Фагоцитоз
Одна из важнейших функций лейкоцитов, вышедших из сосудов в очаг воспаления, — фагоцитоз, во время которого лейкоциты распознают, поглощают и разрушают проникшие в организм микроорганизмы, различные чужеродные частицы, а также собственные нежизнеспособные клетки и ткани.
Явление фагоцитоза было открыто И.И. Мечниковым (1845—1916), ион был первым, кто понял значение фагоцитоза как важнейшего механизма невосприимчивости организма к инфекционным заболеваниям. И.И. Мечников ввел общепринятые теперь термины: «фагоцитоз» (по-гречески — пожирание клеток), «фагоцит» (пожиратель клеток) и «макрофаг» (большой пожиратель).Не все лейкоциты, вышедшие в очаг воспаления, способны к фагоцитозу. Такая способность свойственна нейтрофилам, моноцитам, макрофагам и эозинофилам, которые причисляют ктак называемым профессиональным, или облигатным (обязательным), фагоцитам.
В процессе фагоцитоза различают несколько стадий:
1) стадию прилипания (или прикрепления) фагоцита к объекту,
2) стадию поглощения объекта и
3) стадию внутриклеточного разрушения поглощенного объекта.
Прилипание фагоцитов к объекту в отдельных случаях обусловлено
существованием на мембране фагоцитов рецепторов для молекул, входящих в состав микробной стенки (например, для углевода зимозана), или для молекул, появляющихся на поверхности собственных погибающих клеток. Однако в большинстве случаев прилипание фагоцитов к проникшим в организм микроорганизмам осуществляется при участии так называемых опсонинов — сывороточных факторов, которые попадают в очаг воспаления в составе воспалительного экссудата. Опсонины соединяются с поверхностью клетки микроорганизма, после чего к ней легко прилипает мембрана фагоцита. Главными опсонинами являются иммуноглобулины и фрагмент СЗЬ-комплемента. Свойствами опсонинов обладают также некоторые плазменные белки (например, С-реактивный белок) и лизоцим.
Феномен опсонизации можно объяснить тем, что молекулы опсонинов располагают по меньшей мере двумя участками, один из которых связывается с поверхностью атакуемой частички, а другой — с мембраной фагоцита, соединяя таким образом обе поверхности друг с другом. Иммуноглобулины класса G, например, связываются своими Fab-фрагментами с антигенами микробной поверхности, тогда как Fc-фрагменты этих антител — с поверхностной мембраной фагоцитов, на которой имеются рецепторы для Fc-фрагментов IgG.
Процесс поглощения можно рассматривать в известном смысле как продолжение прилипания. В ходе поглощения фагоцит образует псевдоподии, которые окружают объект, прилипая к его покрытой опсонинами поверхности. Когда мембрана псевдоподий покроет всю поверхность 190
объекта, последний оказывается внутри «мешка», образованного мембраной фагоцитирующей клетки. При этом сам мешок, называемый фаго- сомой, оказывается внутри цитоплазмы фагоцита. Дальнейшее разрушение поглощенных частиц происходит внутри фагосомы — вне внутренней среды клетки.
Если речь идет о живых микроорганизмах, а фагоциты способны захватывать живые микроорганизмы, то сначала они должны быть убиты. В лейкоцитах действуют два бактерицидных механизма:
а) зависящий от кислорода и
б) независящий от кислорода.
Зависящий от кислорода бактерицидный фактор связан с образованием активных метаболитов кислорода. Продукция этих веществ начинается после контакта фагоцитов с опсонизированными бактериями. Именно в это время фагоциты, которые в обычных условиях используют энергию анаэробного гликолиза, начинают усиленно поглощать кислород, что обозначают термином респираторный взрыв.
Возникновение респираторного взрыва обусловлено активацией ци- топламатической НАДФН-оксидазы, которая катализирует одноэлектронное восстановление молекулы кислорода до супероксидного радикального аниона, «отбирая» электрон от восстановленного пиридинового нуклеотида НАДФН:
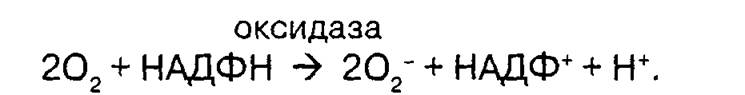
Расходуемые во время «респираторного взрыва» запасы НАДФН начинают немедленно восполняться усиленным окислением глюкозы через гексозомонофосфатный шунт.
Большая часть образующихся при восстановлении О2 супероксидных анионов О2' подвергается дисмутации до Н2О2:
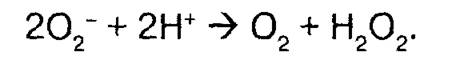
Некоторая часть молекул Н2О2 взаимодействует в присутствии железа или меди с супероксидным анионом с образованием чрезвычайно активного гидроксильного радикала ОН :
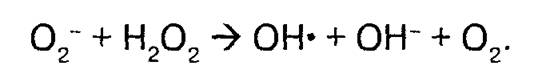
Цитоплазматическая НАДФ-оксидаза активируется в месте контакта фагоцита с микробом, а образование супероксидных анионов происходит на внешней стороне мембраны лейкоцитов, вне внутренней среды клетки. Процесс продолжается и после завершения образования фагосомы, вследствие чего внутри нее создается высокая концентрация бактерицидных радикалов. Проникающие внутрь цитоплазмы фагоцита радикалы нейтрализуются ферментами супероксиддисмутазой и каталазой.
Система образования бактерицидных метаболитов кислорода действует во всех профессиональных фагоцитах. В нейтрофилах совместно с ней действует еще одна мощная бактерицидная система — система 191
миелопероксидазы (сходная с ней пероксидазная система имеется также у эозинофилов, но ее нет у моноцитов и макрофагов).
Миелопероксидаза — фермент, содержащийся в азурофильных гранулах нейтрофилов, катализирует реакцию между ионом галогена (обычно хлора) и перекисью водорода, что приводит к образованию хлорноватистой кислоты (гипохлоритного аниона ОСІ-):
миелопероксидаза

Гипохлорит оказывает выраженное бактерицидное действие сам по себе. Кроме того, он может реагировать с аммонием или аминами, образуя бактерицидные хлорамины.
Независящий от кислорода бактерицидный механизм связан с дегрануляцией — поступлением внутрь фагосомы бактерицидных веществ, которые содержатся во внутриклеточных гранулах фагоцитов.
Когда образование фагосомы завершается, к ней вплотную приближаются гранулы цитоплазмы фагоцитов. Мембрана гранул сливается с мембраной фагосомы, и содержимое гранул вливается внутрь фагосомы. Полагают, что стимулом к дегрануляции является увеличение цитозольного Са2+, концентрация которого возрастает особенно сильно вблизи фагосомы, где располагаются органеллы, накапливающие кальций.
Цитоплазматические гранулы всех облигатных фагоцитов содержат большое количество биологически активных веществ, способных убивать и переваривать микроорганизмы и другие поглощенные фагоцитами объекты. В нейтрофилах, например, имеется 3 типа гранул:
— секреторные пузырьки;
— первичные (азурофильные);
— вторичные (специфические) гранулы.
Наиболее легко мобилизуемые секреторные пузырьки облегчают выход нейтрофилов из сосудов, их миграцию в тканях. Уничтожают и разрушают поглощенные частицы вещества азурофильных и специфических гранул. В азурофильных гранулах, помимо уже упомянутой миелопероксидазы, содержатся действующие независимо от кислорода низкомолекулярные бактерицидные пептиды дефенсины, слабое бактерицидное вещество лизоцим и множество разрушающих ферментов; в специфических гранулах лизоцим и белки, останавливающие размножение микроорганизмов, в частности, лактоферрин, связ’ывающий необходимое для жизнедеятельности микрооргрнизмов железо.
На внутренней мембране специфических и азурофильных гранул находится протонный насос, который переносит водородные ионы из цитоплазмы фагоцита внутрь фагосомы. В результате pH среды в фагосоме понижается до 4—5, что вызывает гибель многих находящихся внутри фагосомы микроорганизмов. После того как микроорганизмы погибают, они разрушаются внутри фагосомы с помощью кислых гидролаз азурофильных гранул.
К числу важных бактерицидных факторов, действующих в активированных макрофагах, следует отнести и продукцию оксида азота (N0), которая осуществляется с помощью индуцибильной NO-синтазы. Фермент этот активируется у-интерфероном, фактором некроза опухолей, ИЛ-1-бета и другими воспалительными цитокинами.
N0 действует цитостатически на опухолевые клетки, бактерии, паразиты, вирусы, ингибируя активность многих ферментов, участвующих в синтезе белков и нуклеиновых кислот. Оксид азота может соединяться с 02*, образуя пероксинитрит, который распадается на цитотоксические свободные радикалы ОН* и N0'.Не все живые микроорганизмы гибнут внутри фагоцитов. Некоторые, например, возбудители туберкулеза сохраняются, оказываясь при этом «отгороженными» мембраной и цитоплазмой фагоцитов от противо- микробных лекарств. -
Активированные хемоаттрактантами фагоциты способны высвобождать содержимое своих гранул не только внутрь фагосомы, но и во внеклеточное пространство. Это происходит во время так называемого незавершенного фагоцитоза — в тех случаях, когда потем или иным причинам фагоцит не может поглотить атакуемый объект, например, если размеры последнего значительно превышают размеры самого фагоцита или если объектом фагоцитоза являются комплексы антиген-антитело, находящиеся на плоской поверхности сосудистого эндотелия. При этом содержимое гранул и продуцируемые фагоцитами активные метаболиты кислорода воздействуют и на объект атаки, и на ткани организма хозяина.
Повреждение тканей хозяина токсичными продуктами фагоцитов становится возможным не только в результате незавершенного фагоцитоза, но и после гибели лейкоцитов или вследствие разрушения мембраны фагосомы самими поглощенными частичками, например частичками кремния или кристаллами мочевой кислоты.
7.5.
Еще по теме Фагоцитоз:
- Учение о фагоцитозе
- Выделяют четыре стадии фагоцитоза:
- Стан фагоцитозу в крові морських свинок у динаміці формування експериментальної пневмонії
- 5.2.4. Препараты, преимущественно воздействующие на показатели врожденного иммунитета (макрофагально-нейтрофильный фагоцитоз, цитотоксичность, интерферонопродукцию).
- Иммунитет
- Метод определения фагоцитарной реакции лейкоцитов крови
- 1. Макрофаги и моноциты.
- УЧЕНИЕ О ПАРАЗИТИЧЕСКИХ ПРОСТЕЙШИХ
- ПОДЦАРСТВО PROTOZOA ТИП SARCOMASTIGOPHORA КЛАСС LOBOSEA
- НЕДОСТАТОЧНОСТЬ ЗАЩИТНОЙ РЕАКЦИИ ФАГОЦИТОЗА КАК ПРИЧИНА ИММУНОДЕФИЦИТА
- Проявление активности опухолевых клеток
- 2.2.2 Антитела
- 2.2 Роль опсонинов в иммунном ответе при криптококкозе
- Поглотительная активность фагоцитов.
- 13.3. Иммунодиагностика аутоиммунных заболеваний