АРГАСОВЫЕ КЛЕЩИ (Argasidae)
Бесщитковые Ixodoidea со слабо выраженным половым диморфизмом. Самцы отличаются от самок формой полового отверстия: у самца оно полулунное, у самки — в виде поперечной щели. Тело аргасовых клещей состоит из гнатосомы (комплекса ротовых частей) и собственно тела — идиосомы.
К последнему прнчленены ноги (4 пары у нимф и половозрелых клещей, 3 пары у личинок). Тело клеща одето мягким кожистым покровом. Хитин на дорсальной поверхности имеет у разных видов клещей различную структуру. Хоботок расположен вентрально у половозрелых клещей и нимф близ переднего конца тела или на некотором расстоянии от него и поэтому сверху не виден. У личинок лежит терминально или субтерминально. Все членики пальп свободные: поровые поля отсутствуют у обоих полов. Половое отверстие находится между основаниями первой и второй пары ног. Глаза, если имеются, расположены над коксальной складкой перитремы у нимф и половозрелых обычно впереди IV коксы. Пульвиллы обычно рудиментарны или отсутствуют у половозрелых и иимф, иногда хорошо развиты у личинок.В мировой фауне аргасовых клещей различают четыре рода: Argas1 Ornithodorus, Otobius, Antricola. В Советском Союзе распространены два рода: Argas и Ornithodorus или по классификации М. В. Штром (1946) они являются подсемействами Argasinae и Ornithodorinae.
Аргасовые клещи распространены во всех частях света: в Америке, Азии, Африке, Австралии и Европе.'В СССР их ареал находится в переходной полосе между пустынно-степной центрально-азиатской и пустынной средиземноморской областях (М. В. Поспелова-Штром, 1953). Большинство видов обитает в засушливых, жарких местностях, приурочены к пустынной, полупустынной, степной зонам, равнинам, предгорьям, встречаются в горах, по каменистым осыпям, оврагам, родникам, долинам рек. Широко распространены в Средней Азии и Казахстане Ornithodorus papillipes, Ornithodorus Iahorensis, Argas persicus.
Кроме того, О. Iahorensis и A. persicus встречаются в Азербайджане, Армении, Грузии, Дагестане; A. persicus на юге Украины, Северном Кавказе; О. tartakovskyi распространен в Казахстане и в Средней Азии; О. Cholodkowskyi — в Туркмении и Узбекистане; О. πereeπsis — в Туркмении; О. verrucosus — на Кавказе и Украине; О. alactagalis — в Закавказье (в Армении, Азербайджане, в Восточной Грузии) и в Туркмении; О. Coniceps — в Таджикистане, Киргизии, Казахстане, в Крыму, единично в Армении, Азербайджане; A. reflexus — в Казахстане, Средней Азии, на юге Украины.Медицинское значение. Изучение роли аргасовых клещей как хранителей и переносчиков возбудителей представляет большой интерес. Эта группа клещей — немногочисленная по видовому составу — включает в себя представителей с широким и массовым распространением. Среди них есть виды, являющиеся источником инфекции в природных очагах. C аргасовыми клещами в первую очередь тесно связано распространение клещевых возвратных тифов. Переносчик клещевого спирохетоза впервые в Советском Союзе был установлен в 1927 г., когда Н. И. Латышев заразился возвратным тифом в результате кормления на себе клещей О. papillipes. Дальнейшее изучение переносчиков этого заболевания осуществлялось Е. Н. Павловским и многочисленными последователями под его руководством.
К настоящему времени переносчиками спирохет (боррелий) в нашей стране установлены: О. papillipes (Средняя Азия, Казахстан), О. tartakov- skyi (Туркмения, Казахстан), О. nereeπsis (Туркмения), О. verrucosus (Азербайджанская ССР, Грузинская ССР, Ставропольский край. Украинская ССР), О. alactagalis (Армянская ССР, Азербайджанская ССР), О. cho- Iodkowskyi (Туркменская ССР, Узбекская ССР).
Клещи в любой фазе и стадии развития заражаются спирохетами п:» питании на больных животных. При этом только часть клещей получат- спирохеты, остальные их не содержат (Е. Н. Павловский, А. Н. Скрынн-:- 1939).
Имеется тесная связь между отдельными видами клещей и определенными спирохетами.
Так, клещи О. Iartakovskyi являются переносчиками спирохет (В. Iatyschevi), патогенных для белых мышей, песчанок и человека и непатогенных для морских свинок. Зарубежные виды — нимфы Ornithc- dorus turicata — показали полную передачу спирохет (В. turicata), а клещ О. talaje были отрицательными в этом отношении. Нимфы О. talaje, наоборот показали передачу спирохет В. talaje в 70% случаев и не отмечено передач - этой спирохеты нимфами О. turicata при заражающих кормлениях на мышах (цит. по Артуру, 1962).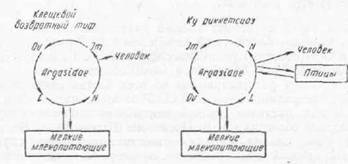
Рис. 71. Тип циркуляции возбудителей болезнен, передаваемых аргасовыми клещами.
Спирохеты широко распространены в различных тканях и органах клещей. Их находили в мальпигиевых сосудах, яичниках, коксальной жидкости, в слюнных железах, в мышечной и соединительной тканях, в нервной системе; онн отсутствуют в экскрементах клещей.
Клещи сохраняют спирохет в процессе своего метаморфоза и передают их через яйцо следующему поколению (рнс. 71). Трансовариальная передача доказана для О. papillipes, О. verrucosus, О. Iartakovskyi (И. В. Троицкий, 1945; Е. Н. Павловский, А. Н. Скрынник, 1948; С. П. Канделаки, 1941).
Инфицирующая способность у О. papillipes сохраняется пожизненно; неснижается при многолетнем голодании клещей (до 14летсрок наблюдений) и содержании клещей при низкой температуре. По данным А. Н. Скрынник (1948), охлаждение до 17° или нагревание до 37° не снизило инфицирующую способность О. papillipes.
Длительное выживание голодных клещей Ornithodorus с сохранением в их организме спирохет, способность трансфазовой и трансовариальной передачи спирохет обеспечивают длительное и устойчивое сохранение природных очагов клещевого спирохетоза без вмешательства человека. В Туркмении П. А. Петрищева (1961) наблюдала сохранение в активном состоянии природных очагов в течение 16—30 лет.
Известно участие аргасовых клещей в эпидемиологии и эпизоотологии других природноочаговых инфекций.
Спонтанная зараженность бруцеллами О. Iahorensis установлена в Казахской CCP (М. М. Ремениова, В. Н. Кусов, 1951) в Армении (В. Н. Зильфян, Е. Л. Аранян, 1953).Выявлена спонтанная зараженность возбудителем чумы О.tartakovskyi (О. В. Афанасьева, 1960). Патогенные лептоспиры, идентичные серотипу Казахстанский II, установлены у О. tartakovskyi, собранных на окраине Муюнкумов (Я. А. Благородный, 1961).
Аргасовые клещи важное значение имеют как хранители и переносчики возбудителя Ку-лихорадки в нашей стране и за рубежом. Доказано спонтанное носительство риккетсий Бернета клещами Argas reflexus, снятых с воробьев в Узбекской CCP (3. М. Жмаева, А. А. Пчелкина, И. К. Мищенко, Б. Е. Карулин, 1955; Лысункипа, 1960), Argas persicus в Узбекской CCP (М. П. Чумаков, 1954; Т. П. Повалишина, 1953), Ornithodorus tartakovskyi из нор большой песчанки в ТуркменскойCCP (3. М. Жмаева, А. А. Пчелкина, Б. Е. Карулин, H. К- Мищенко, Р. И. Зубкова, 1954), Ornithodorus papil- Iipes из нор волка, лисицы в Туркменской CCP (Юго-Западный Копетдаг) (П. А. Петрищева, 3. М. Жмаева, А. А. Пчелкина, 1963). В зарубежных странах выделены риккетсии Бернета из клещей Otobius megnini в Калифорнии, в Лос-Анжелосе (Джелисон, 1948), Ornithodorus moubata в Конго (Жиру, Жадан, 1950), Ornithodorus erraticus в Марокко (Блан, Брюно, 1955), Ornithodorus Iahorensis в Турции (Пейзан и Акки, 1953).
Легко воспринимают возбудителей различных инфекций аргасовые клещи в эксперименте. Доказана восприимчивость к возбудителю бруцеллеза О. papillipes (И. Ф. Самсонов, 1940), A. persicus (М. М. Ременцова, 1949, 1953), О. Iahorensis (М. М. Ременцова, В. Н. Курсов, 1950, 1953); к возбудителю чумы A. persicus (Т. Д. Фадеева, 1932), к возбудителю туляремии О. Iahorensis (Л. С. Ершова, 1959), О. papillipes (Н. Г. Олсуфьев, 1950, Л. С. Ершова, 1963), к возбудителю лихорадки Ky О. moubata (Дэвис, 1943, Вейер, 1953, Бюргдорфер, 1951), О. Iiermsi (Дэвис, 1953), О. turicata, О. parkeri (Дэвис, 1940, 1943), О. gur∏eyi (Смис, 1942), О.
Iahorensis (Пейзан, 1948), О. talaje [Айренс (Irens, 1945)1, Otobuis megnini ІСтоннер (Stonner, 1951)1, О. papillipes (Сягаев, Кондратьев, 1959; И. А. Шифрин, А. Н. Скрынник, 1959; А. Б. Дайтер, II. И. Амосенкова и др., 1961; 3. М. Жмаева, П. А. Петрищева, А. А. Пчелкина, 1963).Следует отметить, что случаи успешной передачи животным возбудителей аргасовыми клещами (за исключением спирохет клещевого рекурренса) очень редки, несмотря на их высокую зараженность некоторыми инфекциями. Так, О. erraticus (Блан и др., 1947; Комбиеску и др., 1953), О. moubata и О. hermsi (Дэвис, 1947) могут во время сосания передавать риккетсии Бернета грызунам. Первый вид передавал риккетсии укусом через 428, второй — через 477 дней. Отмечена передача риккетсий Бернета рогатому скоту во время кровососания О. megnini (Джелисон и др., 1948).
В отдельных случаях О. Iahorensis способен передавать бруцеллы через укус лабораторным и диким животным (Е. Н. Павловский и И. Г. Галузо, 1948).
Наблюдали длительное сохранение возбудителя чумы у О. Iartakovskyi — 171 день (Т. Д. Фадеева, 1932), бруцелл у О. papillipes в течение года (П. Ф. Самсонов, 1940). Риккетсии Бернета сохраняются в клещах О. turicata IOOl день (Гордон, Дэвис, 1940), О. moubata— 670 дней, О. hermsi — 979 (Дэвис, 1943), О. parkeri — 872 (Дэвис, 1943), О. gurney і 538 (Смис, 1942), О. Iahorensis — 19 месяцев (Голем, 1951), О. papillipes — 816 дней при 26— 27е и 797 — при 16° (А. Б. Дайтер, Н. И. Амосенкова, 1961).
Известно сохранение возбудителя туляремии в течение 943 дней у О. Iahorensis (Л. С. Ершова, 1961), 450 дней у О. moubata, О. turicata, О. hermsi (Бюргдорфер и Оуэн, 1956).
Аргасовые клещи способны сохранять возбудителей некоторых болезней в процессе метаморфоза и иногда трансовариально. Например, риккет- 25 Заказ № 228
сии Бернета передаются клещами О. papillipes трансфазово и трансом- риально (И. А. Шифрин, А. Н. Скрынник, 1959; А. Б. Дайтер, Н. И. Am^ сенкова и др., 1961; 3.
М. Жмаева, П. А. Петрищева, А. А. Пчелкина, 196;- . О. moubata (Дэвис, 1943; Вейер, 1953). Передача риккетсий Бернета тать.-.- по ходу метаморфоза изучена также еще у клещей О. erraticus, О. herπ^.ii (Блан и др., 1940), О. turicata (Дэвис, 1943). Клещи О. papillipes передаг - возбудителя туляремии трансфазово (Л. С. Ершова, 1963).Известно выделение вирулентных риккетсий Бернета из организма клещей с экскрементами и коксальной жидкостью (А. Б. Дайтер, Н. И. Amg- сенкова, 1961). Мертвые клещи A. re∏exus сохраняют жизнеспособных риккетсий Бернета 6 месяцев, О. papillipes—11 лет, что не является пределом (3. М. Жмаева, А. А. Пчелкина, Н. К. Мищенко, Б. Е. Карулин. 1955; П. А. Петрищева, 3. М. Жмаева, А. А. Пчелкина, 1963).
Аргасовые клещи благодаря своей способности воспринимать и длительно сохранять возбудителей различных инфекций могут быть широко использованы в качестве благоприятной среды в экспериментальных исследованиях заболеваний человека.
Аргасовые клещи относятся к группе подстерегающих в убежищах и норовых кровососов (В. Н. Беклемишев, 1942, 1945). В отличие от иксодид большинство видов аргасовых клещей питаются на животных короткое время в каждой активной фазе развития — от 2—3 минут до 1—2 часов и более. При этом они поглощают умеренное количество крови. Для них характерно многократное кровососание в одной и той же фазе жизненного цикла. Исключение составляют О. Iahorensis, у которых питание продолжается 20—40 дней. Личинки A. persicus на цыплятах пьют кровь 41∕s— 5 суток. У последних питание имеет много общего с йксодовыми клещами (Ю. С. Балашов, 1957).
У аргасовых клещей более сложный цикл развития от яйца до половозрелого клеща, чем это наблюдается у иксодид. Они проходят пять и более активных фаз развития; личинку, несколько нимф (от 3 до 5) и половозрелую фазу (самцы, самки).
Самки аргазид откладывают десятки или сотни яиц за один прием и до тысячи за всю жизнь — в теплый сезон года (в июле, августе — октябре). О. papillipes откладывает за один прием от 15—60 до 200 яиц (Е. Н. Павловский и А. Н. Скрынник, 1937).
Имеет место автогенное развитие яиц (Е. Н. Павловский, 1946).
Развитие клещей происходит в большинстве случаев по треххозяинному и многохозяинному типам и иногда по однохозяинному. Последнее характерно для О. Iahorensis; от личинки до нимфы (третьей) его развитие осуществляется на одном и том же животном.
Полный цикл развития у различных видов длится от 1 года до нескольких лет. Он может продолжаться более 2 лет у О. tartakovskyi, 2 года 8 месяцев у О. aloctaga- Iis (М. В. Поспелова-Штром, 1953), 3 года у A. reflexus (И. Г. Галузо, 1957), не менее 2, 4, 8 лет у О.verrucosus (В. В. Слесаренко, 1959, А. Н. Скрынник, 1939. М. В. Поспелова-Штром, 1953), 16 лет у О. Iahorensis1 3, 14, 25 лет у О. papillipes (Е. Н. Павловский, А. Н. Скрынник, 1959; Н. А. Филиппенко, 1935).
Особенностью метаморфоза клещей рода Omithodorus является то, что самцы и самки могут появляться по ходу развития отдельных особей одной семьи 2, 3 раза — после линек нимф Ill, IV и V (Е. H. Павловский, 1952). О. papillipes может иметь от III до VI нимфальных стадий, О. verrucosus — от IIl до V, О. tartakovskyi — от 11 до IV, О. Coniceps — IV, V, О. Iahorensis — от III до VI, A. persicus, A. reflexus — III. Продолжитель-
ность метаморфоза определяется условиями среды и наличием прокормителей.
В отсутствие хозяев метаморфоз у клещей затягивается на многие годы — до 25 лет. Это возможно в связи со способностью клещей к длительному голоданшо. О. papillipes может голодать в фазе нимф 1 до 2 лет, нимф II — до 4 лет, нимф III и половозрелых клещей — от 5 до 10 лет (А. И. Скрынник, 1951), 10—14 лет (Е. И. Павловский, 1960). Личинка этого клеща голодать долго не может: при 26° — в течение 20 дней, реже 30, в комнатных условиях — 15 месяцев (Е. Н. Павловский, 1960).
Личинки О. verrucosus голодают 300—200 дней, остальные стадии более 2—3 лет (А. Н. Скрынник, 1948, М. В. Поспелова-Штром, 1953). Меньшие сроки могут голодать клещи О. tartakovskyi: в течение 2 лет половозрелые особи, 72—328 дней личинки (М. В. Поспелова-Штром, 1953), более 4 лет половозрелые О. Iahorensis, до 1 года личинки, 5—6 лет половозрелые A. reflexus (И. Г. Галузо, 1957).
Аргасовые клещи обладают способностью выпрыскивать коксальную жидкость после отпадения с хозяина (у видов нашей фауны) или во время питания на хозяине (О. moubata), что обеспечивает лучшую подвижность для предельно насосавшихся клещей.
Хозяева. Аргасовые клещи являются полифагами, что стоит в тесной связи с их образом жизни как паразитов, подстерегающих добычу в убежищах или в норах. Клещи редко покидают свое убежище и поэтому вынуждены питаться на животных, которые обитают в этих убежищах или их посещают. Особенно часто наблюдается последнее. Прокормителями клещей служат разнообразные животные: млекопитающие, птицы, рептилии, амфибии. Наибольшее число хозяев имеют О. papillipes, О. verrucosus. Эти клещи способны питаться на любом позвоночном животном. Однако замечено, что кровь разных животных не является в одинаковой степени подходящей для них пищей. Для клещей, кормящихся нормально на млекопитающих, кровь кур не является полноценной. Клещи О. papillipes охотно пьют кровь кур, но не все сытые нимфы линяют в следующую стадию без добавочного питания на млекопитающих, иногда погибают, тогда как половозрелые клещи, питаясь па курах, не имеют никаких отклонений от нормы.
П. А. Петрищева (1962) наблюдала массовую гибель О. tartakovskyi и О. papillipes после кровососания на птенцах сизоворонках и щурках.
Argas persicus — паразиты птиц при питании кровью человека, морской свинки иногда погибают или выживают, но мало откладывают яиц (М. В. Поспелова-Штром, 1953). Высокая смертность клещей этого вида наблюдается при питании на жабах, их кровь является для них токсичной (цит. по Артуру, 1962).
Более узкий круг хозяев характерен для клещей О. Iahorensis -— домашние животные (овцы, верблюды, реже крупный рогатый скот, козы, ослы, собаки) и норовых О. alactagalis — грызуны, хищники (песчанки, тушканчики, хомячки, барсуки, лисы и др.).
Биотопы. На хозяйственной территории аргасовые клещи О. papillipes, О. Iahorensis, A. persicus заселяют старинные, примитивные глинобитные и кирпичные жилища человека, надворные постройки — помещения для скота, глинобитные и кирпичные дувалы. О. papillipes селится в массе, кроме того, в необитаемых человеком постройках — мечетях, клубах и т. п. (рис. 72). О. Iahorensis и A. persicus редко встречаются в жилищах человека и в массе заселяют надворные постройки (помещения домашних животных). В природных биотопах аргасовые клещи в районах своего распространения обитают в норах и убежищах насекомоядных, грызунов,
25»
хищных животных, в гнездах птиц. Они многочисленны в убежищах ежей в норах большой, полуденной, краснохвостой песчанок, реже встречаются в норах гребенщиковой песчанки, тонкопалого суслика и т. д. Пещеры с норами дикобразов бывают в массе заселены О. papillipes, иногда О. tar

Рис. 72. Местообитание Ornithodorus papillipes. Окрестности Cepaxca. Древняя мечеть.

Рис. 73. Местообитание Ornithodorus. Туркмения. Предгорья Копет-
Дага.
takovskyi. Норы черепах, варанов являются обычными местами обитания О. tartakovskyi (рис. 73). В большинстве случаев эти животные занимают норы грызунов, но иногда они делают их сами.
Пещеры естественные, а также созданные человеком в давние времена являются характерными биотопами О. papillipes. В этом типе убежищ обитает наибольшее число видов этого рода фауны мира.
В СССР пещерные виды О. Cholodkovskyi и О. papillipes одновременно обитают и в постройках человека.
Аргасовые клещи не способны к дальним миграциям. Однако при появлении животных или человека, например О. papillipes, выползают из трещин, нор и быстро направляются к своей добыче. Отмечено выползание половозрелых клещей из нор большой песчанки в Кара-Кумах в ранние утренние часы до прогревания песка (П. А. Петрищева, 3. М. Жмаева, 1962).
Хорошо выражены у Ornithodorus сезонные передвижения. В зимнее время клещи прячутся в глубине нор; весной и осенью (март — апрель, сентябрь — октябрь) клещи встречаются в их устьях.
Сезонная активность. Большинство видов аргасовых клещей во всех фазах развития активны в сухой жаркий период года с марта по август, сентябрь, октябрь. Активные клещи могут встречаться в Средней Азии круглый год: в теплых помещениях для скота, в жилищах человека — О. papillipes, в курятниках ■— A. persicus, в больших пещерах с животными— О. papillipes, О. Cholodkovskyi (с летучими мышами).
В голодной степи Таджикской CCP (X. Кузыбаева, 1961) половозрелые и нимфы старших возрастов О. papillipes, О. tartakovskyi встречаются с марта по август, личинки и нимфы первого возраста — в июле и августе, молодые нимфы и половозрелые клещи — осенью.
В предгорных и горных районах Сурхандарьинской области Узбекской CCP в осенне-зимний период на животных паразитируют О. Iahorensis. О. papillipes (Т. Е. Куклина, 1962).
Агрессивными видами по отношению к человеку являются О. papillipes и О. verrucosus. Остальные виды рода Ornithodorus (О. tartakovskyi, О. cholodkovskyi) и рода Argas (A. persicus) редко нападают на человека.
Еще по теме АРГАСОВЫЕ КЛЕЩИ (Argasidae):
- Происхождение и эволюция
- ВВЕДЕНИЕ
- Женская половая система
- ПРОДОЛЖИТЕЛЬНОСТЬ ПИТАНИЯ
- ЦИТОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РАЗМНОЖЕНИЯУ САМОК
- ЗАКЛЮЧЕНИЕ
- Аргасовые клещи (сем. Argasidae).
- СЕМЕЙСТВО АРГАСОВЫЕ КЛЕЩИ (ARGASIDAE)
- АРГАСОВЫЕ КЛЕЩИ (Argasidae)
- ЛИТЕРАТУРА
- БОРЬБА C АРГАСОВЫМИ КЛЕЩАМИ