидемий чумы середины XVII в. Со времен «черной смерти» прошло три века. Неконтагиозная болезнь, бубонная чума, вновь «расползается» по Европе в направлении с юга и востока на запад и север и поражает те же города, что и во времена «черной смерти» (Генуя, 1647 г; Барселона, 1653 г.; Копенгаген и Москва, 1654 г.; Неаполь и Силезия, 1656 г.; Амстердам, 1658 г.; Лондон, 1665 г.). В этих эпидемиях гибнет не менее миллиона человек, но ни в одном из перечисленных городов сегодня нет даже «следов» природных очагов чумы в понимании авторов двух вышеприведенных источников.
Теперь посмотрим на «частный случай» — на отдельные обстоятельства бубонной чумы, вспыхнувшей в Одессе в 1812 г. В исторических источниках она проходит как «портовая», т, е. занесенная кораблями. Но материалы комиссии статского советники Н. Трегубова, расследовавшей причины чумы в городе, свидетельствуют об отсутствии заболеваний чумой в Одесском портовом карантине, хотя через него «прошло» за весь 1812 г. 8,5 тыс. человек. Чума началась не в порту, а среди актеров одесского театра (Белиловский В. А. с соавт., 1904), В книге приведена карта, показывающая распространение чумы по Малороссии в 1812 г. Ее зафиксировали среди людей в десятках населенных пунктах, разбросанных по всему краю, не имеющему сегодня природных очагов чумы в традиционном, бактериологическом их подтверждении (Супотницкий М. В., Супотницкая Н. C., 2006). Мы еще многого не знаем о природной очаговости чумы!
Любопытны наблюдения холеры, сделанные локалистами того времени. Исследуя динамику появления холерных вспышек в населенных пунктах Казанской губернии, Н. К. Щепотьев (1890) заметил, что холера распространялась «против течения рек», поэтому он считал, что «извержения холерных больных» не могли служить источником заражения населенных пунктов ниже по течению рек. Он также заметил, что в 1853 г, эпидемии в Демьянском, Ядринском и Чебоксарском уездах начались и прекратились одновременно. Анализ территориального распределения холерных случаев за 30 лет позволил ему установить, что в течение четырех крупных холерных эпидемий (1847-м, 1848-м, 1853-м и 1871 гг.), как правило, болезнь поражала людей в одних и тех же местностях Казанской губернии. Это означает, что холерный вибрион не завозился и не заносился, а существовал в этих местностях, как и полагал Щепотьев, аугохонно.
Теперь обратимся к знаменитому спору Макса Петгенкофера с Робертом Кохом о роли воды в распространении холеры. Не будем сразу считать Петгенкофера ретроградом, а вникнем в его аргументацию. Конта- гионисты во главе с Кохом и Гафки (1887), в подтверждение роли воды в переносе возбудителя холеры, утверждали, что строительство водопровода в Калькутте в 1867 г. привело к резкому снижению заболеваемости населения города холерой. Петгенкофер (локалисты), проанализировав статистические данные по заболеваемости холерой в Калькутте, пришел к выводу, что она имеет пульсирующий характер. Поэтому в течение последних 25 лет холера в Калькутте была значительно слабее, чем в предшествовавшее 25-летие. И как раз слабое проявление болезни совпадает с введением в эксплуатацию водопровода. Поэтому Петгенкофер согласился с тем, что Кох, на первый взгляд, имел некоторое основание приписать именно водопроводу преобладающее значение в ослаблении холеры. Но не более того. Оказалось, что эти скачки холерной смертности — то вверх, то вниз — встречаются как в период времени до 1869 г., так и после него,
Петгенкофер указал на продолжительное снижение заболеваемости холерой в Калькутте в 1847-1850 гг. Холера «держалась» на сравнительно низких цифрах в 1854-м, 1855-м и 1857 гг. Но самое заметное падение холерной кривой начинается с 1864 г., т. е. за 3 года до открытия нового водопровода. В этот год без всякого водопровода холерная смертность была в три раза меньшей, чем в предшествовавшем 1866 г. Минимум холерной смертности в течение этой волны падает на 1871 г,, когда водопроводом было снабжено еще сравнительно небольшое количество домов. Затем, с 1872 г., когда число домов, присоединяющихся к водопроводу, быстро увеличивается, холера снова усиливается и достигает своего максимума в 1876 г., правда, значительно уступающего максимумам прежних периодов. После этого — новое понижение холерной смертности, с минимумом В 1880 г., за которым опятъ следует подъем кривой, максимум которой падает на 1884 г. Холерная волна, начавшаяся с 1880 г., оказалась выше ее предшественницы, занявшей десятилетие с 1871-го по 1881 г. Петгенкофер указал на следующее обстоятельство: «В Калькутте Коху рассказали не всю правду. Верно то, что городской водопровод, разносящий хорошо фильтруемую воду из Ганга, впервые стал снабжать форт Вильяма 25 марта 1873 г,;
но заметное ослабление холеры в крепости началось уже с 1863 г., когда водоснабжение производилось еще из прудов, находящихся на лугах вокруг крепости, Единственная защита прудов от загрязнения состояла из низких деревянньиезаборов, да, пожалуй, еще из стоявших вблизи часовых». Обобщив эти наблюдения, Петтенкофер сделал шуточное заключение, что «параллельно распространению водопровода увеличивается и холера».
Hy а если серьезно, то локалисты пришли к выводу, что «в Калькутте совпадения между водоснабжением и развитием холеры, в котором можно было бы усматривать причинную связь, не существует». Kox был вынужден отвечать оппонентам, так как, с точки зрения его теории о распространении холеры питьевой водой, было непонятно, как это в 1871 г., когда в Калькутте пользовалось новым водоснабжением не более 2000 домов (притом, разумеется, по преимуществу домов в хороших частях города, которые вообще сравнительно свободны от холеры), холера похитила жизни всего 796 человек, тогда как в 1876 г., когда число домов, снабженных водопроводной водой, доходило уже до 10 тыс,, от холеры умерло 2272 человека. Явление это объяснялось Кохом указанием на недостаток водопроводной воды, ощущаемый преимущественно бедной частью населения, вынужденной пользоваться водой из реки и из прудов. Петтенкофер и другие локалисты не приняли объяснение Коха и указали ему на то, что, во-первых, недостаток воды был констатирован официально еще в 1872 г., т. е. как раз в то время, когда кривая холерной заболеваемости стояла очень низко; во-вторых, они привели ему статистику, показывающую, что холерная смертность в предместьях города, вовсе не снабженных водой из водопровода, обнаруживает колебания по времени, соответствующие тем, которые происходят в центре города. И окончательно они «добили» аргументы, приведенные Кохом, используя статистику холерной заболеваемости по всей Бенгальской провинции Индии, в которой тогда жило около 30 млн человек. Оказывается, в 1871-1874 гг. эпидемия холеры была сравнительно слаба, но она снова усилилась в 1876-1877 гг. точно так же, как и в самом городе. Такое же явление повторилось и в 1880 г., когда холерный минимум в Калькутте совпал с таким же минимумом во всей провинции. Если не придумывать эпидемиологию холеры, то ее очень трудно объяснять (более подробно описание пандемий холеры приведено в разд. 4.1.2).
В исторических источниках приводятся описания совпадений эпидемических катастроф, трудно объяснимых с точки зрения контагионистиче- ских представлений. В период с 1918-го по 1922 г. по Поволжью, Прикас- пию, Закавказью, Забайкалью и по разным местностям, прилегающим к границам России, прокатились вспышки чумы, В 1918 г. радикально изменилась ситуация по холере. Вспышки болезни зафиксированы в 40 рос-
сийских губерниях, в Средней Азии, на Кавказе и в Сибири (до Якутска). Холера не прекращалась и в 1919 г. и в 1920 г. даже зимой. Но размах холерной эпидемии 1921 г. был настолько большим, что ее выделили в отдельную эпидемию, сопоставимую по потерям среди населения с наиболее крупными холерными эпидемиями прошлого. Смертность от холеры среди населения составила 55 % от количества заболевших. В некоторых населенных пунктах она достигала 80 и даже 100 %. Летом (!) 1918 г. в Росси вспыхнула пандемия гриппа, названная тогда «испанкой». В период 1918-1922 гг. Россия пережила невиданную в мировой истории эпидемию сыпного и возвратного тифов. Большие и узловые станции железных дорог были забиты трупами людей, умерших от тифа. На некоторых станциях «залежи» трупов достигли огромных размеров, города не успевали открывать тифозные больницы. От «сыпняка» погибла армия адмирала А. В, Колчака. Обычно за сыпным тифом (смертность от 8 до 80 %, что зависело от условий жизни заболевшего) следовал возвратный тиф (смертность до 3 %); тех, кто переживал паразитарные тифы, «прореживали» холера и голод. В 1921 г. внезапно малярия «вышла» далеко за пределы своего обычного распространения, она охватила всю страну и продвинулась даже в ее северные районы. Тяжелые формы тропической малярии, наблюдавшиеся до войны только в предгорьях Кавказа, в Поволжье и в Средней Азии, распространились по всей республике — малярия приняла характер жесточайшего народного бедствия. Ее отличала необычайная смертность — за 1921 г, 11,3 % от числа заболевших первично, но в некоторых группах населения смертность доходила до 80 %.
Эти эпидемические события не поддаются объяснению трудностями гражданской войны и разрухой того времени. Так чума может проникнуть в населенные пункты только после активизации ее природных очагов, на что человек влиять не может. Но активизация таких очагов почему-то происходила в те годы на огромных территориях России и прилегающих к ней стран. Распространение паразитарных тифов в эти годы было отмечено не только в России и в перенесших войну Европейских странах, но и в Латинской Америке.
Для холеры того времени отмечали отсутствие связи ее появления с движениями людских потоков. Например, холеры не было в Красноярске, расположенном на перегруженной холерными больными Транссибирской магистрали, а в Средней Азии ею, как правило, болели только местные жители. Необъясненным осталось повсеместное присутствие в те годы холерного вибриона в источниках питьевой воды даже в северных безлюдных районах Сибири. Например, в реке Иртыш в пределах Тобольска, в реке Туре в пределах Тюмени, в реках Ишиме, Карасуни и Мер- гени в пределах города Ишима Тюменской области (и выше и ниже по реке) и в воде рек и колодцев более умеренных широт.
Напоминание этих фактов должно предостеречь читателя от упрощенного отношения к экологии возбудителей опасных инфекций. Возможно, что при столь значительных различиях между собой по биологическим свойствам (возбудители чумы и холеры — это бактерии; возбудители паразитарных гифов — риккетсии; возбудитель гриппа — РНК-вирус; возбудители малярии — одноклеточные паразиты крови), их экологические ниши на отдельных территориях находятся в зависимости от одних и тех же природных факторов, что и вылилось в чудовищную эпидемическую катастрофу 1918-1922 гг.
* * *
Простейшие, обитатели водных и почвенных экосистем, являются как эволюционными предками макрофагов, так и резервуаром возбудителей инфекционных болезней для многоклеточных организмов, и эти два феномена находятся в причинно-следственной связи друг с другом. Роль простейших в поддержании в природе возбудителей опасных инфекционных болезней людей эмпирически зафиксировал М. Петтенкофер в виде фактора, названного им «фактором Y>>. К микроорганизмам, способным к сапронозному существованию и одновременно являющимся патогенными для людей, относятся те из них, которые начинают свое размножение в организме в фагоцитирующих клетках, т. е, почти все из известных на сегодняшний день. То, что феномен сапронозного существования пока установлен лишь для отдельных микроорганизмов, опасных для человека, означает новизну данного направления исследований и дает шансы исследователям на обнаружение новых природных закономерностей и явлений. Способность многих недавно выявленных микроорганизмов размножаться только в фагоцитирующих клетках, свидетельствует в пользу того, что именно эти клетки, а не искусственные питательные среды современных лабораторий, являются питательной средой, наиболее адекватной их физиологическим потребностям. Нам придется внести очень серьезные изменения в свои представления о физиологии и биохимии микроорганизмов, когда мы начнем их изучать в естественных экосистемах.
Имеющиеся экспериментальные данные по экологии микроорганизмов, эпидемиологические наблюдения и исторические свидетельства позволяют предположить наличие в природе стойких природных очагов возбудителей опасных для людей инфекционных болезней (в том числе и ВИЧ), не подпадающих под рассмотрение современных теорий природно-очаговых болезней и существующих как «подводная часть айсберга» очагов, обнаруживаемых бактериологическими или вирусологическими способами. Границы и организация таких очагов могут быть установлены с помощью методов молекулярной диагностики, но такую задачу ученые еще должны перед собой поставить. Для понятийной дифференциации таких очагов от тех, территориальные границы которых очерчены по эпизоотиям среди диких животных, мы предложили их называть реликтовыми очагами (Супотницкий М. В., 2003, 2004, 2005; Супотницкий М. В., Супот- ницкая Н. C., 2006).
2.2.
Еще по теме Границы феномена сапронозного существования патогенных для людей микроорганизмов.:
- Первичный резервуар патогенных для человека микроорганизмов
- Классификация патогенных для человека микроорганизмов
- Вакцина поликомпонентная из антигенов условно-патогенных микроорганизмов Иммуновак-ВП-4®
- Факторы, влияющие на сохраняемость патогенных микроорганизмов в воде и самоочищение водоемов.
- Использование микроорганизмов для получения БАД, обогащенных микроэлементами
- Использование ассоциации микроорганизмов для очистки сточных вод нефтеперерабатывающей промышленности
- Изучение и использование ассоциаций микроорганизмов для борьбы с нефтяными разливами на воде
- Питательные среды для культивирования микроорганизмов
- Рекомендации по питанию для людей с колостомами
- Ориентация на людей, от людей и против людей
- О РЕЖИМЕ, ОБЩЕМ ДЛЯ ЛЮДЕЙ, ДОСТИГШИХ ВОЗМУЖАЛОСТИ.
- Методические рекомендации по контролю качества питательных сред для микроорганизмов
- Изучение и использование ассоциаций микроорганизмов для борьбы с нефтяными разливами в почве
- Опыление людей без снимания одежды для уничтожения вшей.
- ОТДЕЛ ВТОРОЙ. О РЕЖИМЕ, ОБЩЕМ ДЛЯ ЛЮДЕЙ, ДОСТИГШИХ ВОЗМУЖАЛОСТИ СОСТОИТ ИЗ СЕМНАДЦАТИ ПАРАГРАФОВ
- ГЛАВА X САНИТАРНАЯ ОХРАНА ГРАНИЦ
- Существование и сходимость приближенного решения
- Техника напоминания о недолговечности (бренности) существования.