хНМНТНПШ
кХОХДМШИ АХЯКНИ ОПХПНДМШУ ЛЕЛАПЮМ ОПЕДЯРЮБКЪЕР ЯНАНИ МЕОПНМХЖЮЕЛШИ АЮПЭЕП ДКЪ ЯБНАНДМНИ ДХТТСГХХ ХНМНБ, НАКЮ╜ДЮЧЫХУ МХГЙНИ КХОНТХКЭМНЯРЭЧ ХГ-ГЮ ЯХКЭМНИ ЦХДПЮРЮЖХХ. хНМНТНПШ ОПЕДМЮГМЮВЕМШ ДКЪ РНЦН, ВРНАШ НАКЕЦВХРЭ РПЮМЯОНПР ХНМНБ ВЕПЕГ ЩРХ ЛЕЛАПЮМШ.
яСЫЕЯРБСЧР ДБЮ РХОЮ ХНМНТНПНБ: ОНДБХФМШЕ Х ЯРЮЖХНМЮПМШЕ. оНЯКЕДМХЕ (╚ХНММШЕ МЮЯНЯШ╩) ОПЕДЯРЮБКЪЧР ЯНАНИ ОПНМХГШБЮЧЫХЕ АХЯКНИ ЙЮМЮКШ, МЮОНКМЕМ╜МШЕ БНДНИ. лМНЦНЕ ХГ РНЦН, ВРН ХГБЕЯРМН Н ХНМНТНПЮУ, СЯРЮМНБ╜КЕМН ОПХ ХГСВЕМХХ ОНДБХФМШУ РХОНБ ХНМНТНПНБ, БШДЕКЕММШУ ХГ ЛХЙПНАНБ (МЮОПХЛЕП, БЮКХМНЛХЖХМ, ЦПЮЛХЖХДХМ). рЮЙХЕ ЩЙГН╜ЦЕММШЕ ХНМНТНПШ, ЕЯКХ НМХ ЩТТЕЙРХБМШ, БШЯНЙНРНЙЯХВМШ ДКЪ ЙКЕРНЙ ЛКЕЙНОХРЮЧЫХУ, МН МЕ Б МХГЙХУ ДНГЮУ.уНРЪ ДХСПЕРХЙХ Х ЯЕПДЕВМШЕ ЦКХЙНГХДШ РЮЙ ХКХ ХМЮВЕ ЯБЪ╜ГЮМШ Я ОЕПЕМНЯНЛ ХНМНБ, ХУ МЕ ЯКЕДСЕР ПЮЯЯЛЮРПХБЮРЭ ЙЮЙ ХНМН╜ТНПШ. дЮФЕ хЮд-юртЮГЮ НЯСЫЕЯРБКЪЕР КХЬЭ БЯОНЛНЦЮРЕКЭМНЕ ДЕИЯРБХЕ. рЮЙ, Б МЮРПХЕБНЛ МЮЯНЯЕ ЩРНР ТЕПЛЕМР, ЯБЪГЮММШИ Я ЛЕЛАПЮМНИ, ХЯОНКЭГСЕРЯЪ ДКЪ БШБЕДЕМХЪ МЮРПХЪ Х ББЕДЕМХЪ ЙЮ╜КХЪ Б ЙКЕРЙС. щМЕПЦХЪ, МЕНАУНДХЛЮЪ ДКЪ ОПНРЕЙЮМХЪ ЩРНЦН ОПН╜ЖЕЯЯЮ, БШДЕКЪЕРЯЪ ОПХ ЦХДПНКХГЕ юрт. тЕПЛЕМР ЯБЪГШБЮЕР Х БШЯБНАНФДЮЕР ЙЮРХНМШ МЮ ПЮГМШУ ЯРЮДХЪУ ЖХЙКЮ ПЮАНРШ ХНММН╜ЦН МЮЯНЯЮ. йНМТНПЛЮЖХНММШЕ ХГЛЕМЕМХЪ БН БПЕЛЪ ЖХЙКЮ ОПХБН╜ДЪР Й РНЛС, ВРН ЙЮРХНМ, БНИДЪ Я НДМНИ ЯРНПНМШ ЛЕЛАПЮМШ, БШ╜ДЕКЪЕРЯЪ МЮ ОПНРХБНОНКНФМНИ. вЮЯРХВМН ЛЕУЮМХГЛ ГЮЙКЧВЮЕРЯЪ Б ЯКЕДСЧЫЕЛ: ЦХДПНКХГ юрт БШГШБЮЕР МЮРПХИГЮБХЯХЛНЕ ТНЯ╜ТНПХКХПНБЮМХЕ ЮЯОЮПРХКЭМНИ ЦПСООШ ТЕПЛЕМРЮ, ЯНОПНБНФДЮЧ╜ЫЕЕЯЪ ЙЮКХИГЮБХЯХЛШЛ ОЕПЕМНЯНЛ ЩРНИ ТНЯТНПХКЭМНИ ЦПСООШ ТНЯТЮРЮГНИ НР ЮЯОЮПРЮРЮ Б БНДС. яСЫЕЯРБСЕР ЛМЕМХЕ Н РНЛ, ВРН ОН ЙПЮИМЕИ ЛЕПЕ С МЕЙНРНПШУ БХДНБ БЮМЮДЮР-ЮМХНМ, ЙНМЙСПХ╜ПСЧЫХИ Я юрт ГЮ ЛЕЯРЮ ЯБЪГШБЮМХЪ, ОПХМХЛЮЕР СВЮЯРХЕ Б ТХ╜ГХНКНЦХВЕЯЙНИ ПЕЦСКЪЖХХ ЩРНЦН ТЕПЛЕМРЮ. пНДЯРБЕММШИ ТЕПЛЕМР яЮ,л╖-юртЮГЮ БШЯРСОЮЕР Б ПНКХ ЯНБЕПЬЕММН ЮМЮКНЦХВМНЦН МЮЯНЯЮ, МЮЦМЕРЮЧЫЕЦН ЙЮРХНМШ ЛЮЦМХЪ Б ЙКЕРЙС Х БШБНДЪЫЕЦН ЙЮКЭЖХИ. оПХПНДЮ ХНМНТНПНБ ОНДБХФМНЦН РХОЮ С ЛКЕЙНОХРЮЧ╜ЫХУ ХГСВЕМЮ ЯКЮАН.
б ОПНРХБНОНКНФМНЯРЭ ОПНЖЕЯЯС УЕКЮРХПНБЮМХЪ (ПЮГД. 11),. ОПХ ЙНРНПНЛ ЮРНЛ БНДНПНДЮ КХЦЮМДЮ ГЮЛЕЫЮЕРЯЪ МЕНПЦЮМХВЕ╜ЯЙХЛ ЙЮРХНМНЛ, РПЮМЯОНПР ОНЯКЕДМХУ ХНМНТНПЮЛХ МСФДЮЕРЯЪ КХЬЭ Б ОНЯРЮДХИМНЛ ГЮЛЕЫЕМХХ ХНМНТНПНЛ ЛНКЕЙСК БНДШ, БЙКЧВЕММШУ Б ЙННПДХМЮЖХНММСЧ ЯТЕПС ЙЮРХНМЮ. щМЕПЦХЪ ДЕ╜ЯНКЭБЮРЮЖХХ ЙНЛОЕМЯХПСЕРЯЪ ЩМЕПЦХЕИ ЯБЪГШБЮМХЪ ХНХНТНПЮ. б ОПНЖЕЯЯЕ РЕОКНБНЦН ДБХФЕМХЪ ЯНКЭБЮРХПНБЮММШИ ЙЮРХНМ ЯМЮ╜ВЮКЮ ЯРЮКЙХБЮЕРЯЪ Я ОНКЪПМШЛХ ЦПСООЮЛХ КХЦЮМДЮ-ХНМНТНПЮ, ЙНРНПШИ ГЮРЕЛ НАБНКЮЙХБЮЕР ЙЮРХНМ, Х НАПЮГСЧЫХИЯЪ ЙНЛОКЕЙЯ, ХЛЕЧЫХИ КХОНТХКЭМСЧ БМЕЬМЧЧ ЯРНПНМС, ОНЦКНЫЮЕРЯЪ ЖХРН╜ОКЮГЛЮРХВЕЯЙНИ ЛЕЛАПЮМНИ. йНМТНПЛЮЖХНММШЕ ХГЛЕМЕМХЪ, ЯН╜ОПНБНФДЮЧЫХЕ ГЮУБЮР ЙЮРХНМЮ, ОПНРЕЙЮЧР НВЕМЭ АШЯРПН, МН ЛНЦСР АШРЭ НАМЮПСФЕМШ (ДКЪ БЮКХМНЛХЖХМЮ) Я ОНЛНЫЭЧ ПЕКЮЙ╜ЯЮЖХНММШУ ЛЕРНДНБ (МЮОПХЛЕП, АШЯРПШУ РЕЛОЕПЮРСПМШУ ЯЙЮВЙНБ ХКХ ОНЦКНЫЕМХЪ ГБСЙНБШУ БНКМ) [Grell, Eggers, Funck, 1972].
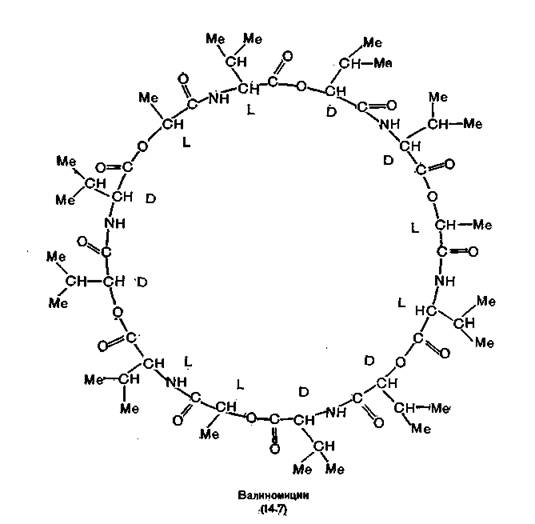
мЕЙНРНПШЕ ЛЕРЮАНКХРШ ЦПХАНБ ОНБШЬЮЧР ОЮЯЯХБМШИ ГЮУБЮР ЙЮКХЪ. нДМХ ХГ МХУ, МЮОПХЛЕП ХНМЮЙРХМ Х ЛНМЮЙРХМ (14.6), НР╜МНЯЪРЯЪ Й ЙКЮЯЯС ЛЮЙПНРЕРПНКХДНБ (ЛМНЦНГБЕММШУ ЖХЙКХВЕЯЙХУ ЯКНФМШУ ЩТХПНБ), ДПСЦХЕ (БЮКХМНЛХЖХМ)≈Й ЙКЮЯЯС ДЕОЯХОЕО- РХДНБ. бЮКХМНЛХЖХМ (14.7) (ХГ Streptomyces fulvissimus) ОПЕД╜ЯРЮБКЪЕР ЯНАНИ 36-ВКЕММШИ ЛЮЙПНЖХЙК, БЙКЧВЮЧЫХИ ОН РПХ НЯРЮРЙЮ L-БЮКХМЮ, D-БЮКХМЮ, L-ЛНКНВМНИ ЙХЯКНРШ Х НЯРЮРНЙ a-D-ЦХДПНЙЯХХГНБЮКЕПХЮМНБНИ ЙХЯКНРШ, ЯБЪГЮММШУ ЯКНФМНЩТХП╜МШЛХ Х ЮЛХДМШЛХ ЯБЪГЪЛХ [Shemyakin et al., 1963]. нМ ПЮЯР╜БНПХЛ Б КХОХДЮУ Х МЕПЮЯРБНПХЛ Б БНДЕ. оЕПХЛЕРП ЩРНИ ОКНЯЙНИ ЛНКЕЙСКШ КХОНТХКЕМ, РНЦДЮ ЙЮЙ БМСРПЕММЪЪ ВЮЯРЭ ЙНКЭЖЮ БШЯР╜КЮМЮ МЕГЮПЪФЕММШЛХ ЦХДПНТХКЭМШЛХ ЦПСООЮЛХ. б ПЕГСКЭРЮРЕ ЙНМТНПЛЮЖХНММШУ ХГЛЕМЕМХИ, БШГБЮММШУ ГЮУБЮРНЛ ЙЮРХНМЮ ЙЮ╜КХЪ, ЛНКЕЙСКЮ ОПХНАПЕРЮЕР АПЮЯКЕРННАПЮГМСЧ ТНПЛС.
бЮКХМНЛХЖХМ ЯОНЯНАЯРБСЕР ОПНМХЙМНБЕМХЧ ХНМНБ ЙЮКХЪ Б ЩПХРПНЖХРШ, ЛХРНУНМДПХХ [Moore, Pressman, 1964] Х ВЕПЕГ АЮЙ╜РЕПХЮКЭМШЕ ОКЮГЛЮРХВЕЯЙХЕ ЛЕЛАПЮМШ [Harold, Baarda, 1967], МН ОНВРХ МЕ БКХЪЕР МЮ ОПНМХЙМНБЕМХЕ ХНМНБ МЮРПХЪ, КХРХЪ Х БН╜ДНПНДЮ. мНМЮЙРХМ НАКЮДЮЕР ОНДНАМШЛ ДЕИЯРБХЕЛ, Ю ЛНМНЮЙРХМ, ЙПНЛЕ РНЦН, ЯКЕЦЙЮ ОНБШЬЮЕР ОПНМХЖЮЕЛНЯРЭ ДКЪ ХНМНБ МЮРПХЪ [Henderson, McGivan, Chappell, 1969].
бЮКХМНЛХЖХМ ОПНЪБКЪЕР ОПНРХБННОСУНКЕБШЕ ЯБНИЯРБЮ (МЮ ЛШЬЮУ) [Carter, Sakurai, Umezawa, 1981].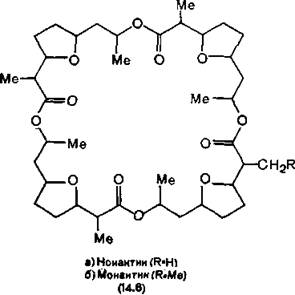
пЕМРЦЕМНЯРПСЙРСПМШИ ЮМЮКХГ ОНЙЮГЮК, ВРН ХНМШ ЙЮКХЪ ЙННП╜ДХМХПСЧР Я ВЕРШПЭЛЪ ЩТХПМШЛХ Х ВЕРШПЭЛЪ ЙЮПАНМХКЭМШЛХ ЮРНЛЮЛХ ЙХЯКНПНДЮ МНМЮЙРХМЮ [Dobler, Dunitz, Kilbourn, 1969] Х Я ЬЕЯРЭЧ ЮРНЛЮЛХ ЙХЯКНПНДЮ ЙЮПАНМХКЭМШУ ЦПСОО БЮКХМНЛХ- ЖХМЮ. рЮЙХЛ НАПЮГНЛ, ПЮМЕЕ ЦХДПЮРХПНБЮММШИ ХНМ ЙЮКХЪ ОПХ╜НАПЕРЮЕР БМЕЬМЧЧ КХОНТХКЭМСЧ НАНКНВЙС Х ОНЩРНЛС ЯОНЯНАЕМ
ДНЯРЮРНВМН КЕЦЙН ОПНМХЙЮРЭ Б ЙКЕРНВМШЕ ЛЕЛАПЮМШ. оПХ ЯРНКЙ╜МНБЕМХХ ЩРНЦН ЙНЛОКЕЙЯЮ Я МЮПСФМНИ ЦПЮМХЖЕИ ЛЕФДС ЛЕЛАПЮ╜МНИ Х БНДНИ ВЮЯРЭ ХНМНБ ЙЮКХЪ ОПНУНДХР ВЕПЕГ МЕЕ, Ю ДПСЦЮЪ ВЮЯРЭ НАЛЕМХБЮЕР ЛНКЕЙСКС МНМЮЙРХМЮ МЮ БНДС (ЛЕДКЕММЮЪ ЯРЮ╜ДХЪ ОЕМЕРПЮЖХХ). б ЩРНЛ ДБХФЕМХХ ЮМХНМ (МЮОПХЛЕП, я1~) ДНКФЕМ ЯНОПНБНФДЮРЭ й+, МН МЮ МЕАНКЭЬНЛ ПЮЯЯРНЪМХХ Х ОН╜ЯРНЪММН ОНДБЕПЦЮЪЯЭ НАЛЕМС. оПЕДОНВРХРЕКЭМНЯРЭ ГЮУБЮРЮ МН- МЮЙРХМНЛ ХНМНБ ЙЮКХЪ, Ю МЕ МЮРПХЪ, НОПЕДЕКЪЕРЯЪ ЯЙНПЕЕ АНКЕЕ МХГЙНИ ЩМЕПЦХЕИ ДЕЦХДПЮРЮЖХХ ОЕПБНЦН, ВЕЛ ПЮГЛЕПЮЛХ БМСРПЕМ╜МЕИ ОНКНЯРХ ЖХЙКХВЕЯЙНИ ЛНКЕЙСКШ [Prestegard, Chan, 1970].
я ОНЛНЫЭЧ ЮКЮЛЕРХЖХМЮ ≈ КХМЕИМНЦН ОЕОРХДЮ, БШДЕКЕММНЦН ХГ ЦПХАНБ Trichoderma viride, ЛНФМН СЯРЮМНБХРЭ, ЙЮЙ НАПЮГСЧР╜ЯЪ ЙЮКХЕБШЕ ЙЮМЮКШ. щРНР ОЕОРХД, ЙПНЛЕ РЕПЛХМЮКЭМНЦН L-ТЕ- МХКЮКЮМХМНКЮ, ЯНДЕПФХР 20 ЮЛХМНЙХЯКНРМШУ НЯРЮРЙНБ, Б РНЛ ВХЯКЕ БНЯЕЛЭ НЯРЮРЙНБ Ю-ЮЛХМНЛЮЯКЪМНИ ЙХЯКНРШ, Ю ДПСЦХЕ ≈ НАШВМШЕ ЮЛХМНЙХЯКНРШ, БУНДЪЫХЕ Б ЯНЯРЮБ АЕКЙНБ. дЮММШЕ ПЕМРЦЕМНЯРПСЙРСПМНЦН ЮМЮКХГЮ ЙПХЯРЮККНБ ЩРНЦН ЯНЕДХМЕМХЪ ЯБХДЕРЕКЭЯРБСЧР Н РНЛ, ВРН ЦХДПНТХКЭМШЕ ТПЮЦЛЕМРШ МЮОПЮБ╜КЕМШ Б НДМС ЯРНПНМС, Ю КХОНТХКЭМШЕ ≈ Б ДПСЦСЧ.
21-734
321
я ОНЛНЫЭЧ ЛНКЕЙСКЪПМШУ ЛНДЕКЕИ СДЮКНЯЭ ОНЙЮГЮРЭ, ВРН ЩРН ЯНЕДХМЕМХЕ ЛНФЕР НАПЮГНБШБЮРЭ ЮЦПЕЦЮРШ, ОПХВЕЛ ЙЮФДШЕ БНЯЕЛЭ ЛНКЕЙСК НАПЮГСЧР ЦХДПНТХКЭМШЕ ОНПШ ДХЮЛЕРПНЛ НЙНКН 0,45 МЛ, ЙНРНПШЕ ЛНЦСР АШРЭ АКНЙХПНБЮМШ ЦКСРЮЛХМНБШЛ НЯРЮР╜ЙНЛ (Gln-7) Б НРБЕР МЮ ХГЛЕМЕМХЪ ДХОНКЭМНЦН ЛНЛЕМРЮ.
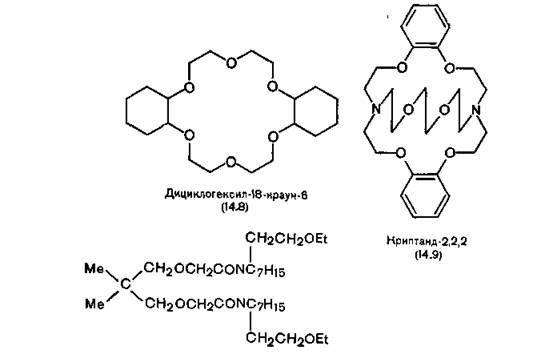
я СБЕ╜ПЕММНЯРЭЧ ЛНФМН ЯЙЮГЮРЭ, ВРН ЮКЮЛЕРХЖХМ ЯЮЛ ЯОНМРЮММН БЙКЧ╜ВЮЕРЯЪ Б КХОХДМШЕ ЛЕЛАПЮМШ Х ЯЙБНГЭ ЕЦН ОНПШ, НРЙПШРШЕ БНПНРМШЛ ОНРЕМЖХЮКНЛ, МЮВХМЮЧР ОПНУНДХРЭ МЕНПЦЮМХВЕЯЙХЕ ЙЮРХНМШ [Fox, Richards, 1982].оПНЯРШЕ ЯХМРЕРХВЕЯЙХЕ Х МЕДНПНЦХЕ ЖХЙКХВЕЯЙХЕ ОНКХЩТХПШ, МЮГШБЮЕЛШЕ ╚ЙПЮСМ-ЩТХПЮЛХ╩, ЯОНЯНАМШ ОЕПЕМНЯХРЭ ЙЮРХНМШ ЫЕКНВМШУ Х ЫЕКНВМН-ГЕЛЕКЭМШУ ЛЕРЮККНБ ХГ БНДМНЦН ЯКНЪ Б КХОНТХКЭМШИ Я НВЕМЭ БШЯНЙНИ ХГАХПЮРЕКЭМНЯРЭЧ. нДМН ХГ МЮХ╜АНКЕЕ ЮЙРХБМШУ ЯНЕДХМЕМХИ ≈ ДХЖХЙКНЦЕЙЯХК-18-ЙПЮСМ-6 (14.8), ОПЕДЯРЮБКЪЕР ЯНАНИ ЙНКЭЖН, Б ЙНРНПНЛ ДБЕ ЛНКЕЙСКШ 1,2-ДХЦХД╜ПНЙЯХЖХЙКНЦЕЙЯЮМЮ ЯБЪГЮМШ ДБСЛЪ ЛНКЕЙСКЮЛХ ДХЩРХКЕМЦКХЙН╜КЪ. йНЛОКЕЙЯННАПЮГСЧЫЮЪ ЯОНЯНАМНЯРЭ ЙПЮСМ-ЩТХПНБ НРМНЯХ╜РЕКЭМН МЕБЕКХЙЮ, МЕЯЛНРПЪ МЮ РН, ВРН ЙЮРХНМ БГЮХЛНДЕИЯРБСЕР Я МЕЯЙНКЭЙХЛХ ЮРНЛЮЛХ ЙХЯКНПНДЮ НДМНБПЕЛЕММН, МЮОПХЛЕП Я ЬЕЯРЭЧ Б ЩТХПЕ (14.8) [Izatt et al., 1971]. щРНР ЙПЮСМ-ЩТХП ЯБЪГШБЮЕР ОПЕХЛСЫЕЯРБЕММН ЙЮКХИ, Ю МЕ МЮРПХИ; ЩТХП Я ЛЕМЭ╜ЬХЛ ВХЯКНЛ ЮРНЛНБ ЙХЯКНПНДЮ ≈ МЮРПХИ, Ю ЕЫЕ Я ЛЕМЭЬХЛ ≈ КХРХИ. оПХ ГЮУБЮРЕ ЯНКЕИ ЩРХ ЛНКЕЙСКШ ВЮЯРН ХГЛЕМЪЧР ЯБНЧ ЙНМТНПЛЮЖХЧ [Pedersen, 1970].
бЯКЕД ГЮ НРЙПШРХЕЛ Pedersen ЙПЮСМ-ЩТХПНБ Б яью, БН тПЮМЖХХ МЮВЮКХ ПЮАНРЮРЭ Я ЙПХОРЮМДЮЛХ. щРХ ЯЛЕЬЮММШЕ ЛЮЙ- ПНЛНКЕЙСКЪПМШЕ ЮЛХМНЩТХПШ МЕ ОПНЯРН БЙКЧВЮЧР ЙЮРХНМ Б ЖХЙК ОНДНАМН ЙПЮСМ-ЩТХПЮЛ, МН ОПНВМН ЯБЪГШБЮЧР ЕЦН ОН БЯЕЛ МЮОПЮБКЕМХЪЛ [Lehn, Sauvage, Dietrich, 1970]. рЮЙ, ЯНЕДХМЕМХЕ (14.9) ЪБКЪЕРЯЪ ЛНЫМШЛ ЙНЛОКЕЙЯННАПЮГНБЮРЕКЕЛ ЙЮРХНМНБ ЙЮ╜КХЪ. х ЙПЮСМ-ЩТХПШ, Х ЙПХОРЮМДШ МЕ МЮЬКХ ЬХПНЙНЦН ОПХЛЕ╜МЕМХЪ Б АХНКНЦХХ ХГ-ГЮ БШЯНЙНИ РНЙЯХВМНЯРХ. яНЕДХМЕМХЕ (14.9), МЮОПХЛЕП, НАКЮДЮЕР ПЮГДПЮФЮЧЫХЛ ДЕИЯРБХЕЛ МЮ ЦКЮГЮ Х ЙНФС, Ю ЕЦН БШЯЬХЕ ЦНЛНКНЦХ ОНПЮФЮЧР жмя [Pedersen, 1972]. х БЯЕ- РЮЙХ ЩРХ ЯНЕДХМЕМХЪ НВЕМЭ ОНКЕГМШ ≈ ХУ ЛНФМН ПЮЯЯЛЮРПХБЮРЭ ЙЮЙ ЛНДЕКХ ЛМНЦНВХЯКЕММШУ Х НВЕМЭ ДНПНЦХУ ОПХПНДМШУ ХНМН╜ТНПНБ. аНКЭЬХМЯРБН МЮХАНКЕЕ КХОНТХКЭМШУ ЙПЮСМ-ЩТХПНБ ХКХ ЙПХОРЮМДНБ ЯБЪГШБЮЧР ОПЕХЛСЫЕЯРБЕММН НДМНБЮКЕМРМШЕ, Ю МЕ ДБСУБЮКЕМРМШЕ ЙЮРХНМШ, Р. Е. МЮРПХИ ХЛЕЕР ОПЕХЛСЫЕЯРБН ОЕПЕД ЙЮКЭЖХЕЛ, МЕЯЛНРПЪ МЮ РН, ВРН ПЮГЛЕПШ ЩРХУ ЙЮРХНМНБ Б МЕЦХД- ПЮРХПНБЮММНЛ ЯНЯРНЪМХХ ОНВРХ НДХМЮЙНБШ.
йПЮСМ-ЩТХПШ АШКХ ХЯОНКЭГНБЮМШ Б ЙЮВЕЯРБЕ ЛНДЕКХ РПЮМЯ╜ОНПРЮ ЮМХНМНБ ЯЙБНГЭ ЛЕЛАПЮМШ ОПНРХБ ЦПЮДХЕМРЮ ЙНМЖЕМРПЮЖХХ (ЯП. Я РПЮМЯОНПРНЛ РХОЮ 2, ПЮГД. 3.2.2). рПЮМЯОНПРХПСЕЛШЛХ ЮМХНМЮЛХ АШКХ N-АЕМГНХКХПНБЮММШЕ ЮЛХМНЙХЯКНРШ Х ЙНПНРЙХЕ ОЕОРХДШ; ЛЕЛАПЮМЮ ОПЕДЯРЮБКЪКЮ ЯНАНИ ОЕПЕЛЕЬХБЮЕЛШИ ЯКНИ УКНПНТНПЛЮ, Я НАЕХУ ЯРНПНМ ЯНОПХЙЮЯЮЧЫХИЯЪ Я БНДНИ. рЮЙ ЙЮЙ ЙПЮСМ-ЩТХПШ ЛЮКНПЮЯРБНПХЛШ Б БНДЕ, НМХ МЮЙЮОКХБЮКХЯЭ Б УКНПНТНПЛЕ. й ОЕПБНИ ЦПЮМХЖЕ ПЮГДЕКЮ БНДЮ ≈ УКНПНТНПЛ АШКХ
ОНДБЕДЕМШ ХНМШ ЙЮКХЪ. щРН ОПХБЕКН Й ЩЙЯРПЮЙЖХХ ХНМНБ Б УКН╜ПНТНПЛЕММШИ ЯКНИ Я НАПЮГНБЮМХЕЛ РПНИМНЦН ЙНЛОКЕЙЯЮ (ЮМХНМ,
кХРХЕБШИ ХНМНТНП (14.10)
й+, ХНМНТНП), ХГ ЙНРНПНЦН НАЮ ХНМЮ БШДЕКЪКХЯЭ БН БРНПСЧ БНД╜МСЧ ТЮГС (ГЮ ЛЕЛАПЮМНИ). нЯБНАНФДЕММШИ ХНМНТНП ЯМНБЮ ГЮ╜УБЮРШБЮК ЮМХНМ Х ЙЮРХНМ ХГ ОЕПБНИ БНДМНИ ТЮГШ, Р. Е. ЖХЙК ОНБРНПЪКЯЪ [Tsukube, 1982]. н ЙНЛОКЕЙЯННАПЮГНБЮМХХ ЮМХНМНБ ЙПХОРЮМДЮЛХ ЯЛ. Dietrich Х ЯНРП. (1978).
юЖХЙКХВЕЯЙХИ ХНМНТНП (14.10) ДКЪ ЙЮРХНМНБ КХРХЪ НАПЮГСЕР БНЙПСЦ ХНМЮ КХОНТХКЭМСЧ ЦЕЙЯЮДЕМРЮРМСЧ НАНКНВЙС ХНМЮ, ХЛЕ╜ЧЫСЧ НЙРЮЩДПХВЕЯЙСЧ ЯРПСЙРСПС. оПЕДОНВРХРЕКЭМНЯРЭ ГЮУБЮРЮ ЙЮРХНМЮ КХРХЪ Б 40 ПЮГ БШЬЕ ОН ЯПЮБМЕМХЧ Я ЙЮРХНМНЛ МЮРПХЪ. щРН ЯНЕДХМЕМХЕ ОПХ БМСРПХБЕММНЛ ББЕДЕМХХ ОНБШЬЮЕР ГЮУБЮР КХРХЪ ЛНГЦНЛ ЙПШЯ [Shanzer, Samuel, Korenstein, 1983].
еЫЕ ОН РЕЛЕ хНМНТНПШ:
- юМХНММШЕ ХНМНТНПШ.
- юРХАЮЙРЕПЁЮКЭМЮ РЮ ОПНРХЦПХАЙНБЮ РЕПЮОЁЪ Б ОЕДЁЮРПЁ©: мЮБВЮКЭМН-ОПЮЙРХВМХИ ОНЯЁАМХЙ 11 БХДЮММЪ / гЮ ПЕД. ОПНТ. б.б. аЕ╜ПЕФМНЦН. - уЛЕКЭМХЖЭЙХИ,2016. - 416 Я., 2016
- релю ╧ 31 цемепюкхгнбюммше онякепнднбше хмтейжхнммше гюанкебюмхъ кюйрюжхнммши люярхр яеорхвеяйхи ьнй б юйсьепярбе
- релю ╧ 30 цмнимн-бняоюкхрекэмше онякепнднбше гюанкебюмхъ
- релю ╧ 29 меопюбхкэмне онкнфемхе окндю ноепюжхх, хяопюбкъчыхе онкнфемхъ окндю. юйсьепяйхе онбнпнрш хгбкевемхе окндю гю рюгнбши йнмеж
- релю ╧ 28 окнднпюгпсьючыхе ноепюжхх люкше юйсьепяйхе ноепюжхх
- релю ╧ 27 юйсьепяйхе ыхожш х бюйссл-щйярпюйжхъ
- релю ╧ 26 йеяюпебн яевемхе
- релю ╧ 25 юмнлюкхх пнднбшу яхк
- релю ╧ 24 оепемюьхбюмхе аепелеммнярх опефдебпелеммше пндш
- релю ╧ 23 хллсмнкнцхвеяйюъ меянблеярхлнярэ лефдс люрепэч х окнднл (МЮ ОПХЛЕПЕ Rh-ЯЕМЯХАХКХГЮЖХХ Х Rh-ЙНМТКХЙРЮ
- релю ╧ 22 пнднбни рпюблюрхгл люрепх
- релю ╧ 20 окюжемрюпмюъ меднярюрнвмнярэ цхонйяхъ окндю х юятхйяхъ мнбнпнфдеммнцн
- релю ╧ 18 юяхмйкхрхвеяйхе бярюбкемхъ цнкнбйх меопюбхкэмше ярнъмхъ цнкнбйх
- релю ╧ 19 аепелеммнярэ х пндш опх яепдевмн-янясдхяршу гюанкебюмхъу, юмелхъу, гюанкебюмхъу онвей, яюуюпмнл дхюаере, бхпсямнл цхоюрхре, рсаепйскеге
- някнфмемхъ онякепнднбнцн оепхндю х ху опнтхкюйрхйю, 2016
- някнфмемхъ онякепнднбнцн оепхндю х ху опнтхкюйрхйю.
- рЕЛЮ: нЯНАЕММНЯРХ РЕВЕМХЪ Х ОПХМЖХОШ РЕПЮОХХ ОНЯКЕПНДНБШУ ЦМНИМН-БНЯОЮКХРЕКЭМШУ ГЮАНКЕБЮМХИ
- рЕЛЮ: йеяюпебн яевемхе б янбпелеммнл юйсьепябре. дпсцхе бхдш пнднпюгпеьючыху ноепюжхи.
- рЕЛЮ: ПНДНБНИ РПЮБЛЮРХГЛ ЛЮРЕПХ Х ОКНДЮ