РОТОВОЙ АППАРАТ
Строение ротового аппарата было изучено на серийных срезах самок Ornithodoros papillipes и Hyalomma asiaticum и участков покровов хозяев вместе с прикрепившимися к ним нимфами Н.
asiaticum в разной степени насыщения, а также на тотальных препаратах отдельных отпрепарованных частей гнатосомы. Анатомическое строение гнатосомы было рассмотрено ранее (см. стр. 8—11), и в настоящем разделе рассматриваются ее функционально-морфологические особенности, необходимые для понимания механизма приема пищи.Hyalomma asiaticum. Нижняя половина хоботка образована вытянутым вперед выростом основания гнатосомы — гипостомом. Последний имеет слегка ложковидную форму, расширяясь и уплощаясь к дистальному концу (рис. 185—188). Его вентральная поверхность выпуклая и покрыта рядами мощных зубцов, отогнутых назад. Дорсальная поверхность гипостома слегка вогнутая и медиально несет пищевую борозду. В дистальной части она представлена сравнительно мелким U-образным углублением. Проксимально борозда углубляется, фланкируется с боков двумя продольными гребнями эпикутикулы, и просвет ее на поперечном разрезе становится Ѵ-образным. В дистальной части пищевой борозды кутикулиновый слой плотно прилегает к экзокутикуле (рис. 188), а в проксимальной отходит от нее и образует так называемую гипостомаль- ную мембрану, так что между ними создается свободное пространство (рис. 189—190).
В работах Р. Артура (Arthur, 1951, 1953, 1957, 1962) эта гипостомаль- ная мембрана ошибочно трактуется как эпифаринкс, сросшийся по краям с гипостомом, а прикрываемая им сверху полость — как пищевой канал. Наши наблюдения заставляют отказаться от подобного толкования, так как пищевой канал проходит не под, а над гипостомальной мембраной и расположенная иод ней полость отсутствует в передней половине хоботка, не открываясь наружу. Подобного же мнения придерживается Д. Грегсон (Gregson, 1960).
У Hyalomma asiaticum и других изученных нами видов поднятия и опускания мембраны оказались очень незначительными, чисто пассивными и вряд ли существенно влияют на изменения объема предротовой полости во время питания.Внутри гипостома по всей его длине прослеживается дорсо-вентраль- ный шов, свидетельствующий о первично парном происхождении гипо-
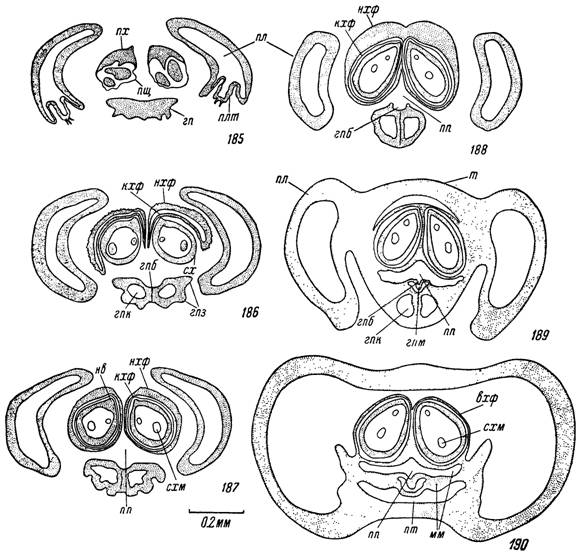
Рис. 185—190. Строение передней половины гнатосомы самки Hyalomma asiati-
сит на поперечных срезах.
Срезы на уровне: 185 — концов пальцев хелицер, 186 — начала наружных хе- лицеральных футляров, 187 — середины наружных хелицеральных футляров, 188 — слияния медиальных стенок хелицеральных футляров, 189 — начала замкнутой предротовой полости, 190 — передней части основания гнатосомы.
т — тектум; пл — пальпы; плт — терминальный пальпальный членик; сх — ствол хелицеры; мо — мембрановидное окно; пх — подвижный палец хелицеры; пщ — мембрановидный плащ; нхф — наружные и кхф — конические хелицеральные футляры; вхф — внутренние хелице- ральные футляры; хм — внутрихелицеральные мышцы; схм — их сухожилия; гп — гипостом; гпз — гипостомальные зубцы; гпк — внутригипостомальные каналы; гпм — мембрана пищевой борозды гипостома; гпб — пищевая борозда; пп — предротовая полость; р — рот; ср — слюнной резервуар; сп — главные слюнные протоки; схп — субхелицеральная пластинка; мм — межкоксальный мост; кг — клиновидный гребень; ко — крыловидные отростки; дклі — дорсальные мышцы крыловидных отростков; вкм — то же вентральные; г — глотка; го — вентральные опорные глоточные склериты; дрг — дорсальные, лрг — латеральные и врг — вентральные расширители глотки; ксг — кольцевые сжиматели глотки; пт — переднее продолжение полости тела; не — направляющие выросты стволов хелицер.
стома в результате слияния передних выростов кокс педипальп. В каждой из половин проходит канал. Оба канала сливаются в проксимальной части гипостома в один непарный, представляющий продолжение полости тела (рис.
190). У недавно перелинявших особей в этих внутренних полостях еще сохраняется гиподермальная выстилка.У клещей Ixodes ricinus, I. hexagonus и I. persulcatus (Arthur, 1946, 1951; Никонов-, 1958) в гипостомальной полости описаны группы дорсо- вентральных мышц, идущих от боковых стенок пищевой борозды к вентролатеральный углам полости. При детальном исследовании полных серий срезов мы обнаружили подобные мышцы лишь в самом проксимальном участке гипостома I. persulcatus, но они отсутствовали у Hyalomma asia- ticum, Dermacenlor pictus и Rhipicephalus turanicus. По-видимому, ги- постомальные мышцы свойственны только видам рода Ixodes, и в этом отношении они сходны с аргасовыми клещами.
Дорсальная половина хоботка образуется хелицерами. Стволы хелицер окружены двойными трубчатыми кутикулярными футлярами, происхождение и строение которых до сих пор вызывает споры. Наиболее мощные наружные футляры несомненно представляют передний вырост крыши гнатосомы (тектума). Он расщепляется на переднем конце и дорсально доходит при втянутых хелицерах до оснований пальцев. С вентральной стороны они несколько короче и тоньше, так что в передней части хоботка на поперечных срезах они вначале выглядят как крыша стволов (рис. 186). Загибаясь вентрально, они заключают последние в твердые трубки. Гребневидные медио-дорсальные выросты стенок стволов входят в соответствующие борозды футляров и предотвращают нежелательные боковые перемещения при скольжении хелицер в передне-заднем направлении (рис. 187). В дистальной части хоботка наружные хелицеральные футляры самостоятельны, но плотно прилегают друг к другу почти прямыми внутренними стенками. Не доходя до основания головки, они сливаются сперва по дорсальной и позднее по вентральной границе, так что формируется твердое полукольцо, разделенное вертикальной перегородкой (рис. 189, 189). Нижняя стенка наружных хелицеральных футляров продолжается в погруженную в основание гнатосомы субхелицеральную пластинку.
В дистальной части наружная дорсальная поверхность хелицеральных футляров покрыта многочисленными мелкими зубчиками, облегчающими фиксацию на хозяине во время питания.
Дистальные концы наружных хелицеральных футляров заворачиваются внутрь и в виде мембрановидной оболочки идут назад, прикрепляясь к стволам хелицер спереди от глоточного отверстия (рис. 187—188). Аналогичные образования описаны у Dermacentor andersoni (Gregson, 1960) и названы коническими футлярами хелицер. По-видимому, они играют существенную роль в выдвижении хелицер, предотвращая проникновение крови и посторонних частиц в хе- лицеральную полость во время питания.Позади от конических футляров стволы хелицер вновь окружены мембрановидной оболочкой, отходящей дорсально от внутренней поверхности тектума, а вентро-латерально — от субхелицеральной пластинки (рис. 189—196). Морфологическая трактовка этих образований затруднительна, и обычно их называют внутренними хелицеральными футлярами.
Наибольшей сложностью строения отличается передняя часть основания гнатосомы. Слияние наружных хелицеральных футляров и гипостома в плотное хитиновое кольцо происходит на значительном расстоянии спереди от ротового отверстия, так что образуется обширная морфологически целостная трубка предротовой полости (рис. 189, 190) шириной 0.32 и высотой 0.04—0.08 мм. Ее крышей служит вентральная стенка наружных хелицеральных футляров, а дном — дорсальная поверхность гипостома; стенка и поверхность соединяются между собой латеральновентральными выростами кокс пальп. В передней трети хоботка морфологическое слияние между его компонентами отсутствует, но благодаря их тесному прилеганию друг к другу предротовая полость фактически продолжается до дистального конца гипостома (рис. 25).
По средней линии дорсальной стенки гипостома (дну предротовой полости) проходит пищевая борозда. В дистальной части она сравнительно мелкая (ширина 0.07 и глубина 0.02 мм), а в проксимальном направлении
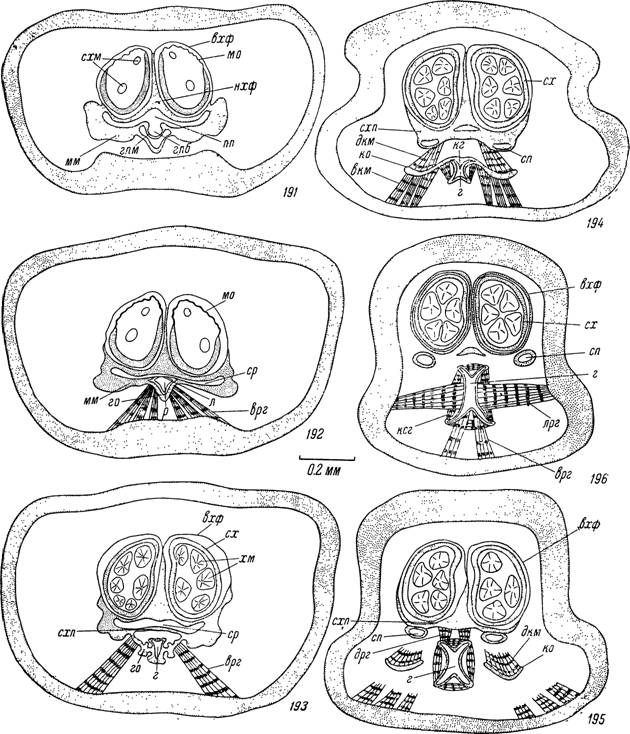
Рис. 191—196. Строение основания гнатосомы самки Hyalomma asiaticum. Поперечные срезы на уровне: 191 — конца предротовой полости, 192 — ротового отверстия, 193 — начала глотки, 194 — переднего глоточного клапана, 195 — середины субхелицеральной пластинки, 196 — заднего конца субхелицер а л ьной пластинки.
Условные обозначения те же, что и на рис.
185—190.постепенно углубляется до 0.04—0.06 мм и приобретает Ѵ-образное поперечное сечение (рис. 190). Ротовое отверстие находится в глубине предротовой полости в месте, где пищевая борозда прикрывается сверху коротким конусовидным лабрумом (рис. 192).
Часть предротовой полости, продолжающаяся назад над лабрумом и дорсальной поверхностью субхелицеральной пластинки, называется слюнным резервуаром (рис. 192—193). Он сильно уплощен дорсо-вент- рально (высота 0.03 и ширина 0.35 мм), и в его заднебоковые углы открываются главные выводные протоки парных слюнных желез размером 0.07 ХО.ОЗ мм. Последние сперва идут в толще субхелицеральной пластинки, а затем отделяются от нее в вентральном направлении (рис. 194— 196). Потолок слюнного резервуара образован вентральной стенкой наружных хелицеральных футляров, переходящей в месте впадения слюнных протоков в субхелицеральную пластинку.
Субхелицеральная пластинка служит главной опорной структурой для хелицер и глотки. В передней части она соединяется вентро-лате- ральными выростами со стенками основания гнатосомы и образует меж- коксальный мост (рис. 190). Далее назад она теряет связь со стенками гнатосомы и сильно утолщается (рис. 193—194). После слияния с вентральной стенкой наружных хелицеральных футляров по ее дорсальной поверхности проходит высокий вертикальный гребень. Он представляет продолжение слившихся медиальных стенок наружных хелицеральных футляров и дорсально переходит во внутренние хелицеральные футляры (рис. 194). С вентральной стороны к утолщенным краям субхелицеральной пластинки прикрепляются пучки мышц, идущие от крыловидных отростков, и дорсальные расширители глотки (рис. 194, 195). Передний конец субхелицеральной пластинки продолжается в короткий лабральный вырост (рис. 25).
Ornithodoros papillipes. Строение ротового аппарата рассматриваемого вида по общему плану строения очень сходно с таковым иксодовых клещей, но имеется и ряд отличий, связанных с различными способами поглощения крови в обеих группах.
Хоботок аргасовых клещей более тонкий и не приспособлен к длительному закреплению клеща на теле хозяина, тогда как пальцы хелицер, напротив, более мощные и приспособлены к быстрому прорезанию покровов (рис. 197, 206).Степень развития зубцов гипостома, величина подвижных пальцев хелицер и терминальное или вентральное положение гнатосомы во многом связаны с продолжительностью питания. Гипостом, вооруженный правильными рядами направленных назад зубов, служит основным органом закрепления в покровах хозяина. Чем дольше продолжается кровососание, тем прочнее должен удерживаться клещ на теле хозяина и тем сильнее должно быть развито вооружение гипостома. Для быстро питающихся видов закрепление не столь важно и на первое место выдвигается задача максимально быстрого поглощения полной порции крови. Поэтому для них существенную роль играет степень развития пальцев хелицер, обеспечивающих внедрение в покровы хозяина.
Внутри хоботка, так же как и у иксодовых клещей, имеется трубкообразная предротовая полость. На поперечных срезах видно, что она сжата в дорсо-вентральном направлении в уплощенный канал. В дистальной части этот канал ограничен тесно прилегающими друг к другу отогнутыми краями хелицер и слегка вогнутым уплощенным гипостомом (рис. 199). На уровне начала хелицеральных футляров трубчатый канал сохраняется за счет соответствующих изгибов вентральной поверхности наружных футляров и увеличивающейся вогнутости гипостомального желоба вместе с его краевыми выростами (рис. 200). Далее назад он, в отличие от иксодид, подразделяется вытянутой верхней губой — «стилетом» на дорсальную и вентральную половины, представляющие самостоятельные каналы (рис. 201).
Нижняя половина предротовой полости снизу и с боков ограничена дном медиального гипостомального желоба, постепенно переходящего
в более глубокий канал. Сверху она прикрыта верхней губой. Дорсальная поверхность последней вместе с латеральными зонами дорсальной стенки гипостома формируют дно верхней половины предротовой полости. Потолок последней образуют слившиеся вентрально наружные футляры хелицер (рис. 202). Нижний канал заканчивается почти треугольным ротовым отверстием (рис. 203), ведущим в глотку. Верхняя трубка открывается в сильно уплощенный слепой мешок слюнного резервуара. Его нижней стенкой служит эпистомальная пластинка, а верхней — вентральная поверхность наружного футляра хелицер (рис. 204). В заднебоковые углы описанной полости открываются протоки слюнных желез. В литературе ее необоснованно называют буккальной, т. е. ротовой, хотя она не имеет никакого отношения к ротовому отверстию. Функционально она представляет резервуар, в котором скапливается слюна, вытекающая из протоков слюнных желез.
Эволюция ротового аппарата. Ротовой аппарат иксодовых клещей представляет среди паразитических клещей крайнюю степень специализации к питанию кровью. Тем не менее в нем можно выявить основные структурные компоненты, свойственные остальным паукообразным. Детальный морфологический анализ ротового аппарата арахнид, выполненный Р. Снодграссом (Snodgrass, 1948), позволил ему воссоздать исходный прототип, общий для всех отрядов (рис. 207).
У взрослых особей часть просомы, происходящая из акрона, включает лабрум, эпистомальный район и узкую медиальную полоску передней стенки тела, соединяющую эпистом с панцирем между основаниями хелицер. Сомиты хелицер остаются посторально, но сами хелицеры в результате преорального перемещения занимают переднее положение, а их стернальные участки почти полностью редуцируются. Педипальпы, исключая Palpigradi, также сдвигаются вперед по бокам рта и дорсально соединяются с эпистомом, формируя межкоксальный мост. Стернальная пластинка педипальпального сегмента образует дейтостернум, выступающий перед ротовым отверстием и формирующий нижнюю губу. Рот погружен в предротовую полость, ограниченную с боков коксами педипальп, снизу разрастаниями кокс педипальп или дейтостернумом и сверху лабрумом, и ведет в глотку сосательного типа.
Ротовой аппарат клещей претерпел по сравнению с приведенной схемой значительные видоизменения, в основных чертах сводящиеся к следующему (рис. 208). Хелицеры и педипальпы объединяются в самостоятельном отделе тела — гнатосоме (капитулуме, головке). В его же состав входит большая часть сохранившегося акрона, тогда как глазная область последнего, как у остальных паукообразных, значительно смещается назад и сливается с тергитами просомальных сегментов (идиосомы иксодовых клещей). Дорсальная поверхность гнатосомы склеротизуется над основаниями хелицер и образует крышу, или тектум. Латерально последний сливается с дорсальными разрастаниями гнатококс педипальп, которые одновременно сливаются между собой вентрально по средней линии. В результате образуется характерное для большинства Parasitiformes, а по мнению Снодграсса вообще для всех клещей твердое хитиновое кольцо основания гнатосомы.
Вентрально передние коксальные отростки педипальп вытягиваются вперед, сливаются по средней линии и образуют гипостом, представляющий дно предротовой полости. Сверху ротовое отверстие прикрыто лабрумом, прикрепляющимся задним концом к субхелицеральной пластинке. Последняя гомологична эпистому других арахнид, так как она всегда участвует в образовании межкоксального моста, а к ее нижней поверхности прикрепляются дорсальные дилататоры глотки. Эмбриологические

Рис. 197—206. Строение гыатосомы самки Ornithodoros papillipes.
197 — вид снизу гнатосомы и окружающих ее частей идиосомы, пальпа слева удалена;
198 — продольный разрез; поперечные срезы на уровне: 199 — дистального конца гипостома, 200 — переднего конца лабрума, 201 — средней части гипостома, 202 — слияния наружных хелицеральных футляров, 203 — ротового отверстия, 204 — слюнного резервуара и переднего конца глотки, 205 — эпи стома и оснований
хелицеральных стволов; 206 — отдельная отпрепарованная хелицера.
кл — клюв идиосомы; щк — щеки идиосомы; п — пальпа; г — гипостом; ог — основание гнатосомы;: рж — рог органа Жене; пх — подвижный палец хелицеры; сх — ствол хелицеры, к — камеросто- мальная складка; ох — основания стволов хелицер; нф — наружные, кф — конические и ѳф — внутренние хелицеральные футляры; т — тектум; вх — внутрихелицеральные мышцы; кп — каналы сухожилий внутрихелицеральных мышц; скн — слюнной канал предротовой полости; пкн — пищевой канал предротовой полости; лб — лабрум; пк — передняя лабральная камера; зк — задняя лабральная камера; ср — слюнной резервуар; эп — эпистом; эпг — боковые гребни эпистома; гпб — гипостомальная борозда; гп — внутригипостомальные каналы; ргж — мышцы-расширители гипостомального желоба; р — рот; плв — клиновидный вырост; гл — глотка; дргл — дорсальные, лргл — латеральные и вргл — вентральные мышцы-расширители глотки; пс — главные выводные-
протоки слюнных желез; х — хелицера.
наблюдения над формированием гнатосомы в основном подкрепляют главные положения Снодграсса.
Ближе всего к гипотетическому прототипу ротового аппарата клещей приближается гнатосома некоторых свободноживущих гамазид, как например Poecilochirus carabi, детально изученной В. Винклером (Winkler, 1886). Усложнения в ее строении связаны с появлением мембрановидных футляров хелицер, с различными модификациями гипофаринкса и лабраль- ных отростков, а также с развитием парных передних гипостомальных выростов.
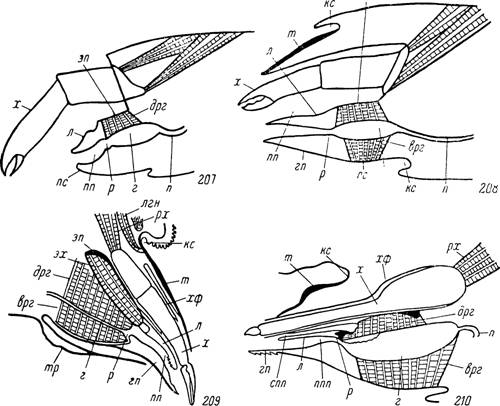
Рис. 207—210.
207 — схема строения передней части просомы паукообразного; 208 — то же гнатосомы кдеща (по Snodgrass, 1948); 209 — строение гнатосомы клеща Sauronyssus saurarum (по Лагутенко, 19626); 210 — то же Отпі- thodoros papillipes (по Балашову, 19616).
m — тектум;£кс — камеростомальная складка; х — хелицера; хф — хелицеральный футляр; рх —мышцы-ретракторы хелицер; эх — мышцы-экстензоры хелицер; лги — мышцы-леваторы гнатосомы; эп — эпистом; л — лабрум; гп — гипостом; пп — предротовая полость; ппп — пищевой канал предротовой полости; спп — то же слюнной канал; р — рот; г — глотка; врг — вентральные расширители глотки; дрг — дорсальные расширители глотки; п — пищевод; пс — педипальпальный стернит; тр —
тритостернум.
Недавно А. Б. Ланге (1962), опираясь на взгляды А. А. Захваткина (1952) о полифилетическом происхождении клещей, выдвинул новую трактовку сегментального состава их ротового аппарата. А. Б. Ланге считает, что в состав гнатосомы в разных отрядах клещей входит различное число сомитов, а четкая интерсегментальная граница между этим отделом и просомой отсутствует. В связи с этим он предлагает пользоваться более нейтральным термином — гнатемой.
Не затрагивая весьма дискуссионный вопрос о филогенетических взаимоотношениях разных групп клещей, нам кажется, что в отношении отряда или подотряда Parasitiformes нет никаких оснований отказываться от общепринятого термина гнатосома, независимо от систематического ранга этой группы. Морфологический состав ее у всех Parasitifor- mes однороден, а отсутствие интерсегментальной границы с просомой не имеет принципиального значения. Задняя граница просомы с описто- сомой у многих паукообразных также не интерсегментальна, но это не вызывает сомнения в самостоятельности данных тагм.
Переход к питанию кровью внес существенные изменения в строение гнатосомы клещей, причём исключительно специализированный ротовой аппарат иксодид можно понять лишь при сопоставлении с таковым более примитивных кровососущих гамазид, детально исследованных в этом отношении (Hughes, 1949; Gorirossi, 1950; Лагутенко, 1962а, 19626). Для ротового аппарата Haemagamasus nidi характерно сохранение многих примитивных черт, в частности массивных клещевидных хелицер с грубыми зубцами по краям обоих пальцев, а также широкий и короткий ротовой конус с незамкнутой предротовой полостью. Этот вид еще не способен получать кровь путем активного прокола кожи хозяина, и ему свойствен смешанный тип питания, включающий подъедание высохшей или полужидкой крови вокруг ранок, схизофагия и энтомофагия (Нель- зина и Данилова, 1956).
Dermanyssus gallinae, Sauronyssus saurarum и Ornithonyssus bacoti — представители группы облигатных кровососов. Их хелицеры преобразуются из грызущих в колюще-режущие органы. Стволы их становятся тоньше и значительно удлиняются. Одновременно происходит вытягивание ротового конуса и образуется почти замкнутая с боков трубковидная предротовая полость (рис. 209). Эти виды уже самостоятельно прорезают покровы позвоночных и пьют вытекающую из ранок кровь.
Клещи надсем. Ixodoidea относятся к числу наиболее совершенных кровососов и их ротовой аппарат приспособлен к поглощению крови непосредственно в толще кожи хозяина, куда он погружается на время питания (рис. 210). Длинные стержневидные хелицеры полностью утратили первичную клешневидную форму, а число их члеников уменьшается до двух. Подвижность их пальцев с режущими зубцами ограничена медио- латеральной плоскостью, и они превратились в исключительно эффективный режущий орган, легко проникающий сквозь твердый роговой слой эпидермиса.
Наружные хелицеральные футляры и гипостом с боков тесно прилегают друг к другу и формируют сильно вытянутый в переднем направлении хоботок с внутренней трубковидной предротовой полостью. Последняя служит проводящим каналом, обеспечивающим откачку крови или инфильтрата непосредственно из внутренних слоев дермы, очень богатых кровеносными сосудами.
Наряду с этими общими чертами между иксодовыми и аргасовыми клещами наблюдаются значительные различия в строении гнатосомы. Они преимущественно адаптивного характера, связаны с различными типами паразитизма в этих семействах; детально они рассмотрены в предшествующем разделе. По ряду морфологических признаков ротовой аппарат аргазид менее специализирован по сравнению с иксодовыми клещами. Хоботок у них вытянут меньше и внутри его сохраняется длинный эпистом, разделяющий предротовую полость на значительном протяжении на вентральный пищевой и дорсальный слюнной каналы. Все четыре пальпальных членика хорошо развиты и подвижно соединены между собой.
Еще по теме РОТОВОЙ АППАРАТ:
- Изолирующий дыхательный аппарат ИП-6
- СОБАЧИЙ КЛЕЩ
- МОШКИ СИМУЛИДЫ - семейство Simuliidae
- ХИАЛОМА
- 38. Работа световоспринимающего аппарата глаза
- Постельный клоп.
- СЛЕПНИ |OTP. DIPTERA, СЕМ. TABANlDAEJ
- Двуустка печеночная (Fasciola hepatica)
- 2.1. Исследуемые препараты
- КЛАСС СОБСТВЕННО КРУГЛЫХ ЧЕРВЕЙ — НЕМАТОДЫ,NEMAТООА